miRNA 在缺血性腦卒中神經保護領域的研究進展
2023-12-13 04:53:26李泓宇蘭瑞付雪琴王瑋瑋鄒旭歡王漫漫張勇唐琛劉
中國比較醫學雜志 2023年11期
關鍵詞:氧化應激
李泓宇蘭 瑞付雪琴王瑋瑋鄒旭歡王漫漫張 勇唐 琛劉 雙
(1.河南中醫藥大學,鄭州 450000;2.河南中醫藥大學第一附屬醫院,鄭州 450000;3.鄭州大學第三附屬醫院,鄭州 450000)
腦卒中是一種殘疾率、復發率和死亡率都很高的腦血管疾病,嚴重影響了患者的生活質量[1]。 腦卒中可以分為缺血性腦卒中(ischemic stroke,IS)和出血性腦卒中,其中87%的病例屬于缺血性腦卒中[2]。 盡管組織纖溶酶原激活劑( tissue plasminogen activator,tPA)溶栓和血管內血栓切除術是臨床治療的重要手段,但改善卒中后長期恢復的方法仍然有限。 因此,保護神經細胞免受腦缺血性損傷一直是IS 的治療目標。
miRNA 屬于小的非編碼RNA 分子,可以通過靶向mRNA 的3’UTR 或促進mRNA 的降解來調節基因表達[3]。 miRNA 主要調節腦內關鍵基因的表達,在細胞凋亡、抗炎、抗氧化應激、神經分化、腦血管生成等過程中均發揮了重要作用[4]。 本文綜述了近年來miRNA 參與缺血性腦卒中的神經保護的文獻證據,從而指出其在神經元保護方面的潛力。
1 miRNA 概論
miRNA 是由18 ~25 個核苷酸組成的小的單鏈非編碼RNA 分子,miRNA 來源于細胞核,它首先被RNA 聚合酶Ⅱ轉錄為初級miRNA,然后通過微處理器切割成前體miRNA,隨后又被轉移到細胞質中并加工成雙鏈RNA,成為成熟的miRNA。 成熟的miRNA 被釋放到受體細胞中參與轉錄并進行基因表達,同時調節多種細胞和分子途徑[5]。 miRNA 幾乎在所有細胞活動中都起著至關重要的作用,它可通過多種途徑發揮生物學功能[6-7]。 并且不同的miRNA 可在缺血性損傷后負責腦細胞之間的通信,缺血后腦細胞產生的miRNA 可以穿過細胞外囊泡或外泌體中的血腦屏障(blood brain barrier,BBB),然后在腦缺血再灌注后的神經修復機制中起著調節作用[8]。
2 miRNA 在缺血性腦卒中神經保護作用
據報道,miRNA 在中樞神經系統疾病發生發展中起著重要的作用[9-10]。 Gámez-Valero 等[11]研究表明,miRNA 對不同退行性癡呆疾病具有神經保護作用,還反映了癡呆疾病差異性特征,有助于路易體癡呆和阿爾茨海默病的鑒別診斷。 Bai 等[12]研究證實,在帕金森疾病中,miRNA 抑制了神經膠質細胞中的NF-κB 信號傳導,減少了神經膠質細胞活化和神經炎癥,多巴胺產生增多,進而達到了神經保護作用。 然而miRNA 在IS 中的神經保護機制了解甚少,有研究認為[3,13-15],miRNA 對缺血性腦卒中的神經保護作用與抑制炎癥、減少凋亡、促進血管新生、抑制氧化應激、增強神經可塑性和神經發生等機制有關。
2.1 miRNA 抑制炎癥反應
炎癥反應是IS 神經元死亡的主要原因之一[16],炎癥反應在腦缺血再灌注損傷(ischemia/reperfution,I/R)后幾分鐘內開始,它的發生機制包括腦駐留細胞的激活和外周白細胞通過BBB 浸潤到腦實質中,活化的小膠質細胞和星形膠質細胞與浸潤的外周免疫細胞一起釋放活性氧(reactive oxygen species,ROS)、促炎細胞因子和其他神經毒性物質,從而導致神經元死亡[17]。 Wang 等[18]實驗證實,miR-34-73b 拮抗劑可以顯著減弱大鼠腦組織缺血中促炎因子(iNOS、COX-2、TNF-α 和IL-6)和細胞因子信號傳導抑制因子3(suppressors Of Cytokine Signaling 3,SOCS3)的表達,來保護神經元細胞,并且在體外實驗中取得相同結果。 郭泉等[19]發現,miR-221 在腦缺血再灌注大鼠模型中,下調miR-221的表達后可以明顯抑制炎癥反應,從而達到神經保護的作用。 保護BBB 的完整性減弱其滲透性也是抑制炎癥反應的重要途徑之一。 Bernstein 等[20]研究表明,miR-98 減少了促炎因子Ly6C 的浸潤,并降低了受影響區域內活化的M1 小膠質細胞的數量,進而減弱BBB 滲透性從而達到神經保護的作用。而來自外周白細胞的miR-29b 則可以通過腫瘤壞死因子(C1q tumor necrosis factor,C1QTNF)來增強BBB 完整性,從而減弱炎癥反應[21]。 也有其他研究證實,來源于間充質干細胞(mesenchymal stem cells,MSC)的外泌體miRNA 對炎癥反應具有抑制作用。例如Zhao 等[22]發現來自MSC 的外泌體miR-223-3p 抑制了M1 小膠質細胞極化引起的促炎反應,減輕了腦缺血再灌注損傷,從而改善了神經功能障礙,促進了學習和記憶能力。 來源于MSC 的外泌體miR-542-3p 通過抑制 Toll 樣受體( toll-like receptor 4,TLR4)來抑制腦缺血后導致的神經膠質細胞炎癥反應[23]。 此外,外泌體衍生的miR-146a-5p 通過抑制IRAK1/TRAF6 信號通路,降低NF-κB活化和M1 極化,最終減少腦缺血再灌注誘導的組織水腫、細胞死亡、缺血性梗死和功能性運動缺陷[24]。 大量證據強調了miRNA 在缺血性腦卒中炎癥反應中的調節作用,并表明miRNA 在IS 的神經保護中的潛在作用。
2.2 miRNA 抑制細胞凋亡
IS 后會出現大量的細胞凋亡。 小膠質細胞在調節細胞凋亡和神經保護起著關鍵的作用,活化的小膠質細胞產生的外泌體,稱為M2 表型小膠質細胞外泌體(bv2-exosome,BV2-Exo)。 有研究證明,BV2-Exo 衍生的miRNA 在體內外會被神經元吸收,從而減弱氧葡萄糖剝奪(oxygen glucose deprivation,OGD) 后的神經元凋亡[25-27]。 Zhang 等[25]發現Notch1 是miR-137 的直接靶標,miR-137 在BV2-Exo表達升高,miR-137 通過靶向Notch1 信號通路保護神經元免受OGD 引起的神經元凋亡。 Song 等[26]發現泛素特異性蛋白酶14(ubiquitin-specicprotease14,USP14)是miR-124 的直接下游靶標,BV2-Exo 可以通過miR-124 及其下游靶標USP14 減輕缺血性腦損傷和凋亡并促進神經元存活。 其他研究表明,miRNA 還可以通過抑制靶基因或信號通路來發揮抗凋亡作用。 FOXO1 被發現是miR-27a-3p 在HT22細胞中的直接靶基因,miR-27a-3p 拮抗劑可以顯著減輕了I/R 大鼠模型的體內癥狀,而miR-27a-3p 可能是通過靶向FOXO1 抑制I/R 損傷[27]。 此外,來自MSC 的外泌體miR-26a-5p 是通過抑制細胞周期蛋白依賴性激酶(cyclin-dependent kinase,CDK6)來減少小膠質細胞凋亡,從而減輕小鼠的腦缺血再灌注損傷[28]。 源自內皮祖細胞的miR-126 也被證明可以抑制細胞凋亡,miR-126 水平與梗死體積和細胞凋亡呈負相關,與微血管密度呈正相關,上調miR-126 水平對缺氧誘導的神經元凋亡和軸突生長具有保護作用[29]。 而miR-29b-3p 的抗凋亡作用是通過調節PTEN/Akt 信號通路來減輕缺氧缺血性腦損傷的作用[30]。 值得一提的是,MSC 衍生的外泌體miR-145 可以顯著抑制氧糖剝奪再灌注(oxygenglucose deprivation/reoxygenation,OGD/R)后小膠質細胞的凋亡,細胞周期停滯和氧化應激反應,這些是通過miR-145 從MSC 轉移到小膠質細胞中,然后小膠質細胞極化轉變為抗炎M2 表型來完成的[31]。
2.3 miRNA 抑制氧化應激反應
氧化應激反應與I/R 損傷的病理進展密切相關,它是由于ROS 的過量產生或降解不足引起的,在生理條件下,ROS 的產生和清除二者之間會保持平衡,I/R 后這種平衡就會被打破,會表現為ROS產量增加、抗氧化劑消耗增加,最后誘導蛋白質、脂質、DNA 和RNA 的氧化損傷。 因此,在腦缺血后,miRNA 可以通過內源性促氧化酶和抗氧化酶調控ROS 生成來減少這些有害作用[32]。 Chu 等[33]證實,Nrf2 是調節內源性抗氧化劑的重要轉錄因子,而miR-144 是I/R 損傷中Nrf2 的上游調節因子,抑制miR-144 活性可以在翻譯后促進Nrf2/ARE 途徑的抗氧化應激防御,從而緩解了I/R 后的氧化應激反應,促進神經元細胞的修復。 另外,在OGD 模型中,miR-31 的過表達和多囊腎病1 型致病基因(polystic kidney disease 1,PKD1)的沉默減輕了氧化應激誘導的神經元損傷,降低了ROS 水平,同時神經元活力升高。 所以說,miR-31 是通過下調PKD1和JAK/STAT3 途徑來減輕IS 小鼠的炎癥反應以及氧化應激誘導的神經元損傷[34]。 Shi 等[35]證實,肌細胞增強因子2D(myocyte enhancer factor 2D,MEF2D) 是miR-217 的直接靶標,miR-217 通過MEF2D 抑 制 NADH 脫 氫 酶 亞 基 6 ( NADH dehydrogenase subunit 6,ND6)的表達。 MEF2D 的過表達可以減少OGD 后神經元氧化應激反應和ROS的生成,減輕神經元損傷。 非編碼RNA OIP5-AS1與缺血性腦卒中密切相關,上調lncRNA OIP5-AS1可以使miR-186-5p 激活TNF 相關蛋白3(tumor necrosis factor-related protein 3,CTRP3),保護神經元免受I/R 誘導的炎癥和氧化應激反應[36]。 而且lncRNA OIP5-AS1 還可以通過調控miR-155-5p/IRF2BP2 軸來抑制氧化應激和炎癥反應,減輕HMC3 和SH-SY5Y 細胞OGD/R 誘導引起的細胞損傷[37]。 綜上所述,miRNA 可以有效抑制氧化應激反應并為缺血性腦卒中的神經保護提供新的靶向基因。
2.4 miRNA 促進血管新生
血管新生是目前公認的治療IS 的一個潛在靶點,它有助于在IS 后腦組織的神經功能的恢復。 血管新生是通過刺激血管的生長來實現的,血管的生成可使腦灌注趨于穩定,繼而促進神經元的生存,提高大腦可塑性及恢復神經功能[38]。 血管內皮生長因子( vascular endothelial cell growth factor,VEGF)是調控血管生成的關鍵分子之一。 miRNA在VEGF 及相關信號通路中發揮著直接或間接調控血管生成的作用。 Sun 等[39]認為miR-15a/16-1 簇的內皮靶向缺失會減輕卒中后腦梗死和萎縮,并改善缺血性卒中后長期感覺運動和認知障礙。 它通過上調VEGF,纖維細胞生長因子2(fibroblast growth factor receptor 2,FGFR2)及受體VEGFR2 和FGFR1的蛋白表達促進血管重塑和血管生成來增加同側腦的血流量,有利于IS 后的神經恢復。 而miR-107模擬物是通過增強內皮VEGF165/164 的水平促進血管生成來減少缺血性腦梗死并增加半影區域的毛細血管數量。 Fan 等[40]研究證實,抑制miR-377的表達可以促進腦微血管內皮細胞(brain microvascular endothelial cells,BMEC)的毛細管樣管形成和細胞增殖遷移,OGD 后miR-377 調節了早期生長應答因子2(early growth response protein 2,EGR2)的抗炎作用和VEGF 的血管生成作用,所以抑制miR-377 可減少MCAO 大鼠腦梗死體積并抑制炎癥促進血管生成,從而達到神經保護的作用。除此之外,還有其他研究證明miRNA 有益于神經保護,例如miR-27b 轉染增加了大腦中動脈閉塞后半影區的血管生成,從而促進了缺血后的恢復并改善了神經行為表現[41]。 另有研究發現抑制miR-155的過表達后,可以明顯緩解大鼠腦組織的病理損傷,升高網絡微血管密度(microvessel density,MVD)和VEGF 表達水平,并通過PI3K/AKT 信號通路降低炎癥反應,促進血管新生,從而發揮神經元保護作用[42]。 總之,越來越多的研究致力于證實miRNA可以通過促進血管新生來發揮神經元保護作用。
2.5 miRNA 增強神經可塑性和神經發生
miRNA 與其他調控因子共同調控神經可塑性和神經發生的不同過程,并對神經發育有著至關重要的作用[43]。 而且它還可以通過促進神經發生、神經突重塑和存活發揮神經保護作用[44]。 例如來自MSC 的外泌體miRNA 在增強神經可塑性和神經發生有著關鍵的作用。 在體外,用來自MSC 的miR-133b 處理后的OGD 星形膠質細胞可顯著促進胚胎皮質神經元的神經突分支和延長,提高了神經可塑性和功能恢復[45]。 在體內,用miR-133b 治療缺血性腦卒中可顯著改善缺血半暗帶的功能和神經突重塑性[46]。 另外,在大腦中動脈閉塞的大鼠和小鼠模型中,MSC 外泌體miR-17-92 和miR-133b 以及內皮細胞外泌體miR-126 對神經元可塑性和神經恢復都有促進作用[47]。 此外,給予富含miR-17-92 簇的外泌體也可增強卒中后的神經發生和少突膠質發生,這表明miR-17-92 簇參與卒中后的神經可塑性和腦恢復[48]。 除此之外,miR-138 可以延長少突膠質細胞終末分化為髓鞘軸突的能力,它是通過調節NF-κB 活性來調節軸突再生,神經元發育和軸突存活[49]。 諸葛陸杰等[50]研究證明,補陽還五湯可上調miR-199a-5p 的水平,并在腦缺血后14 h 后增加VEGF 和腦源性神經營養因子(brain derived neurotrophic factor, BDNF) 的蛋白表達,來促進大鼠血管新生和神經發生。 (見表1)
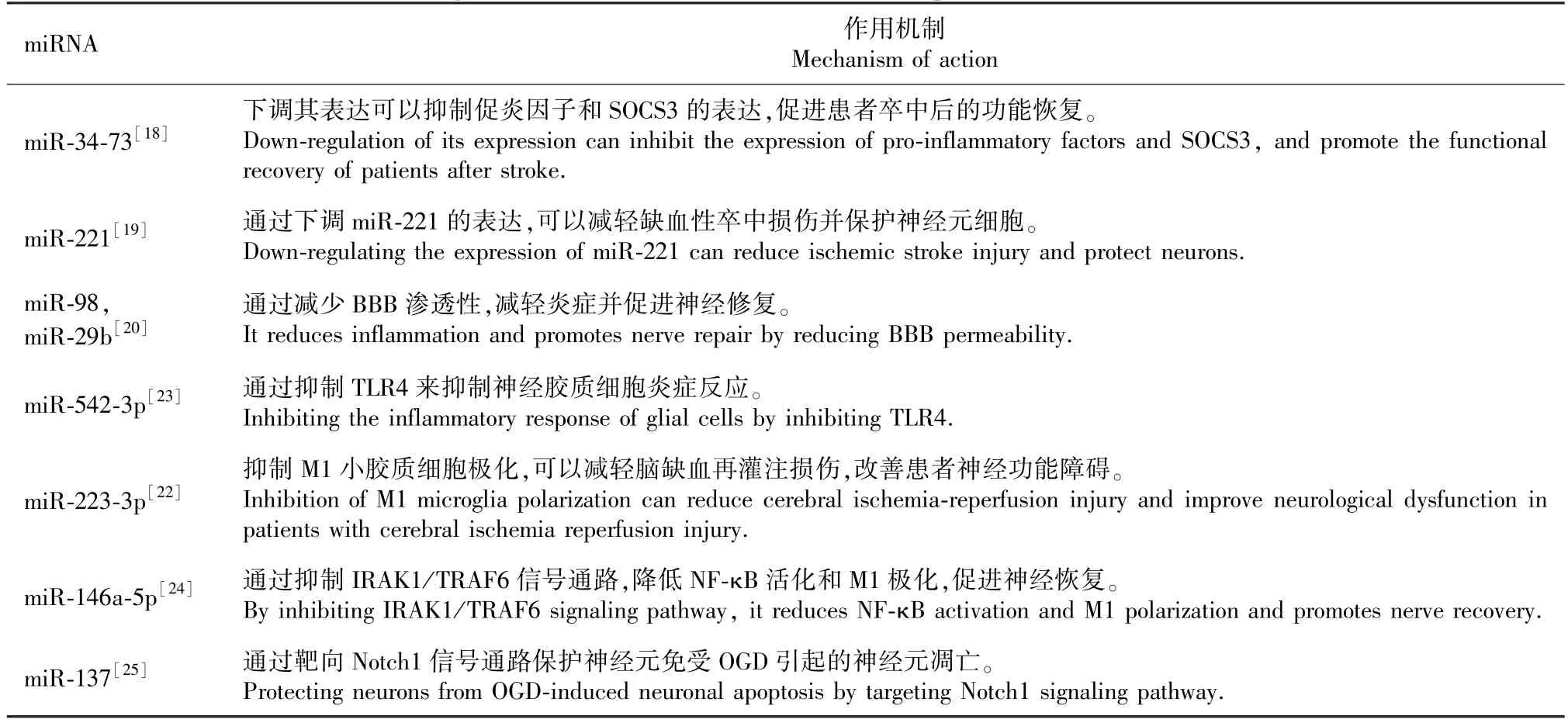
表1 miRNA 在缺血性腦卒中神經保護領域的意義Table 1 Significance of miRNA in the field of neuroprotection in ischemic stroke
3 miRNA 在IS 臨床診斷及治療作用
3.1 miRNA 可作為IS 生物標志物和臨床診斷指標
miRNA 在血液中非常穩定,可以成為潛在的臨床診斷和生物標志物[51]。 有研究表明,miRNA 可作為不同時期卒中患者的生物標志物,在卒中患者中,血漿細胞外的miRNA 和來自血細胞內的miRNA 兩者水平存在差異,在IS 的亞急性和慢性期,血漿中miR-21,miR-221 和miR-145 增加,而miR-210 減少。 在急性卒中中,miR-143-3p、miR-125a-5p、miR-125b-5p 均增加[10]。 眾所周知,引起IS 的發生發展有諸多危險因素,如高血壓、糖尿病、動脈粥樣硬化、年齡、性別和種族等。 所以,與這些危險因素的發病機制相關的幾種miRNA,包括miR-155(高血壓)、miR-33(高脂血癥)、miR-144 和miR-223(糖尿病)以及miR-21、miR-126 和miR-320b(動脈粥樣硬化)都已被提議作為IS 的潛在預測標志物[52]。 miRNA 還可作為缺血性腦卒中后運動和認知恢復的預測生物標志物和診斷指標。 有研究表明,我們可以通過調控miRNA 的表達為運動和認知恢復提供了預測信息[53]。 例如Burlacu 等[54]研究表明miR-132、miR-140-5p、miR-22 和miR-221-3p 在IS 后認知障礙患者中上調,miR-424、miR-100-5p 在預后不良的運動障礙患者中下調。 此外,袁芳等[55]證明,miR-233 與外周血TNF-α、IL-10 水平之間呈正相關,它參與了IS 的整個炎癥過程,miR-233 相對表達量增高提示神經損傷嚴重,miR-233 可能會成為IS 臨床診斷及疾病嚴重程度預測指標。
3.2 miRNA 與IS 治療
目前IS 的急性期和慢性期的治療策略都很有限,我們還需要探求更多的治療方法。 近幾年發現,特異性miRNA 表達的改變有助于缺血性腦卒中的病理生理機制,腦卒中后神經系統難以自我修復,而miRNA 可以促進神經修復,進而有利于IS 后患者預后發展,這說明它有望成為一種潛在的治療方法。 來自小膠質細胞的miR-137 的過表達減少了缺血小鼠的梗死體積和行為缺陷[26]。 來自M2 小膠質細胞的miR-124 通過抑制信號轉導器和轉錄激活因子(signal transducerand activatorof transcription 3,STAT3)的表達,減少了神經膠質細胞瘢痕的形成,促進了IS 的恢復,并緩解了星形膠質細胞的遷移和增殖[56],來源于腦內皮細胞的外泌體miR-126-3p 改變了腦可塑性,促進神經突生長[57]。 而miR-126 的過表達調節了腦損傷后內皮祖細胞(endothelial progenitor cell,EPC)的數量以及增殖、遷移和管形成能力,減少了腦缺血再灌注損傷[58]。這些都可以證明miRNA 可以通過各種機制和途徑來改善神經損傷,促進神經修復,減少I/R 的損傷,這說明miRNA 有望成為IS 的治療方法。
4 展望與總結
近幾年的研究表明,miRNA 作為基因調控因子,可以靶向和調控細胞多個網絡中的數百種蛋白質。 miRNA 的正常表達對大腦的正常發育和功能至關重要,而腦細胞或神經血管系統中miRNA 的失調會增加IS 和其他神經系統疾病的易感性。 腦缺血后,數百種miRNA 的表達在急性和慢性時間點發生顯著改變,這些變化已被證明在IS 的臨床診斷和生物標志物中有著預測的作用。 然而,miRNA 在參與缺血性腦卒中的診斷和治療上仍有很多局限性。盡管miRNA 模擬物和抑制物的神經保護作用已經在體內和體外實驗中得到驗證,但如何使其應用于臨床診斷及如何修飾miRNA 使其在缺血性腦卒中患者神經恢復治療中發揮作用,目前尚不明確。 其次,miRNA 可在各種組織中進行表達,一種miRNA可調控多種基因,一種基因也可被多種miRNA 調控,良好準確的靶向性和安全性應該是未來研究的重點。 未來可針對不同miRNA 及其靶向不同信號通路在缺血性腦卒中的神經保護作用,建立信號通路網絡,并通過基因組學、轉錄組學、蛋白組學、代謝組學等綜合組學技術,去探索miRNA 參與神經保護作用的基因、蛋白質、代謝物和信號通路,從而為IS 提供更多的治療選擇和治療新思路。 綜上所述,miRNA 在調節缺血性腦卒中的神經保護方面發揮了重要作用。 并且miRNA 有著重要的生物學功能,與缺血性腦卒中預后進展緊密相關,miRNA 有望成為缺血性腦卒中的生物標志物和治療靶點。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
