補(bǔ)料策略優(yōu)化促進(jìn)乳酸乳球菌HB03發(fā)酵合成Nisin
2023-12-14 12:45:32熊華儀劉月鋒
食品科學(xué) 2023年22期
熊華儀,陳 曦,劉月鋒,陳 雄,李 沛,王 志,*
(1.湖北工業(yè)大學(xué) 發(fā)酵工程教育部重點(diǎn)實(shí)驗(yàn)室,工業(yè)發(fā)酵省部共建協(xié)同創(chuàng)新中心,湖北 武漢 430068;2.安琪酵母股份有限公司,酵母功能湖北省重點(diǎn)實(shí)驗(yàn)室,湖北 宜昌 443003)
乳酸乳球菌同型乳酸發(fā)酵過(guò)程中合成的乳酸鏈球菌素(Nisin)由34 個(gè)氨基酸組成[1],其能有效抑制革蘭氏陽(yáng)性菌(G+)的生長(zhǎng),是一種安全、綠色的活性抗菌肽,被廣泛應(yīng)用于食品發(fā)酵領(lǐng)域[2]。運(yùn)用基因工程改造菌株有效提高了Nisin合成效率,Papagianni等[3]將黑曲霉A60的磷酸果糖激酶基因pfk13、編碼cAMP依賴(lài)型蛋白激酶pkaC和選擇性氧化酶aox1基因克隆至乳酸乳球菌ATCC11454,提高了其糖酵解活性并促進(jìn)了菌株的氧化呼吸能力,Nisin效價(jià)達(dá)到14 000 IU/mL。Hao Panlong等[4]發(fā)現(xiàn)過(guò)表達(dá)天冬氨酸合成酶基因(asnH)可以使乳酸乳球菌F44在pH 3條件下的存活率提高7 倍,同時(shí)Nisin產(chǎn)量提高57%,達(dá)到5 346 IU/mL。然而基因工程菌應(yīng)用于食品發(fā)酵存在一定的安全風(fēng)險(xiǎn),運(yùn)用發(fā)酵工程策略提高Nisin合成效率仍然具有一定的積極意義。
Nisin合成效率與氨基酸前體或多肽的供給有關(guān)[5],并與乳酸乳球菌生長(zhǎng)存在氮源競(jìng)爭(zhēng)關(guān)系[6]。Nisin含有5 個(gè)脫水氨基酸殘基絲氨酸(serine,Ser)、蘇氨酸(threonine,Thr)和半胱氨酸(cysteine,Cys)合成的硫醚鍵[2],因而Cys的合理供應(yīng)是滿足高效Nisin發(fā)酵的基礎(chǔ)條件之一。另外,乳酸乳球菌厭氧發(fā)酵過(guò)程依然存在氧化脅迫效應(yīng)[7],Gaudu等[8]發(fā)現(xiàn)Cys作為還原劑能顯著增加乳酸乳球菌的存活率并減少氧化脅迫損傷而促進(jìn)細(xì)胞生長(zhǎng)[7]。而有關(guān)Cys對(duì)Nisin發(fā)酵合成和氧化應(yīng)激的影響鮮見(jiàn)報(bào)道。
乳酸乳球菌的蛋白水解體系[9]包括壁膜蛋白酶PrtP、多肽轉(zhuǎn)運(yùn)蛋白DtpP、胞內(nèi)肽酶PepV[10]。該系統(tǒng)的表達(dá)受氮代謝全局調(diào)控子CodY負(fù)控制且主要受異亮氨酸的調(diào)節(jié)[11]。另外,Vido等[12]發(fā)現(xiàn)了不依賴(lài)于CodY的蛋白水解體系調(diào)控機(jī)制,乳酸乳球菌由發(fā)酵轉(zhuǎn)為呼吸代謝可強(qiáng)烈誘導(dǎo)蛋白水解體系中性內(nèi)肽酶(PepO1)和氨肽酶C(PepC)的表達(dá)。
精氨酸(arginine,Arg)在乳酸乳球菌生長(zhǎng)代謝過(guò)程中也發(fā)揮了重要作用,Arg經(jīng)脫亞氨酶(arginine deiminase,ADI)途徑[13]分解,產(chǎn)生銨根和氨甲酰磷酸以及后續(xù)的分解產(chǎn)物CO2、銨根和ATP,這在細(xì)胞應(yīng)對(duì)酸脅迫中發(fā)揮了重要作用[14]。乳酸乳球菌Arg代謝與嘧啶合成代謝也密切關(guān)聯(lián)[15-16],調(diào)控蛋白ArgR阻遏arg操縱子表達(dá),ΔargR菌株中Arg合成基因上調(diào),同時(shí)尿嘧啶核苷酸(uridine monophosphate,UMP)從頭生物合成基因(pyrRPB、carA、pyrEC及pyrKDbF)表達(dá)也上調(diào)[15]。由于肽聚糖合成始于尿苷三磷酸(uridine triphosphate,UTP)和N-乙酰葡萄糖胺-1-P[9],因而UTP的不足也可能導(dǎo)致細(xì)胞肽聚糖合成及生長(zhǎng)受限[17],甚至引起細(xì)胞在脅迫條件下自溶[18],因此Arg在細(xì)胞能量供應(yīng)、酸脅迫應(yīng)激及UTP與肽聚糖合成方面發(fā)揮著重要作用,而有關(guān)Arg對(duì)乳酸乳球菌Nisin發(fā)酵合成效率的影響鮮見(jiàn)報(bào)道。
為了提高Nisin的合成效率,在10 L發(fā)酵罐水平研究碳氮源(糖、Cys、蛋白胨)的補(bǔ)料策略對(duì)Nisin合成的影響,分析Nisin合成關(guān)鍵時(shí)間節(jié)點(diǎn)前后轉(zhuǎn)錄組表達(dá)差異,并結(jié)合胞外氨基酸濃度變化規(guī)律,確定Arg為限制氨基酸,并優(yōu)化補(bǔ)料策略,顯著提高Nisin發(fā)酵合成效率。
1 材料與方法
1.1 材料與試劑
乳酸乳球菌乳酸亞種(Lactococcus lactissubsp.lactisHB03),本實(shí)驗(yàn)室保藏。
蛋白胨、酵母浸粉 安琪酵母股份有限公司;蔗糖(食用級(jí))市售;K2HPO4·3H2O、NaCl、MgSO4·7H2O、MnSO4·H2O、CaCO3、H2SO4、HCl、蒽酮、檸檬酸銨(均為分析純)國(guó)藥集團(tuán)化學(xué)試劑有限公司;Nisin標(biāo)準(zhǔn)品(1 000 IU/mg)美國(guó)Sigma公司。
斜面種子培養(yǎng)基:15 g/L胰蛋白胨,15 g/L酵母浸粉,15 g/L牛肉膏,10 g/L葡萄糖,10 g/L CH3COONa,2 g/L Na2HPO4,2 g/L檸檬酸銨,10 g/L CaCO3,20 g/L瓊脂,pH 6.8。
搖瓶培養(yǎng)基:70 g/L蛋白胨,16 g/L蔗糖,10 g/L酵母浸粉,2 g/L K2HPO4,0.2 g/L MgSO4·7H2O,0.05 g/L MnSO4·H2O,5 g/L檸檬酸銨,6 g/L CaCO3。
10 L發(fā)酵罐培養(yǎng)基:70 g/L蛋白胨,16 g/L蔗糖,10 g/L酵母浸粉,2 g/L K2HPO4,0.2 g/L MgSO4·7H2O,0.05 g/L MnSO4·H2O,5 g/L檸檬酸銨。
1.2 儀器與設(shè)備
SPX-150D恒溫培養(yǎng)箱 上海博訊實(shí)業(yè)有限公司醫(yī)療設(shè)備廠;HNV-211B搖床 天津市歐諾儀器儀表有限公司;YXQ-75SII立式蒸汽滅菌鍋 上海博訊醫(yī)療生物儀器股份有限公司;Ultimate 3000高效液相色譜儀 美國(guó)Agilent公司;S-10生物傳感器儀 西爾曼科技有限公司;C J-2 D 凈化操作臺(tái) 天津泰斯特儀器有限公司;10 L機(jī)械攪拌通風(fēng)發(fā)酵罐 上海保興生物設(shè)備有限公司;V-1300分光光度計(jì) 上海美析儀器有限公司。
1.3 方法
1.3.1 菌株活化
將-80 ℃冰箱保存的乳酸乳球菌HB03甘油管接種于斜面種子培養(yǎng)基的平板,于30 ℃培養(yǎng)箱24 h后轉(zhuǎn)接茄子瓶,30 ℃培養(yǎng)24 h,無(wú)菌操作加入60 mL無(wú)菌水,并用無(wú)菌竹簽將菌苔挑起混合于無(wú)菌水中,得到種子液,備用。
1.3.2 搖瓶培養(yǎng)及10 L發(fā)酵罐實(shí)驗(yàn)
搖瓶培養(yǎng):裝液量為50 mL/250 mL發(fā)酵培養(yǎng)基,調(diào)節(jié)初始pH 6.8~7.0,121 ℃滅菌20 min,每瓶按OD600nm=0.1接種。放置于30 ℃恒溫?fù)u床間歇振蕩培養(yǎng)12 h(每小時(shí)工作30 s,轉(zhuǎn)速200 r/min,隨后調(diào)整轉(zhuǎn)速為0 r/min)。
10 L罐分批發(fā)酵:10 L發(fā)酵罐中裝量7 L發(fā)酵培養(yǎng)基,調(diào)整初始pH 6.8~7.0,121~123 ℃滅菌25 min;接種60 mL種子液,30 ℃、100 r/min不通氣培養(yǎng)20~24 h。
1.3.3 分析檢測(cè)
1.3.3.1 生物量(以細(xì)胞數(shù)計(jì))測(cè)定
稀釋涂布平板法[19]。
1.3.3.2 總糖測(cè)定
參考硫酸-蒽酮法[20]。
1.3.3.3 乳酸測(cè)定
生物傳感儀[21]測(cè)定。
按式(1)計(jì)算:
1.3.3.5 Nisin效價(jià)測(cè)定
采用高效液相色譜法[22]測(cè)定。樣品處理:發(fā)酵液用鹽酸稀釋至pH 2.0~2.7[23],12 000 r/min離心20 min,經(jīng)0.45 μm濾膜過(guò)濾后備用。Nisin標(biāo)準(zhǔn)方程繪制:將0.25 g Nisin標(biāo)準(zhǔn)品稱(chēng)取至50 mL容量瓶,使用0.01 mol/L鹽酸溶液(pH 2.0)定容,得到標(biāo)準(zhǔn)液(5 000 IU/mL),經(jīng)0.45 μm濾膜過(guò)濾備用。色譜條件:流動(dòng)相:水-乙腈(81∶19,V/V)混溶后加入0.05%三氟乙酸;流速1 mL/min;檢測(cè)波長(zhǎng)200 nm;柱溫40 ℃;進(jìn)樣量20 μL。
1.3.3.6 H2O2濃度的測(cè)定
采用索萊寶公司試劑盒,按說(shuō)明書(shū)方法檢測(cè)H2O2濃度。
1.3.3.7 游離氨基酸的測(cè)定
參考文獻(xiàn)[24]的方法進(jìn)行測(cè)定。
1.3.4 轉(zhuǎn)錄組數(shù)據(jù)分析
取發(fā)酵15、18 h的發(fā)酵液20 mL于50 mL離心管中,4 ℃、12 000 r/min離心5 min,無(wú)菌操作留取菌體沉淀并-80 ℃保存。轉(zhuǎn)錄組測(cè)序由美吉生物科技有限公司完成。轉(zhuǎn)錄表達(dá)定量分析每百萬(wàn)讀取次數(shù)中某個(gè)轉(zhuǎn)錄本占比(transcripts per million,TPM)值按照式(2)計(jì)算[25]:
式中:R、L為需計(jì)算基因的read counts和基因長(zhǎng)度/kb;Ri、Li(i=1,2,…,n)為樣品中第i個(gè)基因的read counts和基因長(zhǎng)度/kb。
1.4 數(shù)據(jù)分析
2 結(jié)果與分析
2.1 搖瓶水平Cys添加促進(jìn)Nisin合成
Nisin是由34 個(gè)氨基酸組成的多肽,含有5 個(gè)硫醚橋(由Cys、Thr、Ser生成)[2]。因而,Cys是Nisin的重要前體氨基酸。另外,其作為含硫氨基酸還具有抵御氧化脅迫的作用[8]。因此,搖瓶水平考察了對(duì)數(shù)中期(16 h左右)添加不同濃度Cys對(duì)菌株HB03合成Nisin的影響,如圖1所示。16 h添加0.15~0.6 mmol/L的Cys在24 h的Nisin效價(jià)較對(duì)照(未添加Cys)分別提升7.9%~20%。顯然,Cys的添加促進(jìn)了Nisin的生成。Cys添加量為0.6 mmol/L時(shí)Nisin效價(jià)為6 457 IU/mL。但隨著Cys的添加量增多Nisin效價(jià)呈下降趨勢(shì),當(dāng)添加量為0.75 mmol/L和0.9 mmol/L時(shí),Nisin效價(jià)分別為6 140 IU/mL和5 862 IU/mL,也較對(duì)照提高14.8%和9%。綜上Cys添加量0.6 mmol/L為最優(yōu),提高效價(jià)幅度達(dá)到20%。
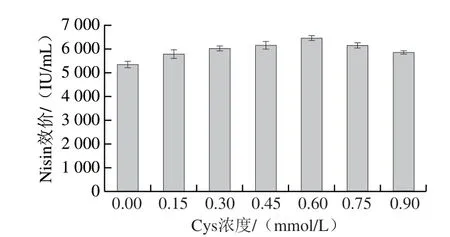
圖1 搖瓶中Cys濃度對(duì)Nisin合成的影響Fig.1 Effect of Cys concentration on nsin production in shake flask culture
2.2 10 L發(fā)酵罐水平Cys添加促進(jìn)Nisin合成
雖然發(fā)酵過(guò)程是靜置(搖瓶)和未通空氣的攪拌狀態(tài)(10 L罐),但是發(fā)酵體系存在空氣接觸過(guò)程,乳酸乳球菌通過(guò)乳酸氧化酶LOX途徑[26-27]、NADH氧化途徑[28]等把氧氣消耗,但同時(shí)產(chǎn)生了H2O2,而乳酸乳球菌是接觸酶陰性菌株,因而細(xì)胞生長(zhǎng)過(guò)程伴隨著氧化脅迫發(fā)生[29-30]。由于Nisin合成與生長(zhǎng)偶聯(lián)[7],為了降低氧化脅迫效應(yīng)以及維持細(xì)胞生長(zhǎng)效率,在罐上將Cys的添加策略調(diào)整為對(duì)數(shù)期及其穩(wěn)定期(11、13.5、18、23.5 h每次添加0.15 mmol/L,總計(jì)0.6 mmol/L)。10 L罐水平Cys添加策略(終濃度0.6 mmol/L,策略1#)對(duì)乳酸乳球菌HB03生長(zhǎng)代謝的影響如圖2所示。

圖2 Cys補(bǔ)料策略1#對(duì)Nisin合成的影響Fig.2 Effect of Cys supplementation strategy 1# on nisin synthesis
細(xì)胞生長(zhǎng)至18 h達(dá)到峰值(1.43×1010CFU/mL),而后自溶至21 h的1.16×1010CFU/mL并維持穩(wěn)定。總糖由15.8 g/L緩慢消耗至14.62 g/L(9 h)后迅速消耗至1.75 g/L(21 h)。Nisin合成于15 h達(dá)到峰值(6 993 IU/mL)后迅速分解。同時(shí),細(xì)胞對(duì)數(shù)早期(9 h)H2O2為0.87 mmol/L,這與文獻(xiàn)[30-31]氧化脅迫伴隨著細(xì)胞生長(zhǎng)過(guò)程的報(bào)道一致。前9 h細(xì)胞生長(zhǎng)啟動(dòng)較慢(糖耗也較低)可能也與細(xì)胞調(diào)整代謝以適應(yīng)氧化脅迫過(guò)程有關(guān)。在細(xì)胞對(duì)數(shù)生長(zhǎng)早期(11 h)添加Cys(0.15 mmol/L)使H2O2迅速降至0.18 mmol/L,在15 h降至0 mmol/L并維持至發(fā)酵結(jié)束。說(shuō)明Cys少量多次添加方式對(duì)消除H2O2有效。
Cys消除H2O2使細(xì)胞生長(zhǎng)加速,18 h細(xì)胞數(shù)為1.43×1010CFU/mL,平均生長(zhǎng)速率為7.9×108CFU/(mL·h),是前9 h生長(zhǎng)速率(3×108CFU/(mL·h))的2.6 倍。Nisin合成偶聯(lián)細(xì)胞生長(zhǎng)至15 h達(dá)到峰值(6 993 IU/mL),15~18 h細(xì)胞繼續(xù)生長(zhǎng),但Nisin處于被降解的狀態(tài),也印證了Nisin合成與細(xì)胞生長(zhǎng)對(duì)氮源存在競(jìng)爭(zhēng)關(guān)系的報(bào)道[10]。此時(shí),Nisin可能作為一種胞外多肽的儲(chǔ)備分子,其被降解可能也是細(xì)胞維持活性的一種生存策略。由于15 h后H2O2濃度為0 mmol/L,因而Nisin的降解可能與其他未知因素有關(guān),并調(diào)整Cys補(bǔ)加策略為:11 h和13.5 h分別加入0.15 mmol/L Cys。
2.3 Nisin對(duì)數(shù)后期降解前后轉(zhuǎn)錄組水平基因表達(dá)差異分析
1#策略發(fā)酵15~18 h期間,Nisin以221.6 IU/(mL·h)的速率迅速降解,但細(xì)胞數(shù)卻提高了30%,達(dá)到了1.43×1010CFU/mL。即15~18 h是Nisin合成和細(xì)胞生長(zhǎng)競(jìng)爭(zhēng)的關(guān)鍵節(jié)點(diǎn),因此,在轉(zhuǎn)錄組水平研究了15~18 h相關(guān)基因轉(zhuǎn)錄差異,包括肽酶相關(guān)基因、UMP從頭合成途徑基因、肽聚糖合成相關(guān)基因等,其顯著差異基因如圖3所示。
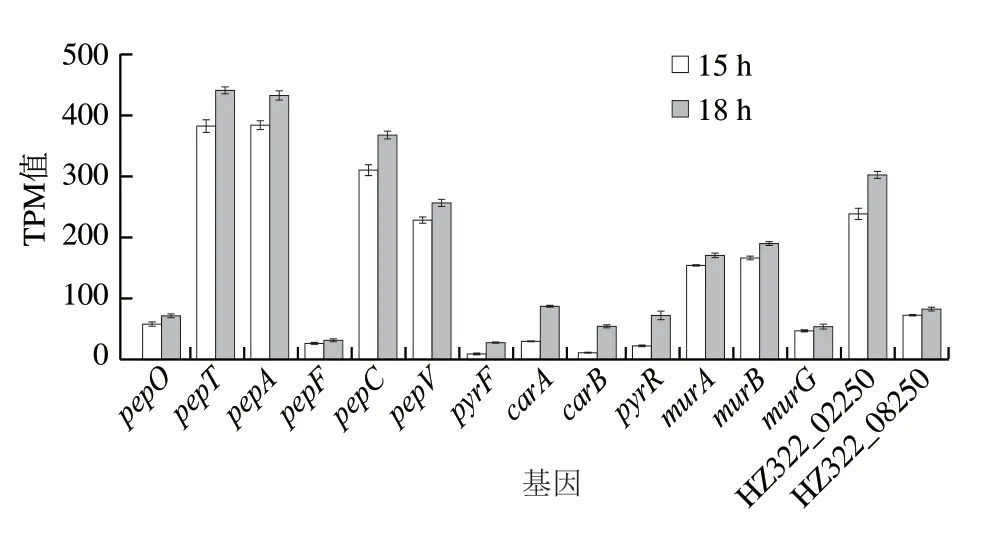
圖3 15 h與18 h相關(guān)基因表達(dá)差異Fig.3 Differential expression of related genes at 15 and 18 h
pepO、pepT、pepA、pepF、pepC、pepV等胞內(nèi)肽酶表達(dá)相關(guān)基因分別上調(diào)12%~24%,說(shuō)明發(fā)酵15~18 h肽酶表達(dá)上升,強(qiáng)化了蛋白、多肽(含Nisin)的分解以用于生長(zhǎng)。同時(shí),18 h細(xì)胞UMP從頭合成途徑合成基因(pyrF、carA、carB、pyrR等)顯著上調(diào)至2~4 倍,而18 h后細(xì)胞大量自溶,這可能與UMP合成(或合成前體)不足有關(guān)。而肽聚糖合成相關(guān)基因(murA、murB、murG等)以及與肽聚糖聚合的LysM蛋白相關(guān)基因(HZ322_08250、HZ322_02250)表達(dá)也均上調(diào)10%~27%。這與文獻(xiàn)[15,32]報(bào)道一致。說(shuō)明UMP合成前體氨甲酰磷酸可能處于限制狀態(tài),使得UMP、UTP供應(yīng)不足,進(jìn)而影響肽聚糖前體二磷酸尿苷葡萄糖的合成。細(xì)胞通過(guò)大幅上調(diào)UMP合成途徑與肽聚糖合成相關(guān)基因以期滿足細(xì)胞需求。
氨甲酰磷酸在Arg ADI途徑中產(chǎn)生[13],說(shuō)明18 h不僅氨甲酰磷酸可能缺乏,同時(shí)Arg也處于限制狀態(tài)。因而,細(xì)胞為尋求Arg的及時(shí)補(bǔ)充,肽酶相關(guān)表達(dá)也上調(diào)12%~24%。這與含硫氨基酸或Arg的缺乏導(dǎo)致肽酶表達(dá)增強(qiáng)[9]的報(bào)道一致,細(xì)胞需要大量分解多肽以響應(yīng)含硫氨基酸的缺乏,以及通過(guò)ADI途徑降解Arg緩解UMP合成壓力和應(yīng)對(duì)酸脅迫壓力。Nisin作為一種胞外的多肽“儲(chǔ)備分子”,在細(xì)胞存在生存壓力而加強(qiáng)肽酶表達(dá)時(shí)必然會(huì)被大量的降解。因此15~18 h細(xì)胞仍能持續(xù)生長(zhǎng),但Nisin效價(jià)下降。
2.4 基于轉(zhuǎn)錄組分析的Nisin補(bǔ)料策略優(yōu)化
Nisin合成與細(xì)胞生長(zhǎng)對(duì)氮源存在競(jìng)爭(zhēng)關(guān)系[10]說(shuō)明氮源的供應(yīng)不足,因而,發(fā)酵過(guò)程中補(bǔ)加氮源(蛋白胨等)能緩解細(xì)胞對(duì)氮源的需求,也可作為前體直接用于Nisin合成;而Arg具有氮源和能源分子的雙功能,也是需要考慮的因素。另外,蔗糖作為發(fā)酵過(guò)程唯一的碳源,適宜的流加也是必要的。補(bǔ)加碳氮源時(shí)間以及氨基酸對(duì)Nisin合成影響的設(shè)計(jì)如表1所示。

表1 10 L發(fā)酵罐上補(bǔ)料策略設(shè)計(jì)Table 1 Design of feeding strategy in a 10 L bioreactor
在補(bǔ)加Cys(11、13.5 h分別加入0.15 mmol/L Cys)的前提下,考慮到細(xì)胞自溶和Nisin降解同時(shí)發(fā)生,以及策略1# 9~18 h的平均糖耗速率為1.42 g/(L·h),確定策略2#為15 h補(bǔ)入蛋白胨(15 g/L)(1.5 g/(L·h)),其對(duì)乳酸乳球菌HB03代謝以及Nisin合成的影響如圖4所示。
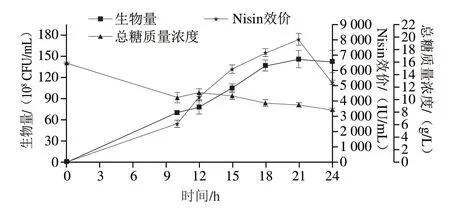
圖4 策略2#對(duì)細(xì)胞生長(zhǎng)代謝的影響Fig.4 Effect of strategy 2# on cell growth and nisin production in a 10 L bioreactor
Nisin 迅速合成至21 h 效價(jià)達(dá)到峰值,為8 042 IU/mL,較1#策略提高15%,后迅速分解。其_vq為383 IU/(mL·h)。10 h開(kāi)始補(bǔ)糖以及15 h補(bǔ)加的蛋白胨使生物量在18~24 h穩(wěn)定,總糖由15.8 g/L降至10.33 g/L(10 h),10~24 h殘?zhí)欠€(wěn)定維持在8~11 g/L之間。
以上結(jié)果說(shuō)明,碳氮源的持續(xù)補(bǔ)加為乳酸乳球菌HB03生長(zhǎng)以及Nisin合成起著積極作用,15 h補(bǔ)入蛋白胨使Nisin持續(xù)合成,但21 h Nisin合成達(dá)到峰值后依然以較快的速率降解。考慮到蛋白胨體系中可能存在氨基酸比例不平衡的問(wèn)題,因此對(duì)策略2#細(xì)胞生長(zhǎng)和Nisin主合成期(10~24 h)胞外氨基酸進(jìn)行了測(cè)定,如表2所示。

表2 策略2#中胞外14 種氨基酸隨時(shí)間的變化Table 2 Changes in concentrations of 14 extracellular amino acids in strategy 2#
Arg發(fā)酵過(guò)程均未檢出,說(shuō)明Arg是一種嚴(yán)重缺少的限制氨基酸。Arg參與ADI途徑產(chǎn)能、緩解胞內(nèi)pH值下降引起的酸脅迫、滿足細(xì)胞生長(zhǎng)、參與嘧啶合成[33]。因而,Arg的嚴(yán)重不足導(dǎo)致嘧啶途徑表達(dá)的大量上調(diào),與轉(zhuǎn)錄組基因差異結(jié)果一致,需要適量補(bǔ)充。
另外,胱氨酸發(fā)酵全過(guò)程質(zhì)量分?jǐn)?shù)穩(wěn)定維持0.015%~0.025%,氧化條件下Cys會(huì)轉(zhuǎn)化為胱氨酸,考慮添加的Cys可能不足,所以在策略2#的基礎(chǔ)上17、19.5 h分別加入0.15 mmol/L Cys。12~15 h為614 IU/(mL·h),15~18 h為356 IU/(mL·h),從Nisin合成速率看12~15 h速率更快,所以將蛋白胨(15 g/L)補(bǔ)充時(shí)間提前至12 h,形成策略3#(11、13.5、17、19.5 h分別加入0.15 mmol/L Cys,10 h補(bǔ)蔗糖(1.5 g/(L·h))以及12 h補(bǔ)充蛋白胨(15 g/L)。其對(duì)乳酸乳球菌HB03代謝以及Nisin合成的影響如圖5所示。
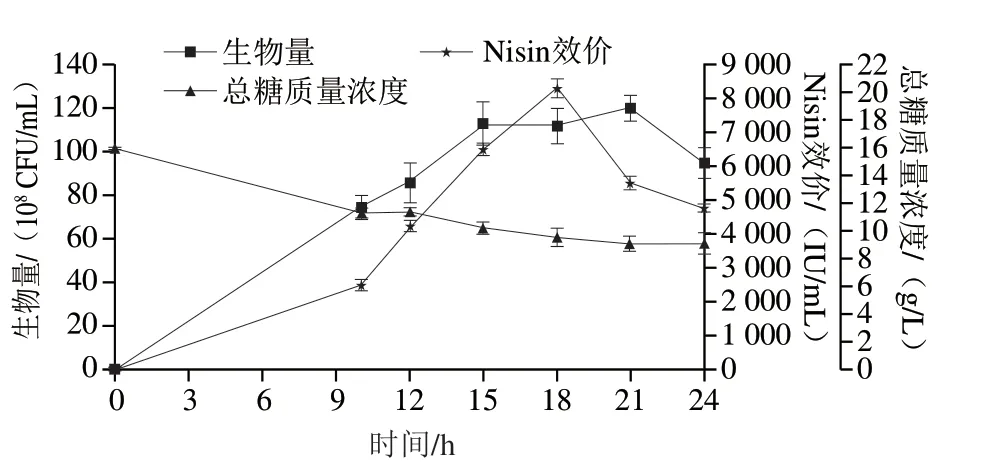
圖5 策略3#對(duì)細(xì)胞生長(zhǎng)代謝的影響Fig.5 Effect of strategy 3# on cell growth and nisin production in a 10 L bioreactor
細(xì)胞在10 h已經(jīng)進(jìn)入對(duì)數(shù)期,持續(xù)生長(zhǎng)至15 h穩(wěn)定后,于21 h生物量達(dá)到峰值,為1.20×1010CFU/mL,為1#策略的84%,后自溶。殘?zhí)怯? h的15.8 g/L迅速下降至10 h的11.32 g/L,10~24 h緩慢利用至9.18 g/L。Nisin合成在18 h達(dá)到峰值,為8 311 IU/mL,較1#策略提高18.8%,后迅速降解。其為461.7 IU/(mL·h),較2#策略提高20%。3#策略與2#策略相比,Nisin峰值效價(jià)差別較小,但達(dá)到Nisin效價(jià)峰值的時(shí)間點(diǎn)有所偏移,提前加入蛋白胨提前了Nisin的合成峰值時(shí)間。顯然,加大含硫氨基酸的濃度以及提前蛋白胨的添加時(shí)間對(duì)于細(xì)胞來(lái)說(shuō)雖然效價(jià)無(wú)明顯提升,卻提高了Nisin合成的速率。然而Nisin效價(jià)達(dá)到最高后仍然迅速分解,18~24 h Nisin以589.3 IU/(mL·h)的速率迅速降解。
細(xì)胞通過(guò)ADI途徑獲得能量、升高胞內(nèi)pH值。ADI中間產(chǎn)物-氨甲酰磷酸可以作為前體進(jìn)入嘧啶代謝,進(jìn)而可能緩解細(xì)胞肽聚糖合成不足導(dǎo)致的細(xì)胞自溶以及肽酶大量表達(dá)等影響。因此需要確定Arg對(duì)Nisin合成的影響,如圖6所示。

圖6 搖瓶中Arg質(zhì)量濃度對(duì)Nisin合成的影響Fig.6 Effect of different concentrations of Arg on nisin production in shake flask culture
搖瓶水平21 h添加1.5、1.8、2.5、3 g/L的Arg,在24 h較對(duì)照(未添加Arg)Nisin效價(jià)分別提升16%、23%、35%、29%。顯然,添加Arg顯著促進(jìn)了Nisin的生成,以2.5 g/L最佳。在策略3#的基礎(chǔ)上12~21 h加入Arg 0.25 g/(L·h),形成策略4#。其對(duì)乳酸乳球菌HB03代謝以及Nisin合成的影響如圖7所示。
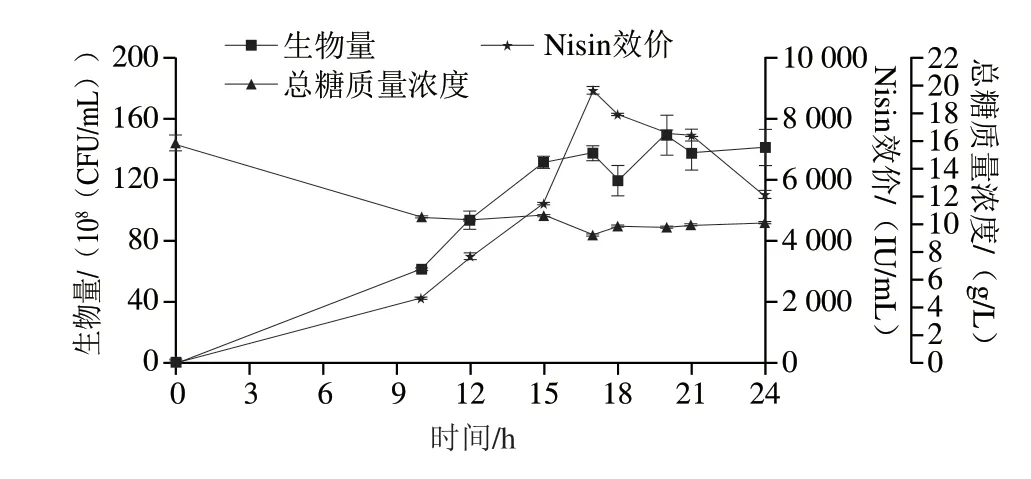
圖7 策略4#對(duì)細(xì)胞生長(zhǎng)代謝的影響Fig.7 Effect of strategy 4# on cell growth and nisin production in a 10 L bioreactor
細(xì)胞在10 h已經(jīng)進(jìn)入對(duì)數(shù)期,持續(xù)生長(zhǎng)至15 h達(dá)到1.32×1010CFU/mL,后15~24 h維持穩(wěn)定,其中生物量峰值為1.50×1010CFU/mL(20 h),較策略3#提高25%。殘?zhí)怯? h 15.8 g/L迅速下降至10 h 10.48 g/L,補(bǔ)糖后10~24 h保持在9~10 g/L之間。Nisin合成在17 h達(dá)到峰值,為8 963 IU/mL,較策略3#提高8%,17~21 h的Nisin以342.5 IU/(mL·h)速率降解,為策略3#的58%。而21 h后Arg不再流加,21~24 h Nisin降解速率恢復(fù)至686 IU/(mL·h),較17~21 h提高100%。說(shuō)明Arg可大幅度延緩Nisin的降解速率,后續(xù)可增大添加量以持續(xù)促進(jìn)Nisin合成和細(xì)胞生長(zhǎng)。
3 結(jié)論
乳酸乳球菌HB03發(fā)酵合成Nisin過(guò)程存在氧化脅迫(9 h的H2O2為0.87 mmol/L),10 L罐發(fā)酵過(guò)程分4 次添加Cys(11、13.5、18、23.5 h分別加入0.15 mmol/L)降低了H2O2濃度,加速了細(xì)胞的生長(zhǎng),促進(jìn)了Nisin合成,達(dá)到6 993 IU/mL。通過(guò)分析胞外氨基酸消長(zhǎng)規(guī)律和肽水解酶系、UMP從頭合成、肽聚糖合成代謝在轉(zhuǎn)錄組水平的表達(dá)差異,確定了對(duì)數(shù)中后期Nisin合成限制因素為碳氮源及Arg供應(yīng)不足,并通過(guò)氨基酸單因素和碳氮源補(bǔ)加優(yōu)化,確定了10 L發(fā)酵罐水平優(yōu)化的補(bǔ)料策略。在此條件下Nisin效價(jià)達(dá)到8 963 IU/mL,比只補(bǔ)加Cys發(fā)酵效價(jià)(6 993 IU/mL)提高了28.2%,也比優(yōu)化前效價(jià)(5 349 IU/mL,資料未顯示)提高了67%。
猜你喜歡
教學(xué)考試(高考化學(xué))(2021年2期)2021-05-30 06:15:52
小讀者(2021年2期)2021-03-29 05:03:48
中學(xué)生數(shù)理化·高一版(2020年3期)2020-04-21 08:03:20
中學(xué)生數(shù)理化(高中版.高考理化)(2020年2期)2020-04-21 05:32:50
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
小學(xué)生作文(低年級(jí)適用)(2019年9期)2019-10-08 08:37:10
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
數(shù)學(xué)大世界(2018年1期)2018-04-12 05:39:14

- 食品科學(xué)的其它文章
- 基于大氣壓化學(xué)電離的GC-MS/MS法測(cè)定動(dòng)物源性食品中8 種酰胺類(lèi)除草劑殘留
- 同位素稀釋-超高效液相色譜-串聯(lián)質(zhì)譜法測(cè)定雞蛋中硝基咪唑類(lèi)藥物及其代謝物
- 基于代謝組學(xué)分析鐵觀音烏龍茶包揉過(guò)程中代謝物動(dòng)態(tài)變化
- 基于GC-MS技術(shù)分析燕山板栗果實(shí)糖組分及其含量特征
- 基于GC-IMS結(jié)合化學(xué)計(jì)量法分析燒烤過(guò)程中溫度對(duì)牦牛肉風(fēng)味物質(zhì)的影響
- 快速濾過(guò)型凈化結(jié)合液相色譜-蒸發(fā)光散射法檢測(cè)茶葉中5 種糖類(lèi)成分