欖李開花生物學和繁育系統
2024-01-02 11:29:14蔡艷清陳玉軍鄧創發黃烈健
植物研究 2024年1期
關鍵詞:系統
蔡艷清 陳玉軍 李 玫 鄧創發 黃烈健*
(1.中國林業科學研究院熱帶林業研究所,廣州 510520; 2.南京林業大學,南京 210037)
紅樹林生態系統是熱帶和亞熱帶海岸的主要濕地類型之一,在河口生態系統中扮演重要角色[1]。紅樹林生存于高鹽度、極端潮汐、強風、高溫以及泥濘的厭氧土中[2],其可以從海洋和大陸獲取養分,是豐富的碳源[3],也是許多大陸和海洋生物的棲息地[4]。20 世紀50 年代以來,我國的紅樹林遭受了嚴重的人為破壞,面積不斷減少[5]。我國37種原生紅樹植物和半紅樹植物近一半的種類處于不同程度的珍稀瀕危狀態[5],導致紅樹植物生物多樣性不斷下降。保護紅樹植物生物多樣性工作迫在眉睫。
欖李(Lumnitzera racemosa)是使君子科(Combretaceae)欖李屬(Lumnitzera)真紅樹植物,常綠灌木或小喬木,一般生長于高潮或大潮可淹及的泥沙灘[6]。據《中國植物志》[7]記載,欖李分布于東非熱帶、馬達加斯加、亞洲熱帶、大洋洲北部和波利尼西亞至馬來西亞;欖李在我國主要天然分布于海南、廣東、廣西、香港及臺灣等地區,最北引種至福建。欖李天然林下更新速度慢[8],目前的保存數量正在減少,《廣東省紅樹林保護修復專項行動計劃實施方案》[9]將其列為廣東省瀕危植物,并提出加強欖李等珍稀瀕危物種的搶救性保護修復。以往關于欖李的研究主要集中在生態學[10]、葉片結構[11]、化學計量特征[12]、木材解剖[13]、遺傳多樣性[8]、藥用價值[14]和化學價值[15-16]等方面,而關于其開花生物學和繁育系統方面的報道則缺乏系統的梳理。因此,開展欖李的開花生物學以及繁育系統方面相關研究,對揭示其瀕危機制以及開展相關保護行動,具有重要意義。
1 材料與方法
1.1 試驗地自然地理概況
研究地點為廣東省珠海淇澳擔桿島省級自然保護區(22°25′38.72″N,113°37′44.3″E),位于高潮帶,大中潮時期可淹。該區域為南亞熱帶海洋性氣候,年平均氣溫為22.4 ℃,年降水量為1 964.4 mm,選擇紅樹林保護區內10 年生的11 株欖李(引種地為海南)作為研究對象,株距大約1.5 m×2.0 m,周圍生長的紅樹植物主要有老鼠簕(Acanthus ilicifolius)、桐花樹(Aegiceras corniculatum)、鹵蕨(Acrostichum aureum)、秋茄樹(Kandelia obovata)、銀葉樹(Heritiera littoralis)等,研究時間為2022年5~9月。
1.2 開花動態及花部特征觀察
參照Dafni[17]方法,從群體、花序、單花水平進行開花動態觀察以及對花部綜合特征進行觀測。花序及花部綜合特征觀測主要包括:花序長度、每花序上小花數目、花總長、花絲長度、每花雄蕊數目、花柱長度、子房長度、每子房胚珠數目、花冠直徑、花瓣長、花瓣寬、每花花瓣數目,使用精度為0.01 mm 的電子數顯游標卡尺進行花序長度及花部器官的測量。
1.3 花粉活力測定
1.3.1 花粉離體培養
為篩選最佳花粉萌發條件,于欖李盛花期,選取當天上午開放的花朵,采用液體培養基法[18],設置不同濃度硼酸、蔗糖組合(0.10、0.20、0.30 g·L-1硼酸,5.00、10.00 、30.00 、50.00、100.00 g·L-1蔗糖),以檢測硼酸、蔗糖梯度處理下對欖李花粉萌發的影響。以上花粉萌發試驗均在溫度27~28 ℃、空氣濕度70%~80%環境條件下進行,每處理重復3 次試驗,每重復隨機選取3 個視野,每視野觀察花粉數量不少于30 粒,每重復試驗總花粉數量不少于100粒,培養12 h后,光學顯微鏡(40倍物鏡)下觀察花粉萌發情況。花粉管長度大于1倍花粉粒直徑,視為花粉萌發標準,統計花粉萌發率并以此評估花粉活力:
花粉萌發率=(平均每視野萌發花粉數量/平均每視野花粉數量)×100% (1)
1.3.2 花粉活力檢測
采用最佳花粉離體萌發處理,分別收集第1~3 天的花粉,進行花粉活力測定,各重復3 次試驗。
1.4 柱頭可授性測定
選取開花前2 天、前1 天,以及開花當天、第2天、第3 天、第4 天、第5 天(分別以A2d、A1d、B1d、B2d、B3d、B4d、B5d表示)的花朵各30朵,采用Dafni 等[17]方法,用尖頭鑷子將花柱取下,放置在凹面載玻片上,滴入適量3%過氧化氫溶液,靜置30 s后于光學顯微鏡(10 倍物鏡)下觀察,若柱頭周圍有氣泡產生,則證明柱頭具有活性,氣泡越多柱頭可授性越強,反之不產生氣泡,柱頭無活性,記錄具有可授性柱頭數目并拍照記錄。
1.5 繁育系統
1.5.1 單花花粉量及花粉-胚珠比
在天氣晴朗的上午,09:00~10:00選取花藥尚未爆裂的欖李花朵20朵,放入45 mL 離心管帶回,將每朵花全部花藥在未開裂時用尖頭鑷子剝下,裝入1.5 mL 離心管中,室內放置待花藥完全開裂后,滴入1.5 mL 蒸餾水,蓋好蓋子搖晃3~4 min,得到花粉懸浮液,使用移液槍吸取1.5 μL 滴于載玻片上,光學顯微鏡(10 倍物鏡)下觀察統計花粉數量,重復10 次,算出花粉平均數量,花粉平均數量乘以1 000,即得到每朵花的花粉量,同時用手術刀沿著小苞片位置縱切,記錄每朵花的胚珠數量。用該花朵的單花花粉量除以對應單花胚珠數量計算得出花粉-胚珠比(P/O)。根據Cruden[19]標準評判繁育系統類型。
1.5.2 雜交指數(OCI)
采用花部數量特征觀測數據以及單花盛花期開花特征觀察結果,計算雜交指數(OCI)。根據Dafni[17]的標準判斷欖李的繁育系統類型。具體評判標準為:(1)花朵直徑(d)為1 mm 記為0,1 mm
1.5.3 授粉試驗
隨機選取3株欖李,進行授粉試驗。①自然授粉(對照組):不做任何授粉處理,不套袋。②人工自花授粉:將同一花序上的其他花苞掐掉,在開花前1 天,只留1 朵花苞進行套袋處理,待花開時用自花的花粉授粉,套袋。③人工同株異花授粉:開花前1 天去瓣,去雄,套袋處理,待2 d 后授于同株花異花的花粉,套袋。④人工異株異花授粉:開花前1 天去瓣,去雄,套袋處理,待2 d 后授于異株花的花粉,套袋。⑤無融合生殖:在花開放前對花去瓣后去雄,套袋。每處理套袋40 朵花,重復3 次試驗,授粉15 d 后將硫酸紙袋摘下,待果實成熟后摘取,帶回,統計結實率(結實率=結果數量/試驗花朵數量×100%)、敗育率(敗育率=空胚或胚敗育的果實數量/試驗花朵數量×100%)。
1.6 數據分析
用Excel 2019軟件進行數據處理和圖表制作,用SPSS 25.0 進行數據處理和方差分析,以鄧肯(Duncan)法進行多重比較評價不同蔗糖、硼酸梯度組合處理下對花粉萌發、不同授粉方式對結實率和敗育率差異的顯著性,結果以平均值±標準差表示。
2 結果與分析
2.1 開花生物學特征
2.1.1 群體、花序開花動態
2022 年4 月25 日開始有花芽產生,從葉腋內爆出,2022 年4 月25 日至5 月16 日處于低溫狀態。5 月16 日有2 株欖李開花,5 月17 日開始氣溫逐漸升高,共有3 株欖李開花,大于25%植株開花,群體進入始花期。5 月18 日溫度升高,位于向陽地帶的欖李花朵大量開放,共有6 株欖李開花,大于50%植株開花,進入群體盛花期,位于庇蔭環境下的欖李尚未進入花期,時間間隔6~7 d開始開花。5 月31 日最后一株欖李開花,群體全部進入花期。8 月14 日,少于10%植株開花,群體進入終花期,個別欖李植株處于零星開花階段,主要表現為新花苞繼續在花序上爆出,或自葉腋內爆出,開花量較少,8 月31 日所有欖李植株開花結束。綜上所述,欖李群體開花時長約為106 d。
欖李花序為腋生總狀花序,偶有花序軸著生花序,或花苞直接從葉腋爆出,花序軸上單花互生排列,成熟花序軸上小花2~12 朵不等,花序上開花順序為自基部由下而上開放,持續時間5~18 d不等,平均天數為(10.80±3.58) d,果實成熟后花序軸變紅褐色留存于枝條。
2.1.2 單花開花動態
欖李的單花期為7~13 d,平均(9.1±1.3) d,欖李的單花花期的開放可分為4個時期,分別經歷萌動期、初展期、盛開期、掉落期。①萌動期(圖1:A~B):花萼緊包花蕾,后花蕾逐漸膨大,花瓣伸長,露白。②初展期(圖1:C~D):花瓣分離逐漸張開,花藥白色尚未開裂,花絲卷曲尚未伸展,花藥位于柱頭下方,且距離花柱較近,柱頭有少量黏液分泌;從萌動期到初展期需3~7 d,平均(4.1±1.1) d。③盛開期(圖1:E~F):盛開期需要2 d 左右。開花當天花瓣完全張開,花絲伸展,花藥呈五五兩輪排列,一般于06:00~09:00 點開放,此階段花藥白色尚未開裂,柱頭花藥之間總體幾乎平齊,隨著溫度升高,花上露水蒸發,花藥迅速開裂。在天氣晴朗無雨水時,4~5 h 內花藥從內輪到外輪依次開裂,此時花粉爆出,呈淡黃色,開始散粉,到傍晚時花藥從淡黃色變為黃色,此時柱頭高度與外輪花藥幾乎平齊,內輪花藥低于柱頭。初展期到花盛開大概需要0.5 或1.0 d,溫度較低時需要經歷1 d 時間,溫度高時當天即可進入盛花狀態。開花第2天,花藥變為淡褐色,肉眼可見花藥攜帶花粉量減少,帶有少許花粉,花粉呈現黃褐色或黃色,此時柱頭與花藥位置總體仍保持平齊且柱頭有黏液分泌。④掉落期(圖1:G~I):從花絲脫落到花瓣脫落大約需要2~3 d,一般于開花第3 天,花絲開始松動,此時柱頭高度高于花藥,柱頭有粘液分泌,花藥從淡褐色變為褐色。開花第4~5 天,花瓣開始脫落,花絲脫落完成,花瓣開始脫落到全部脫落約1~2 d,柱頭宿存,不再有黏液分泌,單花花期結束。⑤果實成熟期(圖1J):從花朵掉落期算起,果實成熟時間平均需(31.7±2.4) d。子房膨大后,果實顏色變為黃色,視為果實成熟。

圖1 欖李單花開花動態A~B.萌動期;C~D.初展期;E~F.盛開期;G~I.掉落期;J.果實。Fig.1 Flowering dynamics of single flower of Lumnitzera racemosa A-B.Budding period;C-D.Initial period;E-F.Full blooming period;G-I.Drop period;J.Fruit.
2.1.3 花序及花部特征
由表1 可知,花序長度為34.18~70.34 mm,平均長度(53.18±9.42) mm。每花序單花平均(6.40±2.02)枚。花總長(15.37±0.94) mm。花萼綠色杯狀合生,基部窄上部寬,上端常有5個三角形萼齒,被絨毛,偶有11 萼齒,具小苞片2 枚,鱗片狀三角形,宿存。花冠直徑為(13.82±1.33) mm,花瓣白色與 萼 齒 互 生,花 瓣 長 為(6.66±0.63) mm,寬 為(2.64±0.24) mm。每花平均花瓣數目為(5.03±0.30),常見4 瓣、5 瓣、6 瓣,偶有8 瓣;雄蕊8~17個,常見10個。雄蕊呈內外兩輪均勻排列,內輪雄蕊高度矮于外側雄蕊高度,花絲長(5.74±0.44)mm,花柱長(7.50±0.69) mm,插生于萼筒中央,彎曲或者波浪狀,個數1,偶有2 個花柱現象。子房下位,常見1室,偶有2室,子房長(5.92±0.58) mm,內含胚珠3~5枚,平均(4.00±0.68)枚,常見4枚,倒掛懸生子房內,長橢圓球狀淡黃色,珠柄合生不等長,白色海綿絮狀。欖李果實為核果,胚珠多數,最終只有1 枚胚珠發育為種子,存在少數多胚現象。
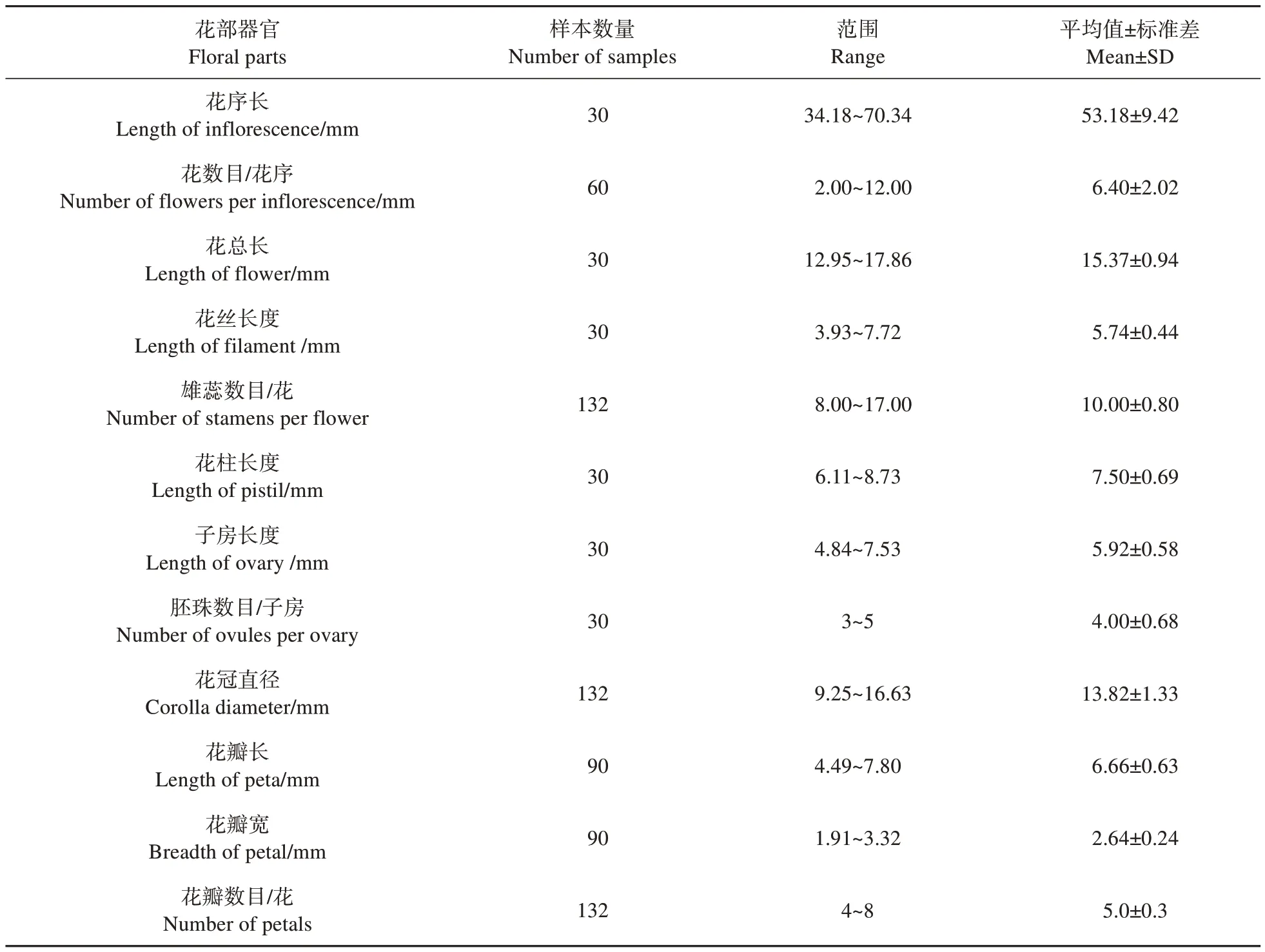
表1 欖李花部器官數量特征Table 1 Statistics of floral organs of Lumnitzera racemosa
2.2 柱頭可授性及花粉活力
2.2.1 柱頭可授性
由圖2 所示,柱頭在開花2 d 前可授性為0。開放前1 天(初展期:圖1C)可授性為20.00%。開放當天上午(盛開期:圖1E)柱頭可授性為30.00%。開花當天下午柱頭可授性為55.67%,柱頭可授性明顯升高,第3 天柱頭可授性達到100.00%,后可授性急速下降,在第5 天之后基本不具可授性。
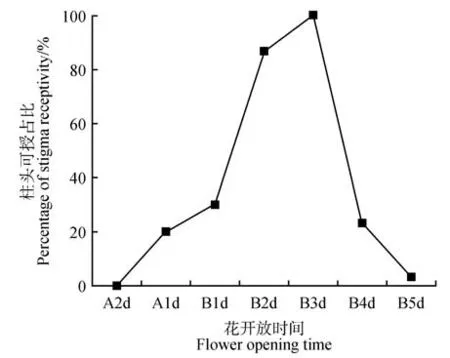
圖2 開放至不同天數柱頭可授情況Fig.2 Stigma receptivity under flower opening different days
開花前2 天柱頭不產生氣泡(圖3A),不具備可授性,肉眼可見花柱為淺綠色。開花當天上午部分柱頭具有可授性,產生氣泡數量很少且速度慢(圖3B),可授性較弱。第2 天柱頭可授性增強,產生氣泡增多(圖3C)。第3 天可授性達到最強,產生氣泡數量多且快速(圖3D),此時柱頭顏色變為淡褐色,花柱顏色為白色。第4天部分柱頭具有可授性,有氣泡產生,但大部分柱頭變干且顏色為褐色到深褐色不等(圖3E),基本不具可授性。
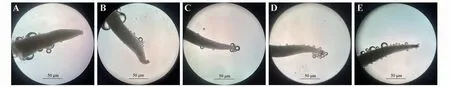
圖3 柱頭可授性變化A.開花前2天;B.開花當天;C.開花第2天;D.開花第3天;E.開花第4天。Fig.3 Changes of stigma receptivity under flower opening different days A.2 days before flowering;B.The day of flowering;C.The second day of flowering;D.The third day of flowering;E.The Fourth day of flowering.
2.2.2 花粉離體萌發
如圖4所示,不同蔗糖、硼酸質量濃度多重比較結果表明,在同一蔗糖質量濃度下,50.00 g·L-1蔗糖+0.20 g·L-1硼酸處理下花粉活力最高(圖5),為58.73%,顯著高于50.00 g·L-1蔗糖+0.3 g·L-1硼酸處理,與5.00 g·L-1、50.00 g·L-1蔗糖+0.2 g·L-1硼酸處理結果不顯著。5.00 g·L-1、3.00 g·L-1蔗糖處理下,花粉活力隨硼酸質量濃度升高而降低,其余為隨硼酸質量濃度升高呈現花粉活力先升高后降低趨勢。在同一硼酸質量濃度下,0.20 g·L-1硼酸+5.00 g·L-1蔗糖處理為最佳處理,顯著高于0.20 g·L-1硼酸下,5.00 g·L-1蔗糖,3.00 g·L-1蔗糖,100.00 g·L-1蔗糖處理,其中與10.00 g·L-1蔗糖處理不顯著,是因為10.00 g·L-1蔗糖下處理試驗中有花粉活力相對較高的重復結果,標準差偏大。0.1 g·L-1硼酸+100.00 g·L-1蔗糖處理下花粉不萌發,說明相對高的蔗糖質量濃度并不利于欖李花粉萌發。

圖4 不同質量濃度蔗糖、硼酸組合處理對花粉萌發的影響不同大寫字母代表同一蔗糖質量濃度下,不同硼酸質量濃度下的花粉活力之間差異顯著(P<0.05);小寫字母代表同一硼酸質量濃度下,不同蔗糖質量濃度下的花粉活力之間差異顯著(P<0.05)。Fig.4 Effect of different combinations of sucrose and boricacid concentrations on pollen germination The graphs containing different capital letters represented significant difference(sP<0.05) between pollen viability at the same sucrose concentration and different boric acid concentrations;Lowercase letters represented significant difference(sP<0.05) between pollen viability at the same boric acid concentration and different sucrose concentrations.
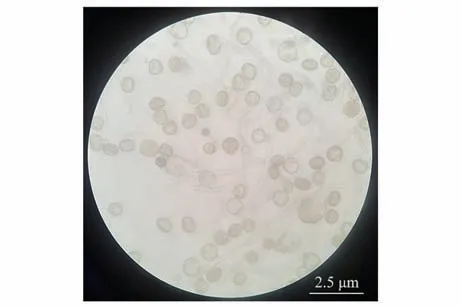
圖5 最佳處理下花粉萌發狀態(40倍物鏡)Fig.5 Pollen germination status under optimal medium(40× objective)
2.2.3 花粉活力測定結果
由圖6 可知,欖李在開花當天花粉活力最高,為58.73%,第2 天花粉活力顯著下降,為20.37%。第3天花粉不再具有活力,花絲開始松動脫落。
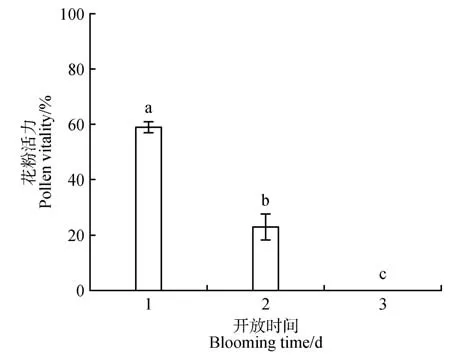
圖6 不同開放時間的花粉活力不同小寫字母代表不同開放時間的花粉活力之間差異顯著(P<0.05)。Fig.6 Pollen viability at different days The graphs containing different lowercase letters represent significant differences between pollen viability of different days.
2.3 繁育系統評判結果
2.3.1 單花花粉量及花粉-胚珠比
經統計,欖李單花平均花粉量為(19 856.8±3 190.6)粒,胚珠數目(3.6±0.6)粒,P/O為5 740.50±1 318.20。根據Cruden[19]標準,欖李的繁育系統為專性異交。
2.3.2 雜交指數(OCI)
欖李盛開時期花朵直徑為(13.82±1.33) mm(n=132),大于6 mm,OCI值記為3;花藥開裂后,柱頭具有較弱可授性,雌蕊尚未完全成熟,此時花粉已具有最高活力,由此推斷,欖李花朵為雄蕊先熟,OCI 值記為1;柱頭與花藥高度平齊,OCI 值計為0;欖李雜交指數(OCI)為4,參照Dafni[17]標準,欖李繁育系統為部分自交親和,異交,需要傳粉者。
2.3.3 授粉試驗結果
欖李果實成熟后,每個果實只有1 粒種子產生,為單種子果實。由表2 可知,無融合生殖試驗結實率較高,為95.00%,敗育率為100%,無種子產生,所以欖李在結實過程中基本不存在無融合生殖現象。自然授粉(對照組)、人工自花授粉、同株異花、異株異花授粉結實率分別為91.67%、78.33%、72.50%、76.67%,敗育率分別為35.00%、85.83%、46.67%、64.17%,各處理下均有種子產生,說明欖李自交和異交均是親和的。欖李果實自然對照下處理敗育率顯著低于人工自花授粉、人工異株異花授粉,低于人工同株異花敗育率(46.67%),但差異不顯著,說明可能在自然狀態下欖李花朵傳粉同株異花傳粉機會更多,其次為異株異花傳粉。授粉試驗結果表明,欖李人工自花授粉和同株異花授粉敗育率的平均值接近于異株異花授粉,則表明欖李繁育系統為自交和異交同時存在的混合交配系統。
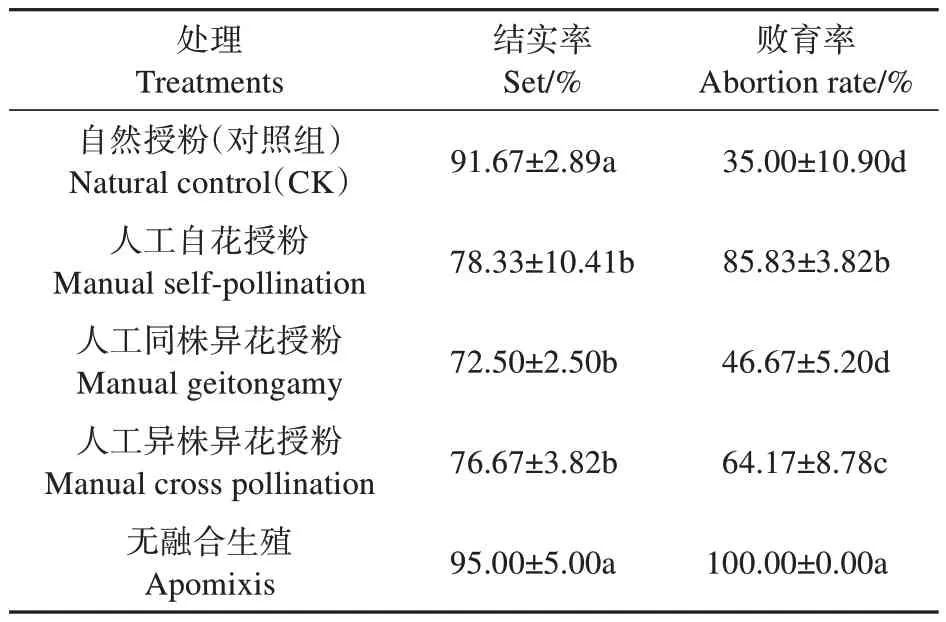
表2 授粉試驗結果Table 2 Pollination test results
通過雜交指數(OCI)、花藥-胚珠比(P/O)、以及授粉試驗測定結果,綜合判定欖李繁育系統為異交為主,部分自交親和,需要傳粉者。
3 討論
欖李存在林下更新慢、更新困難等現象,并因此造成其逐漸瀕危的現狀。開展欖李開花生物學及繁育系統研究,為今后揭示其瀕危機制以及開展相關保護行動,具有重要意義。
位于不同地理位置欖李開花時間以及開花時長不同。位于澳大利亞昆士蘭州的欖李開花時間為10 月到次年3 月[20],為期約6 個月;位于印度安得拉邦的欖李開花時間為7 月中旬至8 月中旬[21],為期約1 個月;本研究中,位于珠海淇澳所引種的欖李花期為5 月中旬至8 月末,約3 個月。 3 個地區欖李開花時間以及時長均不同,位于印度安得拉邦的欖李群體開花時間,開花時間較短,而位于澳大利亞昆士蘭州、廣東珠海淇澳開花時間相對較長,同一樹種在不同的地區,往往表現出不同開花物候來適應環境,從而保障繁殖成功。
本研究中,異花授粉的敗育率顯著低于自花授粉的敗育率,且異株異花授粉的種子更加飽滿,欖李更適合異花授粉。根據柱頭可授性與花粉活力檢測結果,花朵開放第1 天花粉活力最強,為58.73%,開放至第3 天柱頭可授性最強,達到100.00%,為提高欖李授粉成功率,可以采集開花當天的花粉授粉到開花第3天的花朵上,從而提高欖李結實率以及種子質量和產量。
參照Cruden[19]標準,根據花粉-胚珠比(P/O),欖李的繁育系統為專性異交,但根據Dafni[17]標準,雜交指數(OCI)為4 時其繁育系統為異交為主,部分自交親和,需要傳粉者。授粉試驗也表明欖李繁育系統為自交和異交兼有的混合交配系統,所以根據P/O得出其繁育系統為專性異交的結果并不準確,這在植物長柱紫莖[22](Stewartia rostrata)、夏臘梅[23](Sinocalycanthus chinensis)的研究中也得到類似矛盾的結果。P/O 是雄性資源分配量的一個指標[24],本研究中P/O為5 740.50±1 318.20,花粉量較大,利于傳粉者將更多的花粉攜帶到其他花朵上,增加其遠交的機會,可以證實其繁育系統體現更多的是異交。所以綜合來看,欖李繁育系統為異交為主,部分自交親和,需要傳粉者。另外,人工授粉試驗表明欖李繁育系統為自交和異交同時存在的混合交配系統,自交及異交敗育率接近,而不是體現為異交敗育率低,導致這種結果的可能原因是欖李群體花期為5 月中旬,正值雨季,可能導致授粉效果有偏差。
無論從花部結構來看,還是從花壽命角度來看,欖李繁育系統更趨向于異交,不利于自交。欖李花藥呈現內外2圈(每圈各4或5個花藥)的排列方式,內圈花藥低于外圈花藥,所在位置高度低于柱頭,外圈花藥與柱頭位置高度平齊,但距離柱頭位置相對較遠并不利于自花授粉,但有利于傳粉者將花粉攜帶到其他花朵之上,則更有利于異花授粉。Tomlinson 等[20]研究認為,欖李為明顯的雄蕊先熟,單個花朵并不能自花授粉。Raju 等[21]研究結果表明,欖李人工自花授粉結實率為35.00%,非人工自花授粉結實率為21.00%,欖李具備自花授粉能力。本研究中,人工自花授粉結實率為78.33%,敗育率為85.83%,存在自花授粉現象,也得到欖李具有自花授粉能力的結果,這與Tomlinson 等[20]認為欖李不具備自花授粉結論相反,與Raju 等[21]所得結果相一致。經柱頭可授性檢測,柱頭在開花后的第1 天至第3 天均有可授性,但第1、2 天的柱頭可授性小于第3 天,這明顯不利于自花授粉,更趨于異花授粉,人工授粉試驗也表明欖李異花授粉成功率高于自花授粉成功率。
欖李的傳粉機制更傾向于異交。根據相關研究結果,大多數植物的繁育系統表現為自交與異交同時存在的混合交配模式[25-28],這種交配模式可使得植物在面臨差的環境條件保證繁殖成功,欖李的異交繁育系統,也正是符合該模式的一種體現。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
