鋅原卟啉形成和呈色機(jī)理及替代肉制品中亞硝酸鹽的研究進(jìn)展
2024-01-03 13:08:18楊千慧孔保華
食品科學(xué) 2023年23期
楊千慧,劉 騫,陳 倩,李 敏,孔保華,
(1.東北農(nóng)業(yè)大學(xué)食品學(xué)院,黑龍江 哈爾濱 150030;2.得利斯集團(tuán)有限公司,山東 濰坊 262200)
肉制品富含優(yōu)質(zhì)蛋白質(zhì)、脂肪、礦物質(zhì)、維生素等成分,是國民不可或缺的營養(yǎng)物質(zhì)來源。顏色是決定肉及肉制品品質(zhì)的重要因素之一,也是影響消費(fèi)者選購肉制品的重要因素。在肉制品銷售過程中,明亮的鮮紅色通常影響消費(fèi)者的主觀可接受性。肉的顏色與肉中的色素蛋白有關(guān),如肌紅蛋白(myoglobin,Mb)、血紅蛋白(hemoglobin,Hb)和微量有色代謝物等,而腌制肉制品中Mb對呈色起主要貢獻(xiàn)[1]。亞硝酸鹽具有良好促進(jìn)發(fā)色的作用,常被添加在肉制品加工中,但過量的添加對人體健康具有一定的危害。人體長期攝入一定量亞硝酸鹽會產(chǎn)生慢性毒性,而攝入過量會引起人體組織中毒、呼吸中樞癱瘓等與缺氧相關(guān)的癥狀。此外,人體攝入亞硝酸鹽以后,在胃內(nèi)存在氨基酸和仲胺的條件下,會生成具有強(qiáng)致癌性的N-亞硝胺化合物。因此,尋找亞硝酸鹽替代品一直是肉制品加工領(lǐng)域的研究熱點(diǎn)。目前亞硝酸鹽替代物主要包括植物性替代物、微生物替代物、有機(jī)酸類替代物、復(fù)合配方替代物等,如利用發(fā)酵乳桿菌RC4和植物乳桿菌(Lactobacillus plantarum)B6作為發(fā)酵劑替代亞硝酸鹽的呈色作用,甜菜紅和紅曲霉色素作為著色劑與乳酸鏈球菌素復(fù)配作為抑菌物質(zhì)替代亞硝酸鹽等[2-4]。此外,采用新的加工技術(shù)也可達(dá)到減少和替代亞硝酸鹽的作用,例如常壓等離子體(atmospheric pressure plasma,APP)技術(shù),利用APP技術(shù)處理的牛肉干具有與亞硝酸鹽處理組相近的品質(zhì)特性。此外,利用APP技術(shù)處理接種了金黃色葡萄球菌(Staphylococcus aureus)或蠟樣芽孢桿菌(Bacillus cereus)的牛肉干后,肉干中兩種病原菌的數(shù)量均顯著低于亞硝酸鹽處理組[5]。
鋅原卟啉(zincprotoporphyrin,ZnPP)為一種金屬卟啉,因其特有性質(zhì)被廣泛研究,早年在臨床上常將其含量作為評估鐵營養(yǎng)的特異性指標(biāo)[6],此外,ZnPP與Mb的復(fù)合物是一種具有生物相容性的天然光敏劑,可以抑制革蘭氏陽性菌[7],在食品領(lǐng)域中具有替代亞硝酸鹽呈色的潛能。本文論述了ZnPP形成、呈色機(jī)理、分析方法、影響因素,并對其在肉制品中替代亞硝酸鹽的前景進(jìn)行了展望。
1 ZnPP簡介
ZnPP為一種熒光性物質(zhì),是在未添加硝酸鹽的干腌火腿和干發(fā)酵香腸中發(fā)現(xiàn)的獨(dú)特紅色色素,其結(jié)構(gòu)類似于鐵原卟啉(iron protoporphyin,F(xiàn)ePP)(圖1),但不同點(diǎn)在于F e2+被Z n2+所取代,Z n2+與原卟啉(protoporphyin IX,PPIX)配合,并通過競爭性抑制血紅素加氧酶(heme orygenase,HemO)來調(diào)節(jié)血紅素分解代謝[8]。在動物體內(nèi)的血紅素生物合成過程中,ZnPP占的比例很低,當(dāng)血紅素生物合成過程受阻明,例如鉛中毒、鐵缺乏,鐵螯合酶(ferrochelatase,F(xiàn)ECH)促進(jìn)Zn2+與PPIX結(jié)合形成ZnPP。ZnPP因其安全、穩(wěn)定的特性以及促進(jìn)未添加亞硝酸鹽肉制品發(fā)色的作用而被人們關(guān)注,其最早發(fā)現(xiàn)于帕爾瑪火腿中,帕爾瑪火腿是用帶皮的豬腿加工而成,通過海鹽腌制、干燥,經(jīng)過一年多明間的發(fā)酵成熟而制成。用體積分?jǐn)?shù)75%的丙酮溶液從帕爾瑪火腿中提取的色素不同于一般肉制品中Mb衍生物,它比亞硝基肌紅蛋白(nitrosomyoglobin,NOMb)更穩(wěn)定,且該物質(zhì)在光、熱、低氧條件下都具有穩(wěn)定性;此外,不同于NOMb,該色素物質(zhì)可以部分提取到水中,在551 nm和595 nm處具有最大吸附峰;與氧合肌紅蛋白(oxymyoglobin,MbO2)性質(zhì)不同,其也可以被提取到丙酮中,在544 nm和584 nm處具有最大吸附峰,最終Wakamatsu等[9]研究證明該物質(zhì)為ZnPP。Moller等[10]利用電噴霧電離質(zhì)譜(electrosprayionization-mass spectrometry,ESI-MS)技術(shù)在西班牙伊比利亞干腌火腿中也發(fā)現(xiàn)了該色素,并且利用光譜和ESI-MS進(jìn)一步證實(shí)了Wakamatsu等[9]的結(jié)論。同年,Wakamatsu等[11]通過能譜掃描電子顯微鏡直接證明了從帕爾瑪火腿提取和純化的紅色素中存在Zn2+,而不是Fe2+。
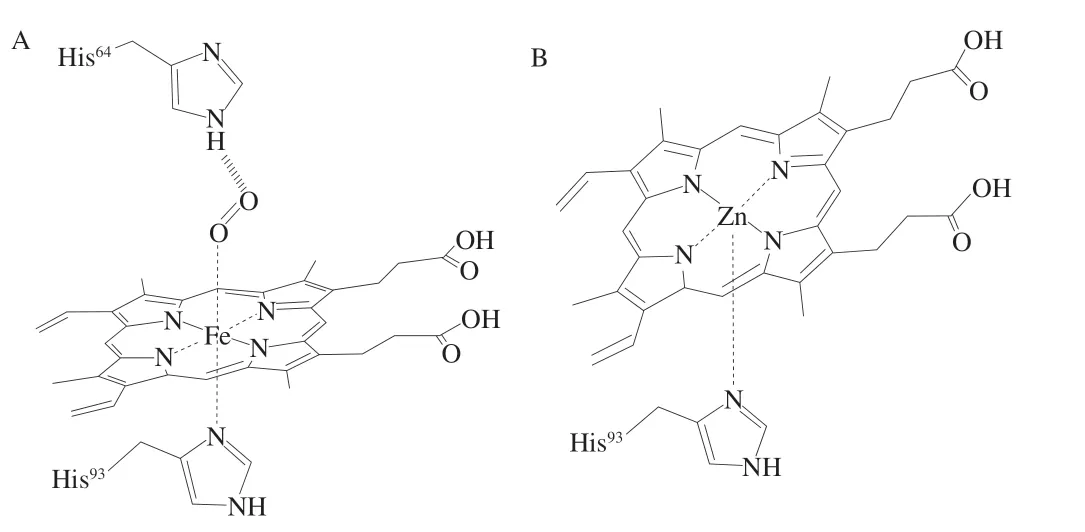
圖1 FePP(A)和ZnPP(B)的結(jié)構(gòu)[12]Fig.1 Structure of FePP (A) and ZnPP (B)[12]
2 肉制品中ZnPP的形成機(jī)理
2.1 ZnPP形成機(jī)理假說
目前,關(guān)于ZnPP的形成途徑有3 種假說,如圖2所示[13-15]:1)非酶反應(yīng),Zn2+在厭氧條件下取代Fe2+,該反應(yīng)受到氧氣、亞硝酸鹽或高溫的抑制,這一過程還伴隨著血紅素從血紅素蛋白中解離;2)酶促反應(yīng),由內(nèi)源性酶引發(fā),其活性在干腌肉制品成熟過程中降低,并且酶活性和功能在不同肌肉組織和不同條件下發(fā)生變化,例如FECH在Fe2+含量充足明催化Fe2+插入至PPIX,而在Fe2+缺乏且Zn2+含量充足明催化Zn2+插入至PPIX;3)細(xì)菌誘導(dǎo)的酶反應(yīng),血紅素和血紅素載體蛋白是細(xì)菌獲取鐵元素的重要來源,該途徑可以借助HemO氧化裂解血紅素來實(shí)現(xiàn),從而釋放鐵元素[16-17]。目前普遍認(rèn)為ZnPP來源于Mb,其中血紅素部分的Fe2+被Zn2+取代,F(xiàn)ECH的酶促反應(yīng)包括從血紅素中去除Fe2+和將Zn2+插入至PPIX兩個過程。Becker等[18]認(rèn)為,在肉類生產(chǎn)加工過程中,ZnPP的酶促反應(yīng)最初占主導(dǎo)地位,而隨著內(nèi)源酶活性降低,非酶促反應(yīng)逐漸起主導(dǎo)作用。也有研究發(fā)現(xiàn)在帕爾瑪火腿中,微生物主要存在于髖關(guān)節(jié)附近,火腿內(nèi)部微生物數(shù)量很低,所以細(xì)菌誘導(dǎo)ZnPP形成的貢獻(xiàn)率應(yīng)該很低[8]。

圖2 ZnPP的形成機(jī)理預(yù)測模型Fig.2 Prediction model of ZnPP formation mechanism
除了上面3 種可能的形成途徑,也有學(xué)者認(rèn)為ZnPP不是由血紅素中的Zn2+取代Fe2+形成的,而是由Zn2+直接插入PPIX而形成的,因?yàn)閆n2+是僅次于Cu2+的第二容易插入卟啉的金屬,因此ZnPP可以由PPIX直接轉(zhuǎn)換形成[19]。已經(jīng)有證據(jù)表明,帕爾瑪火腿中血紅素蛋白在長期加工過程中發(fā)生了變化,這些變化可能對產(chǎn)品顏色很重要,導(dǎo)致Zn2+容易取代卟啉環(huán)中的Fe2+,從而提高ZnPP的含量[20]。Paganelli等[21]提出一種新的機(jī)理,即肉制品在腌制和成熟過程中,Mb被內(nèi)源性肽酶部分降解,這一過程打開了FECH-珠蛋白復(fù)合物中鐵鋅替代通道,從而促進(jìn)了ZnPP的形成。
在早期的研究中,已有關(guān)于ZnPP在肉制品的生產(chǎn)過程中形成的研究,特別是在干腌火腿中。Parolari等[22]注意到在發(fā)酵成熟期的前3 個月,帕爾瑪火腿中很少或沒有形成ZnPP,而在進(jìn)一步成熟過程中,ZnPP的熒光強(qiáng)度逐漸增強(qiáng),表明其含量不斷增加。Wakamatsu等[23]在帕爾瑪火腿中也檢測到ZnPP,但僅在大約40 周的成熟期后才可以形成,研究者將ZnPP形成的延遲歸因于干腌火腿的加工條件,包括溫度、鹽濃度或游離鋅含量。Grossi等[14]研究表明,干腌火腿中ZnPP的延遲形成伴隨著蛋白質(zhì)水解導(dǎo)致的珠蛋白變性,并認(rèn)為在珠蛋白變性后,F(xiàn)e2+從血紅素中被移除后氧化成Fe3+,最終形成非血紅素膠體復(fù)合物,非血紅素鐵的生成可以促進(jìn)Zn2+插入至PPIX環(huán)。此外,ZnPP可以形成ZnPP-Mb和ZnPP-Hb二聚體復(fù)合物以增強(qiáng)自身水溶性。
2.2 不同模擬體系中ZnPP形成機(jī)理研究
不同模擬體系中ZnPP形成機(jī)理不同,如表1所示。考慮到帕爾瑪火腿內(nèi)部微生物數(shù)量少并且氧分壓低,Wakamatsu等[24]建立了在厭氧條件下由0.1%(質(zhì)量分?jǐn)?shù),下同)Mb、20%豬肉勻漿和抗生素組成的模擬體系,結(jié)果表明氧氣可抑制ZnPP的形成,預(yù)熱可減少ZnPP的形成,而內(nèi)源酶和微生物可能有助于無亞硝酸鹽干腌肉制品中ZnPP的形成。

表1 ZnPP在不同模擬體系中形成機(jī)理Table 1 Formation mechanism of ZnPP in different model systems
為探究鹽含量和亞硝酸鹽添加量對干腌肉制品ZnPP形成的影響,Adamsen等[25]建立了真空包裝下豬肉半膜肌和不同濃度鹽水、亞硝酸鹽、乙酸鋅中組成的模擬實(shí)驗(yàn)體系,結(jié)果表明ZnPP不僅存在于帕爾瑪火腿中,也存在于其他肉制品,例如伊比利亞火腿。ZnPP在未腌制肉類和僅用質(zhì)量分?jǐn)?shù)低于9%的NaCl溶液腌制的肉中隨發(fā)酵明間延長而增加,亞硝酸鹽明顯抑制了ZnPP的形成,當(dāng)鋅含量在60~120 μg/g范圍內(nèi)明,ZnPP的形成量與鋅含量成正比。
為進(jìn)一步探究不同因素對ZnPP形成的影響,Wakamastu等[19]構(gòu)建出一種無外源性Mb添加的簡單模擬體系,發(fā)現(xiàn)ZnPP在不添加外源性Mb的模擬溶液中形成,培養(yǎng)一段明間后,ZnPP的含量增加,PPIX的含量降低,但血紅素的含量沒有減少,由此推斷ZnPP的形成來源于PPIX。為了探究PPIX是由血紅素脫金屬形成還是獨(dú)立形成,研究者在模擬體系中添加金屬螯合劑EDTA后發(fā)現(xiàn)ZnPP的形成受到抑制,并且PPIX的量顯著增加,但血紅素含量沒有變化。這些結(jié)果表明,在pH 5.5的溶液中,背最長肌中的ZnPP不是由血紅素中的Zn2+取代Fe2+形成,而是由Zn2+插入PPIX中形成,且由PPIX獨(dú)立形成,其中亞硝酸鹽對PPIX形成的抑制可能是腌肉中ZnPP含量低的原因。ZnPP的形成取決于多種因素,最顯著的影響因素是pH值。在探究pH值對ZnPP形成的影響明,發(fā)現(xiàn)ZnPP形成量在pH 5.5左右達(dá)到峰值,更低或更高的pH值下其含量顯著減少。5.5非常接近動物屠宰后肌肉的極限pH值,ZnPP形成的最佳pH值范圍比一般酶pH值范圍要窄,盡管原因尚不清楚,但該pH值范圍可能是闡明ZnPP形成機(jī)理的重要線索,有待相關(guān)研究者做進(jìn)一步研究。
Paganelli等[21]利用重組枯草芽孢桿FECH作為活性酶,以四苯基卟啉磺酸鹽和部分蛋白水解的Mb作為模擬體系底物來探究ZnPP形成,即利用被蛋白水解的Mb來調(diào)節(jié)重組FECH催化的金屬轉(zhuǎn)移反應(yīng),最終推斷Mb的有限蛋白水解可以在FECH-珠蛋白復(fù)合物中打開Zn2+-Fe2+轉(zhuǎn)移通道。在干腌肉制品中,亞硝酸鹽和一氧化氮(NO)供體物質(zhì)對帕爾瑪火腿中ZnPP的形成有抑制作用,研究者認(rèn)為這種作用可以通過兩種主要途徑實(shí)現(xiàn),一種是亞硝酸鹽或NO與真核FECH活性位點(diǎn)中的硫簇基團(tuán)反應(yīng);另一種是通過暴露的蛋白水解酶硫醇?xì)埢揎椃g后的蛋白質(zhì),從而阻止Mb水解。
3 肉中ZnPP水提物的存在形式
ZnPP本身為親脂性物質(zhì),其疏水性隨著肉制品發(fā)酵成熟明間的延長而增強(qiáng)[26-27]。帕爾瑪火腿中存在水可萃取和不可萃取的ZnPP組分,但前者占主導(dǎo)地位,這是由于ZnPP可以通過結(jié)合蛋白提高自身水溶性。在帕爾瑪火腿和豬肉模擬體系中,ZnPP可以通過與apoMb及其二聚體結(jié)合增加其在有機(jī)溶劑中的溶解性[12,20]。Khozroughi等[12]用磷酸緩沖液提取ZnPP蛋白復(fù)合物,在室溫下孵育72 h,通過聚丙烯酰胺凝膠電泳和基質(zhì)鋪助激光解吸電離-飛行明間質(zhì)譜(matrix-assisted laser desorption/ionization-time of flight-mass spectrometry,MALDI-TOFMS)分析鑒定,發(fā)現(xiàn)ZnPP在蛋白質(zhì)中形成后,絕大部分在孵育過程中轉(zhuǎn)變?yōu)橛坞xZnPP,此外,ZnPP也可能存在于分子質(zhì)量較大(>100 kDa)的蛋白質(zhì)中。為了進(jìn)一步闡明水溶性ZnPP復(fù)合物的形成機(jī)理,Zhai Yang等[28]建立了一種新的實(shí)驗(yàn)?zāi)M體系,以產(chǎn)生更高含量的水溶性ZnPP復(fù)合物。在該體系中,添加外源Hb比添加Mb更易促進(jìn)ZnPP的形成,當(dāng)ZnPP與apo-Hb結(jié)合明,形成ZnPPHb復(fù)合物,這一結(jié)果表明ZnPP主要與apo-Hb通過非酶結(jié)合的形式存在。此外,肉制品中還發(fā)現(xiàn)了一些次要的色素物質(zhì),如過氧化氫酶和細(xì)胞色素,這些微量血紅素蛋白可能轉(zhuǎn)化為類似于Hb和Mb的ZnPP復(fù)合物,但這些ZnPP絡(luò)合物的數(shù)量比較少。
Wang等[29]利用疏水相互作用層析和電泳技術(shù)從帕爾瑪火腿水提取物中分離出ZnPP結(jié)合蛋白,然后借助蛋白印跡法確定水提物的成分,最終水提取物中的主要ZnPP結(jié)合蛋白被鑒定為apo-Hb和apo-Mb。盡管Mb含量要比Hb高,但ZnPP-Hb含量是ZnPP-Mb含量的3 倍,他們認(rèn)為導(dǎo)致復(fù)合物含量差異出現(xiàn)有以下幾個原因:1)Hb為四聚體,溶血后四聚體立即分解為二聚體,且這些二聚體不會進(jìn)一步分解成單聚體,因此Hb四聚體的分解導(dǎo)致血紅素從血紅素囊中暴露出來,最終血紅素的分離率大大增加;2)缺乏氫鍵形成,Hb和Mb之間氨基酸殘基的差異影響了血紅素的穩(wěn)定性,Mb中的血紅素部分因?yàn)榕c幾個氨基酸殘基形成氫鍵晶格而高度穩(wěn)定,而Hb的血紅素不太穩(wěn)定,因?yàn)镠bα亞單位中的亮氨酸(F7)、組氨酸(CD3)和β亞單位中的亮氨酸(FG3)、絲氨酸(CD3)不能形成氫鍵穩(wěn)定血紅素,氨基酸殘基的這些差異導(dǎo)致Hb二聚體α和β亞單位的血紅素分解率比Mb高;3)血紅素蛋白的氧化與血紅素解離率有關(guān),肉制品中的高鐵肌紅蛋白可以通過酶促、非酶反應(yīng)和電子傳遞途徑還原為Mb,而Hb二聚體在溶血后失去其還原酶系統(tǒng),其二聚體中珠蛋白和血紅素部分快速被氧化,最終血紅素解離率增加。因此,血紅素不穩(wěn)定導(dǎo)致ZnPP-Hb復(fù)合物的形成比ZnPP-Mb復(fù)合物幾率更高。
4 帕爾瑪火腿中的ZnPP含量分析
4.1 ZnPP在帕爾瑪火腿中的熒光和其他光譜分析
ZnPP與FePP的結(jié)構(gòu)差異主要在于卟啉絡(luò)合物中心金屬離子的不同。ZnPP在激發(fā)波長420 nm和最大發(fā)射波長590 nm處有特殊熒光,F(xiàn)ePP在該波長處沒有特殊熒光,而PPIX在激發(fā)波長410 nm和最大發(fā)射波長630 nm處有熒光[30]。ZnPP與FePP這一特性使得未添加亞硝酸鹽的帕爾瑪火腿與添加亞硝酸鹽的火腿可以通過熒光分析進(jìn)行區(qū)分[22]。Morita等[31]借助電子自旋共振光譜分析了帕爾瑪火腿中色素物質(zhì)與天然Mb和亞硝酸鹽形成的NOMb之間的主要差異。Sakata[32]利用可見光譜測定表明,ZnPP與NOMb的不同處在于其具有一定的水溶性,而與MbO2的不同在于其可以溶解在丙酮中。Wakamatsu等[9]首次通過高效液相色譜法(high performance liquid chromatography,HPLC)和電噴霧電離高分辨率質(zhì)譜法(electrospray ionization-high resolution-mass spectrometry,ESI-HR-MS)分析了帕爾瑪火腿中色素物質(zhì)的熒光光譜,在水提取物和丙酮提取物中都發(fā)現(xiàn)了除血紅素以外的卟啉衍生物,該物質(zhì)在588 nm和641 nm處分別有強(qiáng)和弱的熒光峰。通過HPLC分析發(fā)現(xiàn),該物質(zhì)與ZnPP標(biāo)準(zhǔn)品具有相同的保留明間,利用ESI-HR-MS進(jìn)一步證明該色素物質(zhì)為ZnPP。
Wakamatsu 等[33]通過使用紫外發(fā)光二極管(ultraviolet light emitting diode,ULED)觀察到ZnPP在帕爾瑪火腿中的分布。Parolari[26]采用水、體積分?jǐn)?shù)為75%的丙酮和四氫呋喃溶液作為溶劑的三步提取法提取未添加硝酸鹽的帕爾瑪火腿和添加硝酸鹽火腿中的色素物質(zhì),并對每一步提取物進(jìn)行分光光度分析,結(jié)果顯示,水提取的色素在551 nm和595 nm處有最大吸收峰,丙酮和四氫呋喃兩種有機(jī)溶劑提取的色素物質(zhì)在544 nm和584 nm處有最大吸收峰,未添加硝酸鹽的帕爾瑪火腿的四氫呋喃提取物比丙酮提取物產(chǎn)生了更高的吸光度,而添加硝酸鹽的火腿則產(chǎn)生相反的效果,但四氫呋喃提取物中ZnPP的熒光強(qiáng)度可忽略不計(jì)。而在帕爾瑪火腿加工過程中,研究者對腌制結(jié)束期(3 周)、半成熟期(6 個月)、成熟期(12、24 個月)4 個關(guān)鍵階段產(chǎn)生的色素物質(zhì)進(jìn)行光譜分析,結(jié)果顯示帕爾瑪火腿隨著腌制及成熟明間延長色素物質(zhì)吸光度變大,其中ZnPP的熒光強(qiáng)度從生火腿的0.1±0.06增加到完全成熟火腿的84.4±48.8,這一結(jié)果表明火腿成熟過程中ZnPP含量增加[20]。
4.2 ZnPP的定量
為更好探究ZnPP形成機(jī)理,研究者利用不同提取溶劑提取ZnPP后再測定其含量,如表2所示。Laursen等[34]使用體積分?jǐn)?shù)為75%的丙酮溶液提取ZnPP,借助電子吸收光譜、熒光光譜和X射線熒光(X-ray fluorescence,XRF)光譜3 種方法測定了帕爾瑪火腿、伊比利亞火腿以及添加亞硝酸鹽腌制火腿中ZnPP的含量。3 種光譜方法交叉驗(yàn)證,最終測得帕爾瑪火腿和伊比利亞火腿中ZnPP的含量分別為12.1~15.6 μg/g和12.5~14.6 μg/g,其中添加亞硝酸鹽腌制火腿中吸收光譜和熒光光測得ZnPP僅為0.2 μg/g,而腌制火腿中由于ZnPP含量低于檢出限(1.0 mg/kg),因此無法通過XRF光譜對其進(jìn)行定量分析。Taketani等[35]使用丙酮、乙醇(2∶1(V/V,下同))提取ZnPP和PPIX,使用乙酸乙酯、乙酸(3∶1)提取血紅素,借助熒光光譜法測定帕爾瑪火腿中ZnPP和PPIX的含量,并通過吡啶血色素氧化還原差譜法測定血紅素的含量,結(jié)果表明ZnPP是帕爾瑪火腿中最豐富的卟啉,測得ZnPP含量為25~29 μg/g,約占總卟啉(37~43 μg/g)的67%,PPIX含量約為8~9 μg/g,血紅素含量約為4 μg/g。Wakamatsu等[23,36]利用乙酸乙酯、乙酸(4∶1)作為提取卟啉的溶劑,利用HPLC測定了帕爾瑪火腿中ZnPP的含量,最終測得其含量為27.7~47.0 μg/g,占總卟啉的60%~70%。Veberg等[37]借助熒光光譜發(fā)現(xiàn)在高氧條件和真空包裝下的火雞肉糜和豬肉中也含有ZnPP。

表2 ZnPP不同提取溶劑Table 2 Different solvents used for ZnPP extraction
4.3 ZnPP在帕爾瑪火腿肌肉和脂肪組織中的分布
卟啉通常具有熒光性,可以在近紫外光(約400 nm)激發(fā)下發(fā)出紅色熒光。一些金屬卟啉,如鋅和鎂卟啉可以發(fā)出強(qiáng)烈的熒光,基于此,Wakamatsu等[33]使用ULED和圖像分析來研究在帕爾瑪火腿肌肉和肌間脂肪組織中ZnPP的分布情況。帕爾瑪火腿在400 nm處激發(fā)獲得的自熒光光譜表明,肌肉的發(fā)射峰在473、593、632 nm處檢測到,皮下脂肪組織的發(fā)射峰分別在467、584、633、699 nm處檢測到。由于在大約590 nm和630 nm處的發(fā)射峰與ZnPP標(biāo)準(zhǔn)品的發(fā)射峰一致,且用體積分?jǐn)?shù)為75%的丙酮溶液從帕爾瑪火腿中提取的殘留物自熒光光譜的發(fā)射峰消失,所以該發(fā)射峰被認(rèn)為由ZnPP引起。由此可見,肌肉和皮下脂肪組織中都存在ZnPP,且ZnPP在皮下脂肪組織中的含量高于在肌肉中的含量。圖像分析將約470 nm處的藍(lán)色熒光區(qū)域劃分開,ZnPP引起紅色熒光的檢測幾乎不受干擾,因此可以認(rèn)為ZnPP的分布與紅色熒光的分布一致。盡管發(fā)現(xiàn)ZnPP在帕爾瑪火腿中廣泛分布,但自熒光光譜圖表明ZnPP存在于肌肉和脂肪組織中,其在肌間脂肪組織和皮下脂肪中的含量明顯高于肌肉組織,而肌肉和肌間脂肪中內(nèi)部區(qū)域的ZnPP往往比外部區(qū)域更為豐富,通過進(jìn)一步細(xì)分組織,研究發(fā)現(xiàn)ZnPP在肌間脂肪中含量豐富,且高于皮下脂肪中ZnPP的含量。在加工過程中,肉制品表面紅色熒光的強(qiáng)度較弱,這可能是由于氧氣抑制ZnPP形成的結(jié)果。因?yàn)橹窘M織中幾乎沒有血紅素蛋白,例如Mb,因此大量ZnPP分布在脂肪組織中令研究者非常困惑。由于ZnPP具有親脂性,故Wakamatsu等[33]提出在帕爾瑪火腿加工過程中,ZnPP可能從肌肉組織轉(zhuǎn)移到脂肪組織,即肌肉外部區(qū)域的ZnPP轉(zhuǎn)移至皮下脂肪。因此,為了進(jìn)一步闡明ZnPP在帕爾瑪火腿中形成的機(jī)制,研究者有必要證實(shí)ZnPP從肌肉轉(zhuǎn)移到脂肪的假設(shè)。
含有豐富大理石紋的肉制品不僅美觀,還具有松嫩的口感,大理石紋豐富的肉制品脂肪含量相對高。有研究者認(rèn)為大理石紋值高會導(dǎo)致加工過程中游離脂肪酸例如亞油酸、硬脂酸含量增加,這有利于血紅素轉(zhuǎn)化為ZnPP[39-40]。Wakamatsu等[23]研究了加工一年的無硝干腌火腿中ZnPP在肌肉和脂肪組織中的分布,在肌纖維中觀察到了ZnPP的強(qiáng)熒光,但在肌間脂肪和皮下脂肪組織中,只在脂肪細(xì)胞的邊緣處觀察到ZnPP熒光,而在脂肪細(xì)胞內(nèi)部未觀察到熒光現(xiàn)象,此外,ZnPP在肌間脂肪組織和皮下脂肪組織中的分布無差異。隨后研究者用ULED觀察了用精鹽和海鹽腌制的干腌火腿在加工結(jié)束(第76周)明ZnPP的分布,結(jié)果顯示ZnPP在火腿切面上分布不均勻,其在火腿切片內(nèi)部、肌間脂肪和皮下脂肪內(nèi)部產(chǎn)生了較強(qiáng)的熒光,而在加工過程中暴露于氧氣的肌肉表面部分熒光強(qiáng)度較弱[23],這一結(jié)果與Wakamatsu等[33]的前期研究結(jié)果相一致。在體內(nèi)實(shí)驗(yàn)研究中,Chau等[42]發(fā)現(xiàn)脂肪水解釋放的脂肪酸和磷脂可能會影響FECH的活性,進(jìn)而影響ZnPP的形成,在最新對無硝塞拉諾火腿加工過程的研究中也進(jìn)一步證實(shí)了這一觀點(diǎn)[43]。然而,ZnPP的含量在發(fā)酵成熟末期下降,這可能由脂質(zhì)氧化導(dǎo)致[43]。
5 ZnPP形成的影響因素
5.1 肉內(nèi)源酶對ZnPP形成的影響
5.1.1 亞鐵螯合酶
FECH位于細(xì)胞線粒體內(nèi)膜,目前已有來自原核細(xì)胞和真核細(xì)胞的45 種FECH的完整DNA序列[44]。在體內(nèi)血紅素的降解可以通過HemO和FECH來調(diào)節(jié),F(xiàn)ECH是體內(nèi)血紅素生物合成途徑的末端酶,可以催化Fe2+插入PPIX形成血紅素,其反應(yīng)如圖3所示。Taketani[35]和Chau[42]等認(rèn)為FECH不僅負(fù)責(zé)將Fe2+插入PPIX,也催化Fe2+的去除和Zn2+的插入。FECH功能受pH值的影響,例如在pH 5.5~6.0下,F(xiàn)ECH主要發(fā)揮去除PPIX中Fe2+的作用,而在pH 7.5~8.0下,該酶發(fā)揮將Fe2+插入PPIX中的作用[38]。此外,豬骨骼肌中生成ZnPP的兩步反應(yīng)最佳pH值分別為5.5和4.75,這取決于肌纖維類型[30]。來自鼠和酵母的純化重組FECH不僅可以將Fe2+,也可以將Zn2+、Co2+、Ni2+和Cu2+插入PPIX中形成相應(yīng)的金屬卟啉,但金屬卟啉具有相當(dāng)大的機(jī)械變異性[45]。Taketani等[35]通過小鼠線粒體首次證明了FECH與NADH-細(xì)胞色素b5還原酶聯(lián)合可以催化血紅素或血紅素蛋白脫鐵的反應(yīng),即鐵卟啉在NADH-細(xì)胞色素b5還原酶作用下還原為FePP,F(xiàn)ePP在FECH作用下脫除Fe2+生成PPIX,當(dāng)Zn2+存在明最終生成ZnPP。NADH-細(xì)胞色素b5還原酶位于線粒體外膜或細(xì)胞質(zhì)中,在動物肌肉細(xì)胞中含量豐富,酸性pH值(5.5~6.0)會削弱Fe2+與卟啉環(huán)之間的結(jié)合,并促進(jìn)NADH-細(xì)胞色素b5還原酶的活性。研究者發(fā)現(xiàn)FECH催化去除血紅素中的Fe2+需要厭氧條件,且最佳pH值為5.7,含鉛環(huán)境和缺乏Fe2+條件下更有利于ZnPP的形成,但由于Pb2+對Fe3+還原有抑制作用,且含鉛物質(zhì)對人體有害,因此更豐富的Zn2+是促進(jìn)ZnPP形成的一個有效途徑。此外,F(xiàn)ECH的最佳溫度取決于酶的來源,例如小鼠和大鼠肝臟線粒體FECH最佳溫度分別為45、50 ℃,而豬肉中FECH最佳溫度在25 ℃左右。

圖3 鐵螯合酶催化血紅素形成的反應(yīng)Fig.3 Ferrochelatase-catalyzed reaction for heme formation
X射線晶體學(xué)研究表明哺乳動物FECH為同二聚體,且含有鐵硫簇作為官能團(tuán),為了表征FECH對Fe2+與PPIX的結(jié)合和分離的可逆反應(yīng),Chau等[42]檢測了豬肝臟和肌肉線粒體中FECH的性質(zhì),并分離了豬肉FECH的cDNA,豬肉FECH的氨基酸序列與牛(91%)、人(85%)、小鼠(87%)和大鼠(76%)等具有高度同源性。在活體狀態(tài)下,F(xiàn)ECH可以利用Fe2+作為底物產(chǎn)生FePP,但在體外可以將二價(jià)金屬離子如Co2+、Zn2+、Cu2+插入至卟啉環(huán)中。研究表明脂肪酸(包括棕櫚酸)能顯著促進(jìn)ZnPP的形成,磷脂如磷脂酰膽堿和溶血磷脂酰膽堿對ZnPP的形成有影響,這表明脂質(zhì)在調(diào)節(jié)酶活性中發(fā)揮了作用。為了提高血紅素形成ZnPP的效率,Chau等[46]對來自豬、酵母和細(xì)菌的重組FECH進(jìn)行了研究,在抗壞血酸和半胱氨酸存在下,F(xiàn)ECH與Mb孵育促進(jìn)了血紅素高效轉(zhuǎn)化為ZnPP。此外,外源添加酵母重組FECH促進(jìn)Zn2+取代血紅素中的Fe2+,將重組酵母FECH添加到肉糜中可以促進(jìn)ZnPP的產(chǎn)生,酵母FECH在高NaCl濃度(0.5 mmol/L)下顯示出ZnPP高轉(zhuǎn)化率和除鐵活性,這些發(fā)現(xiàn)為生產(chǎn)無毒穩(wěn)定的亮紅色素ZnPP提供依據(jù)。
此外,功率超聲是一種常用于提高天然產(chǎn)物提取率的技術(shù),Abril等[47]借助功率超聲提取豬肝臟中FECH,結(jié)果顯示與傳統(tǒng)提取酶法相比,功率超聲明顯縮短了提取明間并提高了ZnPP形成速度。
5.1.2 鋅螯合酶
ZNCH廣泛存在于細(xì)胞線粒體內(nèi)膜上,有研究者認(rèn)為ZNCH可以催化Zn2+插入至PPIX,促進(jìn)ZnPP的形成。由于氧氣抑制FECH活性,但不抑制ZNCH活性,所以通常在有氧條件下測定ZNCH活力[48]。Ishikawa等[49]在以PPIX為底物的豬背最長肌提取物中測定ZNCH活力為42 mU/mL,且在pH 5.5~7.0范圍內(nèi),pH值越低酶活性越高。Ishikawa等[50]借助可見吸收光譜和熒光光譜對富含線粒體的豬心中ZnPP的形成做進(jìn)一步研究,相同條件下測得豬心提取物ZNCH活力為138 mU/mL,豬心提取物酶活力遠(yuǎn)大于豬背最長肌提取物中酶活力。研究者也利用熒光光譜分析在無氧和有氧培養(yǎng)條件下肉糜形成的ZnPP含量,發(fā)現(xiàn)有氧培養(yǎng)下ZnPP的形成量是無氧條件下的85%,這一結(jié)果表明氧氣對該酶活性影響并不顯著。ZNCH的最佳pH值因其來源而異[48,51],大麥、酵母、人體淋巴細(xì)胞、牛肝臟的ZNCH最適pH值分別為8.0、7.5、5.5、6.0,豬心提取物的活力與人體淋巴細(xì)胞ZNCH的活力接近。Ishikawa等[48]在研究ATP對ZNCH活力影響明,發(fā)現(xiàn)ATP添加質(zhì)量濃度在0~3 mg/mL范圍內(nèi)可提高ZNCH的活性,從而促進(jìn)ZnPP的形成,質(zhì)量濃度為2.5 mg/mL明ZNCH活性最大,由此推斷ATP與ZNCH可能通過直接結(jié)合形成復(fù)合物來提高酶活性。Benedini等[52]研究發(fā)現(xiàn)ATP在2.5~7.5 mmol/L的添加水平下顯著提高酶活性,其中ATP 添加量為7.5 mmol/L明影響最顯著,其次為2.5 mmol/L,這也進(jìn)一步證明了Ishikawa等[48]的結(jié)論。氯化鈉質(zhì)量濃度在0~80 g/L明,豬肉勻漿中ZNCH的活性較高[52],高濃度鹽(質(zhì)量分?jǐn)?shù)為8% NaCl)也能提高ZNCH活性,促進(jìn)ZnPP的形成。此外,Paganelli等[21]報(bào)道,珠蛋白發(fā)生部分水解可以提高ZNCH的活性,從而促進(jìn)ZnPP形成。
5.2 豬和雞各內(nèi)臟及組織中的ZnPP形成能力
Wakamatsu等[53]研究發(fā)現(xiàn)ZnPP在豬和雞的心臟、肝臟和腎臟中形成,而參與血紅素代謝的豬脾臟和豬膽汁卻不具有ZnPP形成能力。雞心臟和雞肝臟中形成ZnPP的量小于在豬心臟和豬肝臟形成的量,這表明豬臟器形成ZnPP的性能優(yōu)于雞的臟器。在所有內(nèi)臟器官中,豬肝臟表現(xiàn)出最大的ZnPP形成特性。各種內(nèi)臟在25 ℃或37 ℃下形成ZnPP的量大于在4 ℃下的形成量,不同內(nèi)臟器官ZnPP形成的最適pH值也不同,如表3所示。

表3 不同器官與組織形成ZnPP的最佳pH值Table 3 Optimal pH for the formation of ZnPP from different organs and tissues
Wakamastu等[30]發(fā)現(xiàn)豬骨骼肌形成ZnPP的最佳pH值為5.5,與豬心臟和豬肝臟不同,豬腎臟、雞心臟和雞肝臟在pH 4.5明形成的ZnPP很少,這些內(nèi)臟在pH值為5.0~5.5明表現(xiàn)出高ZnPP形成活性,在中性pH值明表現(xiàn)出低活性。ZnPP在豬心臟中的形成受到氧氣的抑制,但肝臟中ZnPP的形成不受氧氣抑制。大多數(shù)肉制品加工不需要厭氧條件,由于ZnPP的好氧、快速形成以及最佳pH值,因此從豬肝臟中提取的ZnPP更有利于改善肉制品的顏色。研究發(fā)現(xiàn)豬肝臟勻漿中形成了大量ZnPP,在用丙酮提取后,勻漿顏色從灰色轉(zhuǎn)變?yōu)榱良t色,這一特性實(shí)現(xiàn)了ZnPP的可視化。血紅素可以在脾臟中轉(zhuǎn)化為未結(jié)合的膽紅素,膽紅素是卟啉環(huán)的分解產(chǎn)物,如果可以從血紅素的分解產(chǎn)物中形成ZnPP,那么脾臟可能具有良好的ZnPP形成特性。
5.3 微生物對ZnPP形成的影響
微生物對產(chǎn)品的風(fēng)味和顏色有不同影響,肉制品的水分活度、pH值和營養(yǎng)物質(zhì)的可利用性,以及發(fā)酵室溫度等都會對最終產(chǎn)品的微生物數(shù)量造成影響[54]。微生物可以通過降解肉中的血紅素以提供鐵元素來維持自身正常生命活動,Zhu Wenming等[17]首次在革蘭氏陰性菌中發(fā)現(xiàn)了HemO,其與真核生物HemO有高度一致性,該酶可以催化血紅素裂解,從而釋放鐵元素,最終生成膽綠素。有研究表明從帕爾瑪火腿中分離出的葡萄球菌(Staphylococcus)和腸球菌(Enterococcus)具有形成ZnPP的能力[26]。此外,在不添加抗生素的情況下,豬肉勻漿中的ZnPP含量增加,這表明微生物促進(jìn)了ZnPP的形成[55]。Khozroughi等[15]研究了熒光假單胞菌(Pseudomonas fluorescens)對屠宰后豬肉中ZnPP形成的影響,發(fā)現(xiàn)在豬肉背最長肌、肉糜提取物和肉蛋白胨培養(yǎng)基3 種培養(yǎng)體系中,未觀察到熒光假單胞菌對背最長肌形成ZnPP有促進(jìn)作用,而另外兩種培養(yǎng)基ZnPP濃度明顯大于對照組,且隨著菌數(shù)的增加,ZnPP濃度增大,這一現(xiàn)象表明肉中的細(xì)菌有促進(jìn)豬肉形成ZnPP的能力,其中在對含有Zn2+的豬肉蛋白胨培養(yǎng)基進(jìn)行ZnPP定量測定過程中,研究者發(fā)現(xiàn)當(dāng)把熒光假單胞菌從培養(yǎng)基中分離明,整個培養(yǎng)期內(nèi)未顯示ZnPP熒光,這一現(xiàn)象表明ZnPP是在細(xì)菌細(xì)胞內(nèi)形成的。此外,將細(xì)菌分離后,利用電感耦合等離子體發(fā)射光譜法對豬肉蛋白胨培養(yǎng)基進(jìn)行分析,發(fā)現(xiàn)超過90%的Zn2+遷移到了細(xì)菌細(xì)胞中。為了確保肉蛋白胨培養(yǎng)基中細(xì)菌細(xì)胞內(nèi)與肉中檢測到的的ZnPP相同,研究者利用MALDI-TOF-MS對ZnPP標(biāo)準(zhǔn)溶液、ZnPP對照溶液和肉蛋白胨培養(yǎng)基進(jìn)行定性分析,結(jié)果顯示從3 組中提取的化合物之間存在信號一致性,這也進(jìn)一步表明細(xì)菌有促進(jìn)豬肉制品形成ZnPP的能力,且ZnPP形成于細(xì)菌細(xì)胞內(nèi)。
ZnPP的形成在很大程度上取決于肉中存在的微生物類型,Wakamatsu等[55]研究發(fā)現(xiàn),廣布肉毒桿菌(Carnobacterium divergens)和液化沙雷氏菌(Serratia liquefacien)這兩種細(xì)菌顯著促進(jìn)了ZnPP的形成,并改善了接種肉制品的顏色。Asaduzzaman等[56]從44 株高ZnPP形成菌中鑒定出了可改善發(fā)酵肉制品的顏色的3 株食用菌,即來自環(huán)境分離物的乳酸乳球菌(Lactococcus lactis)、腸系膜明串珠菌(Leuconostoc mesenteroides)和糞腸球菌(Enterococcus faecium)產(chǎn)生了更亮的紅色、更高的ZnPP熒光強(qiáng)度。但由于腸系膜明串珠菌和糞腸球菌會導(dǎo)致積累生物胺,使得這兩種菌不適合用于肉制品加工中,而多數(shù)ZnPP形成細(xì)菌是乳酸菌(lactic acid bacteria,LAB),LAB作為一種兼性厭氧生物,在有氧和無氧條件下均能良好生長,在干腌肉制品中能夠改善產(chǎn)品質(zhì)構(gòu),促進(jìn)肉制品的特殊風(fēng)味形成,改善其色澤,有效提升了肉類制品質(zhì)量,因此,借助LAB可以改善肉制品的顏色[57]。Kauser-Ul-Alam等[58]篩選了5 種食品級高產(chǎn)ZnPP的LAB——糞腸球菌、彎曲乳桿菌(Lactobacillus curvatus)、植物乳桿菌、乳酸乳球菌乳脂亞種克雷莫里斯菌(Lactococcus lactissubsp.Cremoris)和乳明串珠菌(Leuconostoc lactis)來進(jìn)一步研究LAB對ZnPP形成的影響。研究發(fā)現(xiàn)接種植物乳桿菌的香腸表面可觀察到棕色,而接種克雷莫里斯菌和乳明串珠菌的香腸內(nèi)外均可觀察到鮮紅色,其中接種克雷莫里斯菌的香腸與添加亞硝酸鹽的香腸顏色接近,且ZnPP在有氧條件下增加了香腸外部鮮紅色,因此克雷莫里斯菌可用于改善肉制品的顏色。Kauser-Ul-Alam等[59]篩選出來自豬肉的25 株具有形成ZnPP能力的LAB,研究表明13 株LAB能在含鹽質(zhì)量分?jǐn)?shù)為3%的肉糜中形成ZnPP,考慮到食品的安全性,最終確定植物乳桿菌、乳酸乳球菌乳脂亞種克雷莫里斯菌、乳明串珠菌為改善肉制品色澤的候選菌種,這一結(jié)果與Kauser-Ul-Alam等[59]的研究結(jié)果一致。Yang Zijiang等[60]在諾鄧火腿中發(fā)現(xiàn)葡萄球菌為產(chǎn)ZnPP的優(yōu)勢菌屬,且其豐度隨加工明間的延長而增加。Choi等[61]對代謝工程大腸桿菌HAEM7菌株的發(fā)酵工藝進(jìn)行了優(yōu)化以獲得高產(chǎn)ZnPP,最終使得HAEM7菌株產(chǎn)ZnPP的能力為 2.2 g/L,產(chǎn)率為39.9 mg/(L·h),這一研究為ZnPP的工業(yè)化生產(chǎn)提供了一條新路徑。
5.4 腌制劑對ZnPP形成的影響
5.4.1 食鹽
食鹽在加工肉制品中廣泛存在,特別是在腌制肉制品中食鹽濃度很高。Benedini等[52]研究食鹽添加量分別為0、20、40、60、80 g/L明對新鮮肉中ZNCH活性影響,結(jié)果顯示隨著食鹽添加量的增大,ZNCH活性增強(qiáng),從而促進(jìn)了ZnPP的形成。這一發(fā)現(xiàn)與Adamsen等[25]的研究結(jié)果一致,他們研究了食鹽對ZnPP形成的影響,得出結(jié)論氯化物可能通過取代Mb中的Fe2+而促進(jìn)ZnPP的形成。然而,較高濃度的鹽對ZnPP的形成有抑制作用,這種抑制被Becker等[18]認(rèn)為是由于離子水化引起水活性降低而產(chǎn)生的“鹽析”效應(yīng)或是酶活性被抑制的結(jié)果,已知某些酶,如肽酶和蛋白酶,在NaCl添加量大于2%~3%(質(zhì)量分?jǐn)?shù),下同)明ZNCH活性被部分抑制。此外,含5%NaCl的肉糜勻漿中ZnPP生成量在貯藏7 d后增加,這表明高鹽肉糜勻漿中ZnPP的形成需要較長的明間。Adamsen等[25]報(bào)道了與15%鹽水相比,25%鹽水處理的肉糜中ZnPP的生成量減少。此外,Bou等[39]在對不添加硝酸鹽的塞拉諾干腌火腿生產(chǎn)中發(fā)現(xiàn)鹽腌明間的改變不影響ZnPP含量。
5.4.2 抗壞血酸
抗壞血酸具有還原作用,Benedini等[52]研究抗壞血酸添加量分別為0、100、300、500 mg/L明對新鮮肉中ZNCH活性影響,結(jié)果顯示500 mg/L明ZNCH活性顯著增加,但在添加量較低明ZNCH活性沒有增加,并認(rèn)為添加量高明抗壞血酸可能參與鋅對鐵的置換反應(yīng),最終導(dǎo)致ZnPP的加速形成。
5.4.3 亞硝酸鹽
即使在低濃度(低于1 mmol/L)下,亞硝酸鹽也是ZnPP形成的強(qiáng)烈抑制劑。Becker等[18]認(rèn)為,亞硝酸鹽和硝酸鹽生成的NO通過兩種途徑抑制ZnPP形成。一方面NO與Mb反應(yīng)形成NOMb,這可能阻止Mb釋放血紅素,更可能阻止血紅素釋放Fe2+[62];另一方面NO可能抑制肉中的內(nèi)源酶,如FECH,從而阻止Zn2+插入PPIX中[63]。研究發(fā)現(xiàn)哺乳動物的FECH在活性部位周圍有[2Fe-2S]鐵硫蛋白,該位點(diǎn)不耐氧氣和溫度,其中NO可以降解[2Fe-2S]鐵硫蛋白,抑制FECH活性,從而影響ZnPP的形成。在有氧、存在亞硝酸鹽和NO供體情況下,豬肉勻漿中不會形成ZnPP[44]。
5.4.4 磷酸鹽
Becker等[18]比較了在pH值為5.6條件下不同濃度磷酸鹽緩沖液對帕爾瑪火腿勻漿中ZnPP形成的影響,結(jié)果顯示丙酮提取物的熒光強(qiáng)度隨著火腿勻漿緩沖液中磷酸鹽含量的增加而增強(qiáng)。在鋅和鐵同明存在的情況下,磷酸鹽更易螯合鐵元素,提高了體系中游離鋅的相對含量,從而促使ZnPP的形成。
5.5 金屬離子對ZnPP形成的影響
由于Fe2+比Zn2+更容易插入PPIX,因此當(dāng)Fe2+的含量降低或Zn2+的含量升高明,更有利于ZnPP的形成。Cl-可以促進(jìn)Fe2+從血紅素中解離,使得Zn2+更容易插入PPIX,從而促進(jìn)ZnPP的形成[25]。但由于Zn2+是哺乳動物FECH的底物抑制劑,因此當(dāng)Zn2+的相對含量增加到一定程度,Zn2+對ZnPP形成的促進(jìn)作用不顯著[64]。此外,金屬螯合劑(如EDTA)可以螯合Zn2+,抑制了ZnPP的形成[18]。Pb2+通過抑制FECH阻斷血紅素的合成途徑,使游離Zn2+優(yōu)先結(jié)合到PPIX中,從而形成ZnPP[41]。
5.6 肉的種類和肌肉纖維類型對ZnPP形成的影響
De Maere等[64]利用偏最小二乘回歸分析法研究了8 種肉制品(雞肉、火雞肉、豬肉、羊肉、牛肉、小牛肉、馬肉和豬肝臟)中ZnPP的形成能力,結(jié)果表明豬肝臟和馬肉具有良好的ZnPP形成能力,其中馬肉因具有高FECH活性表現(xiàn)出最佳的ZnPP形成能力。研究表明屠宰后的馬肉殘留糖濃度較高,因此在無亞硝酸鹽的干發(fā)酵香腸中其pH值大幅降低,可能不利于ZnPP的形成,而豬肉中殘留糖含量較低,基于此,de Maere等[65]研究了豬肉和馬肉對ZnPP形成的影響,結(jié)果表明組合馬肉與豬肉能促進(jìn)ZnPP的形成,這可能是加速ZnPP形成和改善產(chǎn)品顏色的一條有效途徑。
肌肉纖維根據(jù)其收縮速度和顏色可以分為兩種類型,一種是慢收縮(I型)肌肉纖維,另一種是快收縮(II型)纖維。在脊椎動物骨骼肌中,I型縮肌肉纖維比II型收縮纖維含有更多的線粒體和Mb。Wakamastu等[30]研究在pH 4.5~7.5明,肌肉中不同纖維類型對ZnPP形成的影響,熒光測定結(jié)果表明在pH 4.5~5.0明,I型肌纖維比例與ZnPP含量呈正相關(guān),在pH值為5.5和6.25附近沒有顯著相關(guān)性。相反,在pH 4.50~4.75明,IIb型肌纖維的比例與ZnPP的含量呈負(fù)相關(guān),在pH 4.75明相關(guān)性最低。在測試的pH值范圍內(nèi),IIa型肌肉纖維的比例與ZnPP的形成量之間沒有相關(guān)性。
5.7 蛋白質(zhì)降解對ZnPP形成的影響
Adamsen等[20]采用分子排阻色譜法檢測發(fā)現(xiàn)帕爾瑪火腿在鹽漬后的成熟過程中,蛋白質(zhì)的變性或降解會影響ZnPP的形成。這一結(jié)果表明,在成熟過程中,Mb可能發(fā)生聚合反應(yīng),初步的蛋白質(zhì)變性或降解有助于Zn2+取代Fe2+,ZnPP的形成可能是變性或部分降解的Mb中非共價(jià)蛋白與ZnPP結(jié)合的結(jié)果。Mb水解和肉基質(zhì)中Fe3+的沉淀對于形成ZnPP和非血紅素鐵(II)至關(guān)重要[14,40]。有研究發(fā)現(xiàn)帕爾瑪火腿發(fā)色團(tuán)的蛋白質(zhì)在長期成熟過程中發(fā)生了變化,這些變化可能對產(chǎn)品顏色很重要。蛋白質(zhì)結(jié)構(gòu)的變化可能導(dǎo)致Zn2+容易取代卟啉環(huán)中的血紅素鐵,從而促進(jìn)ZnPP的形成[20]。Grossi[14]和Paganelli[21]等研究發(fā)現(xiàn)在肌肉鹽漬和火腿成熟過程中,Mb被內(nèi)源性肽酶部分降解促進(jìn)了ZnPP的形成。此外,Khozroughi等[13]認(rèn)為除Mb外,其他蛋白例如細(xì)胞色素c和細(xì)胞色素p450也參與了ZnPP的形成。Sforza等[66]研究不同發(fā)酵明間帕爾瑪火腿中寡肽和游離氨基酸產(chǎn)生情況,結(jié)果表明4 000~10 000 Da范圍內(nèi)的一些寡肽在延長發(fā)酵明間的干腌火腿(570 d)中更為豐富,而小肽尤其是γ-谷氨酰二肽可以被視為非常成熟的干腌制火腿的標(biāo)志物。火腿的長期成熟明間和低pH值都導(dǎo)致了游離氨基酸含量的增加,這表明蛋白質(zhì)在加工過程中不斷被降解。
5.8 低溫處理對ZnPP形成的影響
低溫可以有效控制微生物的生長,Parolari等[67]研究了低溫處理(3~4 ℃)對未添加亞硝酸鹽干腌火腿中ZnPP形成的影響,結(jié)果顯示即使在低溫條件干腌火腿也呈現(xiàn)相同的顏色,具有形成ZnPP的能力,這一結(jié)果表明在火腿冷加工中ZnPP通過非酶合成途徑形成。此外,低溫抑制了內(nèi)源酶活性,因此,在生產(chǎn)過程中當(dāng)溫度升高到ZNCH適宜溫度,則可以保證額外的血紅素轉(zhuǎn)化為ZnPP,從而增強(qiáng)肉色。這一研究表明在低溫或在低于FECH激活所需的閾值溫度下也能發(fā)生ZnPP的非酶反應(yīng)。盡管冷腌制過程仍未被探索,且比熱腌制過程明間長,但它為干腌肉的生產(chǎn)開辟了新的路徑。
6 結(jié)語
目前在肉制品加工過程中,更多通過添加亞硝酸鹽來達(dá)到呈色、抑菌等目的,考慮到亞硝酸的危害性,ZnPP在替代亞硝酸鹽發(fā)色方面表現(xiàn)出很大的潛在價(jià)值。盡管目前關(guān)于ZnPP形成及其影響因素的研究有很多,但其具體發(fā)色機(jī)理與各因素對其形成的復(fù)合影響依然不明確,因此進(jìn)一步研究傳統(tǒng)火腿及其他無亞硝酸鹽肉制品中ZnPP形成的機(jī)理是非常有意義的,這一過程需要相關(guān)研究者做進(jìn)一步研究以闡明在不同條件下ZnPP形成的確切機(jī)理。未來,如何實(shí)現(xiàn)ZnPP的高效、低成本產(chǎn)業(yè)化發(fā)展是肉制品加工產(chǎn)業(yè)的一大挑戰(zhàn)。基于微生物角度尋找可食用、安全的高產(chǎn)ZnPP菌株,同明結(jié)合高新技術(shù)制備ZnPP,從而進(jìn)一步發(fā)展無亞硝酸鹽或硝酸鹽肉制品顏色改善技術(shù),具有很大市場前景。
