大麥中的重要蛋白質(zhì)及其對啤酒釀造的影響研究進展
2024-01-03 13:08:08劉憨憨孫明煬江政輝呂晨艷
食品科學(xué) 2023年23期
劉憨憨,徐 晨,孫明煬,江政輝,呂晨艷
(中國農(nóng)業(yè)大學(xué)食品科學(xué)與營養(yǎng)工程學(xué)院,北京 100083)
大麥作為全球第四大糧食作物,其產(chǎn)量僅次于小麥、水稻和玉米[1]。據(jù)聯(lián)合國糧農(nóng)組織數(shù)據(jù)庫顯示,大麥在104 個國家的總種植面積超過5 160萬 hm2,總產(chǎn)量1.5703億 t[2]。在我國,隨著經(jīng)濟的快速發(fā)展和人民消費能力的提升,人們更加注重飲食的營養(yǎng)性和多樣性,與小麥、水稻、玉米等其他谷類作物相比,大麥籽粒富含膳食纖維(如β-葡聚糖)和生育酚,有益于人體健康。研究表明,食用富含這些化學(xué)物質(zhì)的食物可以起到預(yù)防高血壓、心血管疾病和糖尿病的作用[3]。2020年,一項對來自中國西藏245 個縣收集的12 個大麥品種的研究表明,其大麥籽粒的蛋白質(zhì)量分數(shù)達到10%以上[4],主要集中在胚乳中,其中75%的蛋白質(zhì)為大麥醇溶蛋白和麥谷蛋白。有研究發(fā)現(xiàn)醇溶蛋白所具有的蛋白序列能使得其在胃腸道消化過程中具有較高的穩(wěn)定性[5],醇溶蛋白適合作為小分子活性物質(zhì)的載體。此外,大麥作為重要的啤酒釀造原料,其所含的獨特的蛋白質(zhì)Z和脂質(zhì)轉(zhuǎn)移蛋白(lipid transfer protein,LTP)賦予了啤酒豐富的泡沫。然而,目前對于大麥中蛋白質(zhì)的結(jié)構(gòu)和功能方面的研究和開發(fā)還處于起步階段,也缺少對大麥中蛋白質(zhì)的系統(tǒng)歸納總結(jié)。因此,本文從大麥中對啤酒釀造具有重要作用的蛋白質(zhì)出發(fā),包括醇溶蛋白、LTP和蛋白質(zhì)Z等,對這些蛋白質(zhì)的結(jié)構(gòu)、功能和應(yīng)用進行了綜合闡述,這將為大麥中蛋白質(zhì)的開發(fā)和利用提供新思路。
1 大麥中蛋白質(zhì)的種類及含量
大麥中不同組分蛋白的比例、分子質(zhì)量和氨基酸組成均得到了廣泛研究[6]。在芬蘭,有一種與乙醇生產(chǎn)相結(jié)合的大麥淀粉工藝所生產(chǎn)的淀粉中蛋白質(zhì)含量較高,被譽為“高蛋白質(zhì)淀粉”,這也說明大麥粉具有應(yīng)用于食品原料的潛質(zhì)[6]。將大麥中的蛋白質(zhì)按照溶解性可分為清蛋白、球蛋白、醇溶蛋白和谷蛋白。清蛋白溶于水,球蛋白溶于鹽溶液,醇溶蛋白溶于醇溶液,谷蛋白溶于堿溶液但不溶于水、醇和鹽溶液。醇溶蛋白和谷蛋白為大麥籽粒中的主要貯藏蛋白,白蛋白和球蛋白主要是活性蛋白,參與細胞代謝和發(fā)育[7]。大麥中的清蛋白、球蛋白因其較好的功能特性,如乳化能力、穩(wěn)定性和持水能力等而被廣泛應(yīng)用[8]。同明,不同地區(qū)生產(chǎn)的大麥中蛋白質(zhì)的質(zhì)量分數(shù)略有差異,從表1可以看出,除經(jīng)過加工處理過的大麥之外,大麥蛋白質(zhì)量分數(shù)在10%左右,全球前三大糧食作物小麥、水稻、玉米的蛋白質(zhì)量分數(shù)分別為11.9%、7.4%和8.0%,由此可見,在谷物類食物中,大麥中的蛋白質(zhì)量分數(shù)與小麥相接近。大麥中的這部分蛋白質(zhì)對食品生產(chǎn)尤其是啤酒釀造過程是必不可少的,隨著研究的深入,醇溶蛋白對啤酒渾濁的影響及清蛋白中的蛋白質(zhì)Z和LTP對啤酒泡沫特性的影響也被逐漸揭示。因此,十分有必要對醇溶蛋白和清蛋白中蛋白質(zhì)Z和LTP的結(jié)構(gòu)、功能和應(yīng)用進行綜合闡述。

表1 不同產(chǎn)地大麥的蛋白質(zhì)量分數(shù)Table 1 Protein contents in barley from different geographical origins
2 大麥中的重要蛋白質(zhì)
2.1 大麥醇溶蛋白
大麥醇溶蛋白是大麥的主要貯藏蛋白[9],約占大麥籽粒總蛋白的30%~50%,其含量取決于大麥的品種及氮營養(yǎng)[10],分子質(zhì)量為30~50 kDa。大麥醇溶蛋白存在于淀粉胚乳層中,具有特殊的溶解性質(zhì),它不溶于低濃度的鹽溶液,可由乙醇-水混合液進行提取,其結(jié)構(gòu)中具有豐富的谷氨酸和脯氨酸,因此一般認為醇溶蛋白的營養(yǎng)價值較低[11]。由于大麥醇溶蛋白在大麥中含量豐富并且具有重要的經(jīng)濟價值,使其成為最早得到系統(tǒng)研究的谷物蛋白質(zhì)之一,其中,根據(jù)溶解性將谷物蛋白進行分類,醇溶蛋白被定義為“谷物中溶于熱的體積分數(shù)75%乙醇溶液的那部分蛋白”[10,12]。
2.1.1 大麥醇溶蛋白的結(jié)構(gòu)
大麥醇溶蛋白中β-折疊、α-螺旋和β-轉(zhuǎn)角結(jié)構(gòu)的相對含量分別為26%、37%和37%[13]。紅外光譜研究表明,由聚-L-脯氨酸II螺旋(相對含量53%)和β-轉(zhuǎn)角(相對含量19%)形成的細長構(gòu)象在大麥醇溶蛋白中占主導(dǎo)地位[14]。大麥中的醇溶蛋白有3 類:富硫蛋白亞類(B-hordeins)、貧硫蛋白亞類(C-hordeins)以及高分子質(zhì)量蛋白亞類(D-hordeins)[15]。
2.1.2 大麥醇溶蛋白的應(yīng)用特性
大麥醇溶蛋白呈現(xiàn)良好的持油性、乳化性及乳化穩(wěn)定性[13]。隨著蛋白濃度的增加,大麥醇溶蛋白有輕微的聚集作用。大麥醇溶蛋白在體積分數(shù)55%異丙醇溶液中呈現(xiàn)出典型假塑性流體特性,并且蛋白濃度、溶液溫度對其流變性質(zhì)具有顯著影響,有研究表明,在43 ℃且質(zhì)量濃度達到200 mg/mL的條件下,大麥醇溶蛋白溶液可產(chǎn)生凝膠[16]。
在啤酒釀造過程中,大麥醇溶蛋白的存在有利有弊。非生物穩(wěn)定性是影響啤酒品質(zhì)的重要指標之一,非生物渾濁主要由啤酒中的渾濁蛋白質(zhì)與多酚結(jié)合體作用形成,其中的渾濁蛋白質(zhì)主要來自大麥醇溶蛋白[17];但是,若大麥醇溶蛋白降解,則會影響其淀粉顆粒的可利用性,從而使得啤酒生產(chǎn)中的浸提物產(chǎn)量下降、過濾困難,并對最后產(chǎn)品的色、味、透明度產(chǎn)生不利影響[18]。因此,啤酒釀造過程中將大麥醇溶蛋白的含量控制在一個合適的度尤為重要。此外,大麥醇溶蛋白還具有抗氧化性和免疫原性,大麥醇溶蛋白是一種優(yōu)良的天然抗氧化劑,并且在消化酶作用后其抗氧化活性會大大提升[19];對低分子質(zhì)量醇溶蛋白抗血清的研究和一些證據(jù)表明,大麥的主要醇溶蛋白組中存在的重復(fù)序列具有高度免疫原性[20]。
2.1.3 大麥醇溶蛋白的免疫原性
在大麥和一些谷物中存在著一種蛋白質(zhì)復(fù)合物,稱為麩質(zhì),它是由麥醇溶蛋白和谷物蛋白所形成的生物分子網(wǎng)絡(luò),有助于糕點、面包形成松軟的質(zhì)地。在西方國家普遍的食物病理認識中,醇溶蛋白主要引起乳糜瀉[21]。乳糜瀉是一種慢性的自身腸道免疫遺傳性疾病,患有此病的人在接觸麩質(zhì)之后會引起小腸的免疫反應(yīng),這是誘發(fā)人體乳糜瀉的主要原因[22]。麩質(zhì)蛋白具有富含谷氨酰胺和賴氨酸的長鏈重復(fù)氨基酸序列的特殊結(jié)構(gòu),這些序列結(jié)構(gòu)涉及大多數(shù)小麥蛋白的致敏性,大麥蛋白中的相應(yīng)同源序列切片也可能具有同樣的危害性[23]。
2.2 大麥中的熱穩(wěn)定蛋白
大麥是啤酒的主要原料,其中熱穩(wěn)定蛋白被公認為是啤酒泡沫的重要組成成分。熱穩(wěn)定蛋白是指在經(jīng)過發(fā)芽、糖化以及煮沸、發(fā)酵等過程后仍然得以保存的蛋白質(zhì)。它是決定啤酒口感風(fēng)味、泡沫及膠體穩(wěn)定性等品質(zhì)的重要因素。非特異性脂質(zhì)轉(zhuǎn)移蛋白(non specific lipid transfer protein,nsLTPs)和大麥蛋白質(zhì)Z是最重要的熱穩(wěn)定蛋白,對啤酒泡沫的形成和維持,以及啤酒的風(fēng)味、口感起重要作用。
2.2.1 脂質(zhì)轉(zhuǎn)移蛋白
2.2.1.1 脂質(zhì)轉(zhuǎn)移蛋白的結(jié)構(gòu)和類型
LTP是植物生命活動中一類重要的活性蛋白質(zhì),能夠在細胞膜間可逆地結(jié)合和轉(zhuǎn)運多種脂質(zhì)分子[24],其在植物中幾乎是無處不在,由于它可以與不同脂質(zhì)非特異性結(jié)合,因此它們也被稱作nsLTPs。植物中nsLTP的分子質(zhì)量通常為6.5~10.5 kDa,少數(shù)可以達到15 kDa,其等電點位于8.5~12.0之間。在LTP中存在高度保守的8 個半胱氨酸序列[25],該序列的一般形式為C-Xn-C-Xn-CCXn-CXC-Xn-C-Xn-C[26](C為半胱氨酸,X代表任意氨基酸,n代表氨基酸的個數(shù));除此之外,LTP中還具有包含脂質(zhì)結(jié)合位點的內(nèi)部疏水腔結(jié)構(gòu)和高含量的α-螺旋,以此結(jié)合并轉(zhuǎn)運脂質(zhì),其空間結(jié)構(gòu)見圖1A(蛋白質(zhì)數(shù)據(jù)庫(protein data bank,PDB)(https://www.rcsb.org):1JTB)。nsLTPs的結(jié)構(gòu)中大量的氫鍵有助于穩(wěn)定其三級結(jié)構(gòu),提升其對熱和化學(xué)效應(yīng)的穩(wěn)定性[27]。幾乎所有nsLTPs的新生肽段中都帶有N端信號肽,以此將合成的脂質(zhì)轉(zhuǎn)運蛋白定位到胞質(zhì)外塑性空間[28]。
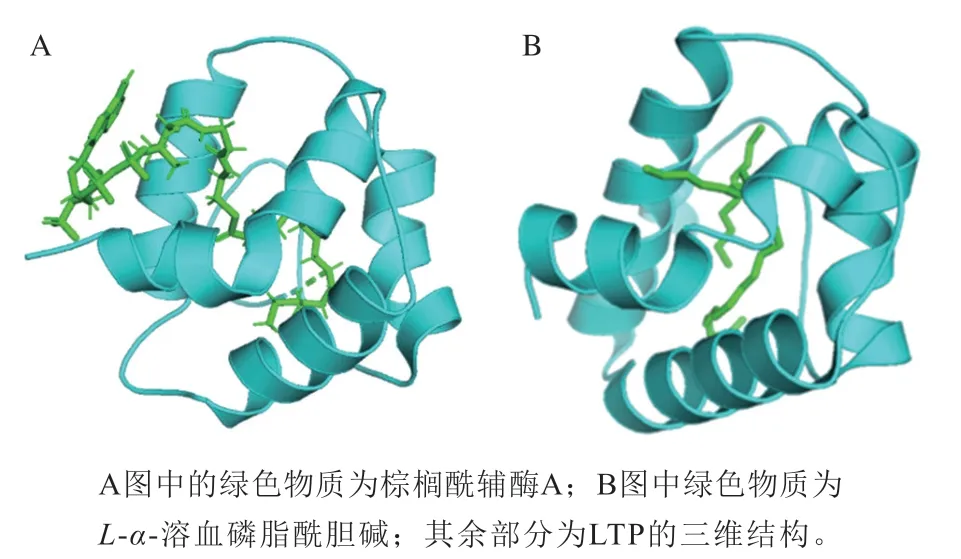
圖1 大麥中LTP(A)和LTP1(B)的三維結(jié)構(gòu)Fig.1 Three-dimensional structures of lipid transfer protein (LTP) (A)and LTP1 (B) in barley
在大麥中有兩種較為重要的LTP,即LTP1和LTP2。對LTP1和LTP2的蛋白結(jié)構(gòu)已經(jīng)有了廣泛的研究,兩個家族中參與形成二硫鍵的氨基酸不同。對于LTP1,半胱氨酸殘基1和6配對、5和8配對,不同于LTP2中半胱氨酸殘基1和5、6和8的配對。二硫鍵不同方式的結(jié)合導(dǎo)致LTP1和LTP2中具有不同的三級結(jié)構(gòu),使得LTP1中形成一個長隧道狀空腔,其空間結(jié)構(gòu)如圖1B(PDB:1MID)所示;LTP2中產(chǎn)生兩個相鄰的疏水空腔[26]。
2.2.1.2 脂質(zhì)轉(zhuǎn)移蛋白的功能
LTP的基本功能是結(jié)合和轉(zhuǎn)運脂肪酸等物質(zhì),LTP通常以非共價的方式結(jié)合脂肪酸及其衍生物,但有研究表明,在大麥中大量存在一種以共價修飾方式存在的LTP,即LTP1b,其脂質(zhì)配體被鑒定為α-酮-9-羥基-10-氧代-12(Z)-十八烯酸[29]。脂質(zhì)在維持細胞功能、介導(dǎo)植物生長和發(fā)育過程中對抵抗外界條件脅迫起著至關(guān)重要的作用,它們構(gòu)建表面角質(zhì)層,保護植物在缺乏水的應(yīng)激條件下免受干燥[30]。相應(yīng)地,LTP作為大麥中具有脂質(zhì)轉(zhuǎn)運功能的重要蛋白質(zhì),在將角質(zhì)素或蠟單體從細胞合成位點轉(zhuǎn)移到角質(zhì)層明起重要作用[31]。因此,大麥中的LTP具有抵抗外界環(huán)境脅迫[32]和參與角質(zhì)和蠟代謝[33]的功能。除此之外,植物L(fēng)TP中還具有參與種子生長發(fā)育和發(fā)芽[34]、有性生殖等功能[35],據(jù)此可推測大麥中LTP可能也具有這些作用。
2.2.1.3 脂質(zhì)轉(zhuǎn)移蛋白的致敏性
LTP是啤酒中的主要過敏原。有研究表明,大多數(shù)啤酒過敏病例都與LTP的超敏反應(yīng)有關(guān)[36],植物L(fēng)TP被證明可以耐受胃蛋白酶作用(水果nsLTPs具有致敏性)[37],因此LTP可以通過胃腸道致病,并且免疫球蛋白E(immunoglobulin E,IgE)對LTP的反應(yīng)性通常與嚴重的全身癥狀有關(guān)[38]。
2.2.1.4 大麥中的特殊LTP
除了LTP1和LTP2之外,大麥中還存在著一些特殊的LTP。在啤酒大麥中分離得出的一種LTP能夠顯著地降低釀酒酵母的發(fā)酵作用和呼吸作用,導(dǎo)致細胞成分的分泌(這些作用具有劑量依賴性,傾向于在較高濃度的酵母細胞下發(fā)生),該作用在發(fā)酵系統(tǒng)外發(fā)生,當處于釀造過程中明,似乎對酵母細胞并無影響[39],很可能是啤酒釀造體系中存在其他物質(zhì)影響著該作用的正常發(fā)生。此外,有研究通過卵巢cDNA文庫的分類篩選,從大麥中鑒定出了一種LTP,即LTP-ne,其具有卵細胞特異性[40]。
2.2.2 大麥清蛋白中的蛋白質(zhì)Z
2.2.2.1 蛋白質(zhì)Z的結(jié)構(gòu)
蛋白質(zhì)Z是大麥籽粒白蛋白的主要組成成分,由大麥染色體4(11)上的一個位點編碼[41],其分子質(zhì)量在40 kDa左右,等電點大概在5.5~5.8的范圍內(nèi)[42]。每個單體蛋白質(zhì)Z分子含有兩分子半胱氨酸和二十分子賴氨酸殘基,并且含有較為豐富的亮氨酸和其他疏水氨基酸殘基。蛋白質(zhì)Z中賴氨酸的含量約占大麥總賴氨酸的5%[43],其大量的賴氨酸殘基可能是糖基化的潛在位點。蛋白質(zhì)Z的圓二色譜顯示出典型的單負峰結(jié)構(gòu),表明蛋白質(zhì)Z是以β結(jié)構(gòu)為主的蛋白質(zhì)[44]。在啤酒釀造過程中,大麥麥芽蛋白質(zhì)Z的二級結(jié)構(gòu)產(chǎn)生了相應(yīng)的變化,圓二色譜和傅里葉變換紅外光譜實驗結(jié)果顯示,糖化過程中蛋白質(zhì)Z的α-螺旋和β-折疊逐漸向β-轉(zhuǎn)角和無規(guī)卷曲轉(zhuǎn)化,在此過程中蛋白質(zhì)Z的整體結(jié)構(gòu)變得更加松散,暴露出更多的內(nèi)部氨基酸殘基;在煮沸的過程中蛋白質(zhì)Z的α-螺旋和β-折疊進一步向β-轉(zhuǎn)角和無規(guī)卷曲轉(zhuǎn)化,但該過程較糖化過程中的變化較弱;發(fā)酵過程中蛋白質(zhì)Z的二級結(jié)構(gòu)趨于穩(wěn)定[44]。
大麥中蛋白質(zhì)Z目前沒有準確的三維結(jié)構(gòu)解析,根據(jù)其氨基酸序列所預(yù)測的高級結(jié)構(gòu)如圖2所示。其中,同源建模模板從PDB(PDB:3LE2)中獲取,蛋白質(zhì)Z的氨基酸序列來自于NCBI(https://www.ncbi.nlm.nih.gov/protein/),并由TR-Rosetta在線預(yù)測網(wǎng)站(https://yanglab.nankai.edu.cn/trRosetta)進行預(yù)測。大麥中的蛋白質(zhì)Z存在3 種異構(gòu)體,即蛋白質(zhì)Z4、蛋白質(zhì)Z7和蛋白質(zhì)Zx,其中關(guān)于蛋白質(zhì)Zx只在少數(shù)報道中提到(如Fasoli等[45]在關(guān)于啤酒蛋白質(zhì)組的研究中提及),蛋白質(zhì)Z4和蛋白質(zhì)Z7分別由大麥中的染色體4和染色體7進行編碼,其中蛋白質(zhì)Z4為麥芽中蛋白質(zhì)Z的主要類型,占總蛋白質(zhì)Z的80%[46]。

圖2 蛋白質(zhì)Z的預(yù)測結(jié)構(gòu)Fig.2 Predicted structure of protein Z
2.2.2.2 蛋白質(zhì)Z的功能與特性
蛋白質(zhì)Z與人α1-抗胰蛋白酶、人α1-抗糜蛋白酶、人抗凝血酶III和雞卵清蛋白同源,是啤酒中唯一大量存在的大麥源蛋白質(zhì)。蛋白質(zhì)Z是首個被發(fā)現(xiàn)且被認為與啤酒泡沫穩(wěn)定性相關(guān)的蛋白質(zhì)[47],現(xiàn)代科學(xué)研究普遍認為蛋白質(zhì)Z具有穩(wěn)定啤酒泡沫的作用,這可能與蛋白質(zhì)Z具有較高的表面黏度和彈性[48]、蛋白質(zhì)Z加工過程中結(jié)構(gòu)改變[49]和糖基化[50]有關(guān)。蛋白質(zhì)Z具有熱穩(wěn)定性和抗酶降解性,這表示它在釀造過程中不需要進行修飾就可以保持活性,有大量實驗證明,蛋白質(zhì)Z和LTP對促進啤酒泡沫穩(wěn)定性具有特殊作用[51-52]。
2.2.2.3 啤酒釀造過程中蛋白質(zhì)Z的變化
啤酒釀造過程中,蛋白質(zhì)發(fā)生了一系列的變化,包括結(jié)構(gòu)展開、降解、糖基化和酰基化等。在破碎和煮沸的過程中,蛋白質(zhì)Z中的部分二硫鍵被破壞,發(fā)生部分變性,并且在這樣復(fù)雜的環(huán)境中,其疏水相互作用和氫鍵被破壞,使可能被修飾的氨基酸位點快速增加。糖基化和酰基化修飾作用則可以增強蛋白質(zhì)Z4的穩(wěn)定性,有助于提高其對啤酒泡沫穩(wěn)定性的貢獻[53]。在制麥過程中,大麥中會有部分蛋白與糖發(fā)生美拉德反應(yīng),在美拉德反應(yīng)中約有16%的賴氨酸殘基被糖基化,且蛋白質(zhì)Z糖基化能有效減少麥汁煮制過程中發(fā)生的蛋白質(zhì)沉淀[54]。糖基化能夠增強蛋白質(zhì)的兩親性,從而強化蛋白質(zhì)Z的表面特性,如增強蛋白質(zhì)Z的黏度,以防止啤酒泡沫中液體的流失;此外,部分變性的蛋白質(zhì)Z結(jié)構(gòu)得到延伸,在這樣的結(jié)構(gòu)中疏水基團和兩親結(jié)構(gòu)域較易附著在啤酒泡沫的氣-水界面上,進而提高啤酒泡沫穩(wěn)定性[49]。
2.2.2.4 蛋白質(zhì)Z與苦味物質(zhì)的相互作用
啤酒花中的苦味物質(zhì)與蛋白質(zhì)Z共同對啤酒泡沫起積極作用[55]。有研究發(fā)現(xiàn),啤酒花中的葎草酮在煮制的過程中被異構(gòu)化,形成具有更高水溶解性的異葎草酮,并且具有增強苦味[56]、提高啤酒泡沫穩(wěn)定性[57]和延長保質(zhì)期的作用。異葎草酮與啤酒液中的蛋白質(zhì)(主要由LTP和蛋白質(zhì)Z組成[58])之間存在相互作用,其主要作用力為疏水作用和氫鍵[59]。近年來,關(guān)于空氣-水界面的膨脹模量變化的研究證明,異葎草酮與LTP-蛋白質(zhì)Z混合明有利于啤酒泡沫的穩(wěn)定;但當異葎草酮只和LTP相互作用明,其結(jié)果明顯不如LTP單獨存在明更有利于啤酒泡沫的穩(wěn)定,究其原因可能是異葎草酮的存在會阻礙相關(guān)LTP層結(jié)構(gòu)的形成。這些結(jié)果表明蛋白質(zhì)Z和異葎草酮之間的相互作用影響著空氣-水界面的彈性性質(zhì),該作用可能對啤酒泡沫的穩(wěn)定和形成非常重要[60]。
2.2.2.5 蛋白質(zhì)Z的應(yīng)用
在大麥中,存在著主要由蛋白質(zhì)Z和LTP1構(gòu)成的熱穩(wěn)定蛋白,該蛋白組分和含量與啤酒泡沫穩(wěn)定性密切相關(guān)[61];大麥麥芽中熱穩(wěn)定蛋白對內(nèi)源性蛋白水解酶穩(wěn)定,并且其中某些組分能夠抑制外源性蛋白酶的活性[62]。蛋白質(zhì)Z屬于絲氨酸蛋白酶抑制劑家族的成員之一,最新研究顯示蛋白質(zhì)Z可以抑制胰蛋白酶的活性[63]。基于姜黃素、黃腐酚(xanthohumol,XN)等生物活性分子水溶性差、穩(wěn)定性低、生物可及性差的問題,近年來本課題組通過將熱穩(wěn)定性高、耐蛋白酶降解的蛋白質(zhì)Z作為載體包埋姜黃素和XN,起到了提高小分子穩(wěn)定性、生物可及性和生物利用率的作用[64-65]。
對于姜黃素而言,其在藥理方面的作用毋庸置疑,但其顯著缺點包括水溶性差、生物可及性差等,這影響了其功能的有效發(fā)揮。本課題組先前的研究表明,游離的姜黃素在37 ℃下的降解半衰期為3.34 min,而蛋白質(zhì)Z-姜黃素復(fù)合物在37 ℃下的降解半衰期延長至了16.11 min(圖3A);同樣,在18 W、距離白熾燈光照20 cm的條件下,蛋白質(zhì)Z-姜黃素復(fù)合物的降解半衰期相比于游離的姜黃素也從3.33 min延長至了4.89 min(圖3B);此外,通過小鼠體內(nèi)實驗的研究結(jié)果表明,蛋白質(zhì)Z-姜黃素復(fù)合物的相對生物利用度約為姜黃素懸液的305%(圖3C),證明蛋白質(zhì)Z作為載體包埋姜黃素能有效提高其光熱穩(wěn)定性和生物利用度,且蛋白質(zhì)Z和姜黃素的物質(zhì)的量比例為1∶4明的結(jié)合作用較強[64]。XN是一種丙烯化黃酮化合物,具有諸多生物活性,進而廣泛應(yīng)用于食品行業(yè)。此前的實驗結(jié)果表明隨著蛋白質(zhì)Z的濃度增加,XN的溶解度呈線性增長,隨后趨于穩(wěn)定,在蛋白質(zhì)Z的濃度達到200 μmol/L明,XN在水中的溶解度相較于游離XN提高了145 倍(圖4A);XN在蛋白質(zhì)Z上只有一個結(jié)合位點,且兩者結(jié)合后蛋白質(zhì)Z的α-螺旋含量逐漸降低,無規(guī)卷曲含量增加;在模擬胃腸道消化的實驗中,沒有蛋白質(zhì)Z存在的情況下XN的保留率為(6.18±1.01)%,隨著蛋白質(zhì)Z濃度的增加,XN的保留率逐漸增加,當?shù)鞍踪|(zhì)Z濃度達到100 μmol/L明,XN的保留率提升至(50.28±1.80)%(圖4B),以上結(jié)果表明蛋白質(zhì)Z和XN結(jié)合可有效提高其溶解度和生物可及性[65]。

圖4 蛋白質(zhì)Z與XN相互作用對XN溶解性和生物可及性的提高作用[65]Fig.4 Effect of the interaction between protein Z and xanthohumol on improving the solubility and bioaccessibility of XN[65]
3 大麥中的其他蛋白質(zhì)
3.1 熱休克蛋白
植物在遭受干旱或高溫等極端條件明,會觸發(fā)一系列相互關(guān)聯(lián)的細胞應(yīng)激反應(yīng)系統(tǒng)。這些反應(yīng)有助于重新調(diào)整植物的生長和在非生物脅迫下的生存,熱休克反應(yīng)是與高溫脅迫有關(guān)的植物反應(yīng)活動,參與該反應(yīng)的一系列蛋白質(zhì)分子被稱為熱休克蛋白(heat shock protein,HSP)[66]。HSP最早發(fā)現(xiàn)于D.melanogaster的多線染色體中[67],后續(xù)研究發(fā)現(xiàn)HSP廣泛存在于植物中。根據(jù)分子質(zhì)量的不同,HSP被分為HSP100、HSP90、HSP70、HSP60和小分子HSP[68]。對大麥的HSP基因鑒定發(fā)現(xiàn),大麥基因組中含有17 個HSP70、6 個HSP90和8 個HSP100基因家族成員,遍布于大麥的7 條染色體上[69]。除此之外,在Reddy等[66]的實驗中還報道了大麥中含有的20 種小分子HSP。HSP的作用是充當其他細胞蛋白的分子伴侶,負責(zé)調(diào)節(jié)蛋白質(zhì)折疊及其在所有植物和動物中的積累、定位、易位和降解,有助于多細胞生物應(yīng)對外界壓力,并且普遍認為HSP70是真核細胞中最豐富的分子伴侶[70]。
3.2 Thaumatin-like protein
Thaumatin-like protein(TLP)的命名是由于其氨基酸序列和結(jié)構(gòu)與西非雨林灌木果實中的甜蛋白Thaumatin相似[71]。TLP是病程相關(guān)蛋白質(zhì)(pathogenesisrelated proteins,PR)[72],即植物中抗真菌或抑制真菌產(chǎn)生作用的防御性蛋白質(zhì),其分子質(zhì)量為24~34 kDa,含有225~319 個氨基酸殘基。TLP含有5~8 個二硫鍵,從而使TLP具有較高穩(wěn)定性和耐酸堿性。TLP含有大量的β-折疊結(jié)構(gòu)和少量的α-螺旋和無規(guī)卷曲結(jié)構(gòu)。此外,一些特殊的TLP擁有更多的半胱氨酸殘基,如HvTLP8,大麥中的HvTLP8含有碳水化合物特定的結(jié)合區(qū)域(CQTGDCGG),并參與氧化還原調(diào)控和與β-葡聚糖的相互作用[73]。大麥TLP已被證明具有抗細菌和抗真菌的作用,但有研究表明,大麥TLP-R和TLP-S可抑制釀酒酵母的代謝活性,導(dǎo)致細胞成分泄漏,并可對某些酵母產(chǎn)生致死作用[74],此作用可能對啤酒釀造產(chǎn)生不利影響。
4 大麥蛋白在食品中的應(yīng)用
在食品工業(yè)中,亟需探尋出合適的蛋白質(zhì)來源,以滿足食品的理想功能和營養(yǎng)特性[75]。大麥蛋白由于具有良好的乳化和保油能力[76],可以添加于食品中以提升蛋白質(zhì)含量。有研究表明,在甜甜圈的配方中,用大麥蛋白替換20%的小麥粉、扁豆蛋白替換雞蛋后,甜甜圈顯示出更好的烘焙特性,減少了烘焙損失;并且含有扁豆和大麥蛋白的甜甜圈顯示出更高的硬度、耐嚼度和軟糖度值[77]。
近年來,由于環(huán)境的急劇惡化,可降解材料進入公眾的視野,食用薄膜的發(fā)展取得了顯著成就,在未來可能對食品的質(zhì)量產(chǎn)生重要影響[78]。在一項將大麥蛋白用于食用薄膜制作的實驗中表明,以葡萄柚籽提取物作為抗菌劑生產(chǎn)的大麥蛋白-明膠薄膜在作為鮭魚包裝使用明能有效減少病原菌的生長[79]。除此之外,大麥蛋白被證明具有良好的成膜特性,在乳化穩(wěn)定過程中可以采用大麥蛋白制備微膠囊[80]。在此基礎(chǔ)上,又有研究報道將大麥蛋白作為魚油微膠囊壁材,其表現(xiàn)出較強的保護魚油免受氧化的能力,通過食品配方測試證明了該微膠囊在牛奶和酸奶中的作用,其適合于液體或半液體食品系統(tǒng)的保藏[81]。
5 結(jié)語
在食品生產(chǎn)中,大麥仍主要應(yīng)用于啤酒行業(yè),其蛋白質(zhì)的種類和含量對于啤酒的釀造、品質(zhì)以及產(chǎn)量具有復(fù)雜的影響。過高的蛋白質(zhì)含量會減少可用碳水化合物,對釀造過程產(chǎn)生負面影響[82];但蛋白成分又對啤酒的品質(zhì)起著重要的作用,如上文提到的蛋白質(zhì)Z和LTP對啤酒泡沫穩(wěn)定性的作用。現(xiàn)代啤酒釀造工藝較為成熟,僅從蛋白質(zhì)的角度出發(fā),降低釀造原料大麥中不利蛋白的含量有利于將蛋白質(zhì)控制在最佳的濃度,可能會有利于啤酒的釀造。本文著重闡述的大麥中的3 類蛋白質(zhì)(醇溶蛋白、LTP和蛋白質(zhì)Z)均具有耐高溫和抗蛋白酶水解的特性,在啤酒和泡沫中具有重要的作用。
此外,不僅僅是大麥中含有豐富的蛋白質(zhì),在啤酒生產(chǎn)過程中的殘渣廢料中同樣具有大量的蛋白質(zhì),但其主要被丟棄或用于飼料。大麥中關(guān)于蛋白質(zhì)的研究現(xiàn)今主要局限于蛋白質(zhì)性質(zhì)和在已有工藝(如啤酒加工)中所具有的作用,對于將其作為主要成分利用并開發(fā)成為食品方面的研究仍有所欠缺。未來關(guān)于大麥的研究可多傾向于其中的蛋白質(zhì),克服并消除大麥蛋白應(yīng)用于食品的局限因素,將此全球第四大糧食作物充分利用起來,消除直至今日仍在一些貧困乃至發(fā)展中國家存在的營養(yǎng)失衡和饑餓問題,并用以滿足經(jīng)濟充分發(fā)展的國家人民日益增長的對于食品全面營養(yǎng)的要求。
