江西小流域范圍稻田土壤真菌群落分布特征及其影響因素
2024-01-17 00:00:00郭雨婷劉亞軍吳永明段亦鵬何石福鄧濤李榮富
農業環境科學學報 2024年12期

關鍵詞:小流域;稻田土壤;理化性質;重金屬;真菌群落
中圖分類號:S154.3 文獻標志碼:A 文章編號:1672-2043(2024)12-2974-10 doi:10.11654/jaes.2024-0048
水稻作為我國第一大糧食作物,其產量和品質與土壤性質息息相關[1]。微生物是土壤生態系統的重要組成部分,在促進土壤養分循環和植物生長方面發揮著重要作用[2-3]。真菌是土壤中的主要分解者之一,對環境變化極度敏感,與土壤環境有著復雜的相互作用,其結構組成、多樣性是反映土壤質量和生態系統變化的重要指標[4]。因此,了解稻田土壤真菌群落分布特征與驅動因子對保持土壤健康和農業發展具有重要意義。
近年來,農田生態系統作為農業綠色可持續發展的重要基礎,農田土壤微生物群落結構分布與驅動因素逐漸成為國內外專家學者們的研究重點[5]。基于不同尺度下的土壤微生物分布規律及其驅動因子存在顯著差異[6],在大空間尺度下氣候、空間距離、緯度等是影響微生物群落空間分布的關鍵因素[7-9],而小流域尺度下(1~100 km)氣候、空間距離、緯度等差別較小,微生物群落分布特征規律更與土壤環境、農田管理措施和人為活動有關[10-11]。研究發現,土壤pH值和土壤養分如有機質、全氮、全磷等在一定程度上會影響土壤微生物群落結構[12-13]。農田管理措施(施肥方式、種植制度、耕作模式)的變化會影響土壤結構及土壤有機碳的穩定機制,進而對土壤微生物活性產生影響[14]。有研究者指出,礦山開采及金屬冶煉帶來的重金屬污染對土壤微生物豐度和功能具有持續的抑制作用,且與未污染土壤的真菌群落組成差異明顯[15-16]。鑒于土壤微生物影響因素的復雜性及細菌與真菌菌體結構和代謝物質的差異性,目前研究大多針對大空間尺度范圍下土壤細菌群落的影響因子進行分析,而對小流域尺度下土壤真菌群落差異性及影響因素的了解十分有限。
據此,本研究以萬年縣范圍內稻田土壤為研究對象,排除大尺度上地理距離和氣候等因素,深入分析土壤的理化性質、重金屬元素以及真菌群落的分布特征,探究江西小流域范圍內影響稻田土壤真菌的關鍵環境因子。研究結果將有助于稻田土壤的恢復與穩定,為創建生態友好型水稻種植模式提供理論指導。
1 材料與方法
1.1 研究區概況
萬年縣(28°30′15″~28°54′05″N,116°46′48′′~117°15′10″E)位于江西省東北部、鄱陽湖東南岸,總面積約1 140 km2[17]。全縣地形以低山、丘陵為主,地勢東南高西北低,珠溪河自南向北穿流而過匯入樂安河,具有“六山(丘)一水二分田”的地貌特點,形成了典型的相對封閉的小流域農業生態環境。該地區屬于亞熱帶季風氣候,四季冷熱分明,年均氣溫17.4 ℃,年均降水量1808.0 mm,年均日照時數1 803.5 h。萬年縣地區農業發達,是江西省重要的水稻種植區,享有“中國貢米之鄉”的美譽。同時萬年縣礦產資源豐富,主要有金、銀、砷、鉛等。
1.2 樣品采集與處理
稻田土壤布樣原則依據《農田土壤環境質量監測技術規范》(NY/T 395—2012)和《土壤環境監測技術規范》(HJ/T 166—2004)進行。選擇萬年縣內主要水稻種植區作為采樣區域,沿著珠溪河下游、中游、上游空間分布情況分為珠田北、珠田南、上訪西、上訪東、裴梅東和裴梅西6個區域。區域水稻種植情況如下:施肥為三元復合肥(氮、磷、鉀各15%),約600 kg·hm-2,基施和追肥各占50%;水稻品種為裴梅鎮西種植晚秈稻“塢源早”,其他區域主要種植早秈617、早秈902、陵兩優265和贛菌稻1號等當地常見品種;耕作模式為一年兩季、機械耕作,秸稈還田率gt;90%。共布設48個土壤采樣點,樣點東西跨度約16 km,南北跨度約23km,采樣點位分布見圖1。為了避免當季管理措施的短暫影響,在基肥施撒前進行土壤樣品采集。在每個采樣點,采用5點采樣法使用土鉆采集深度為0~20cm的表層土壤(土壤質量gt;500g),將其混勻后作為1份土壤樣品。土壤樣品裝入密封袋保存,低溫條件運至實驗室,一份經自然風干、研磨、過篩后用于土壤基本理化性質和重金屬的測定,另一份保存在-80 ℃用于土壤真菌高通量測序分析。
1.3 土壤理化性質和重金屬的測定
土壤含水量(SM)采用烘干差質量法測定;顆粒組成(黏粒、粉粒、砂粒)采用激光衍射法測定;pH采用電位法(水土比2.5∶1)測定;陽離子交換量(CEC)采用乙酸銨交換-蒸餾滴定法測定;有機質(OM)采用重鉻酸鉀外加熱法測定;總氮(TN)和水解氮(AN)采用凱氏定氮儀測定;總磷(TP)用氫氧化鈉堿熔-鉬藍比色法測定;有效磷(OP)采用聯合浸提-比色法測定;速效鉀(AK)采用乙酸銨提取-火焰光度法測定;金屬元素(Cd、Cr、Pb、Zn、Cu、Hg、Ni 和As)均采用電感耦合等離子體質譜法測定。
1.4 DNA提取與高通量測序分析
利用MOBIO的強力土壤DNA提取試劑盒(Power?Soil? DNA Isolation Kit)提取樣品總DNA,利用凝膠電泳檢測DNA完整性,并通過NanoDrop2000檢測DNA濃度和純度。基于Illumina Miseq PE300測序平臺進行高通量測序,選擇引物ITS1F(5′-CTTGGTCATT?TAGAGGAAGTAA-3′)和ITS2R(5′-GCTGCGTTCTTCATCGATGC-3′)擴增真菌的ITS1~ITS2 區。測序原始序列經質控、拼接、篩選、去除嵌合體等步驟獲得的有效序列用于物種注釋等后續信息學分析。
1.5 數據統計與分析
數據分析和圖表繪制采用Excel 2016 和R 語言(https://www.r-project.org/)完成。采用單因素方差分析(ANOVA)評價結果的顯著性。基于Bray-Curtis距離的主坐標分析(PCoA)和地理距離衰減分析使用R語言vegan包展示了真菌群落結構概況和真菌群落隨距離變化情況。通過Mantel相關分析和相關性Heat?map分析(Spearman等級相關系數)探究土壤環境因子和真菌群落的關系。
2 結果與分析
2.1 土壤理化參數及重金屬元素
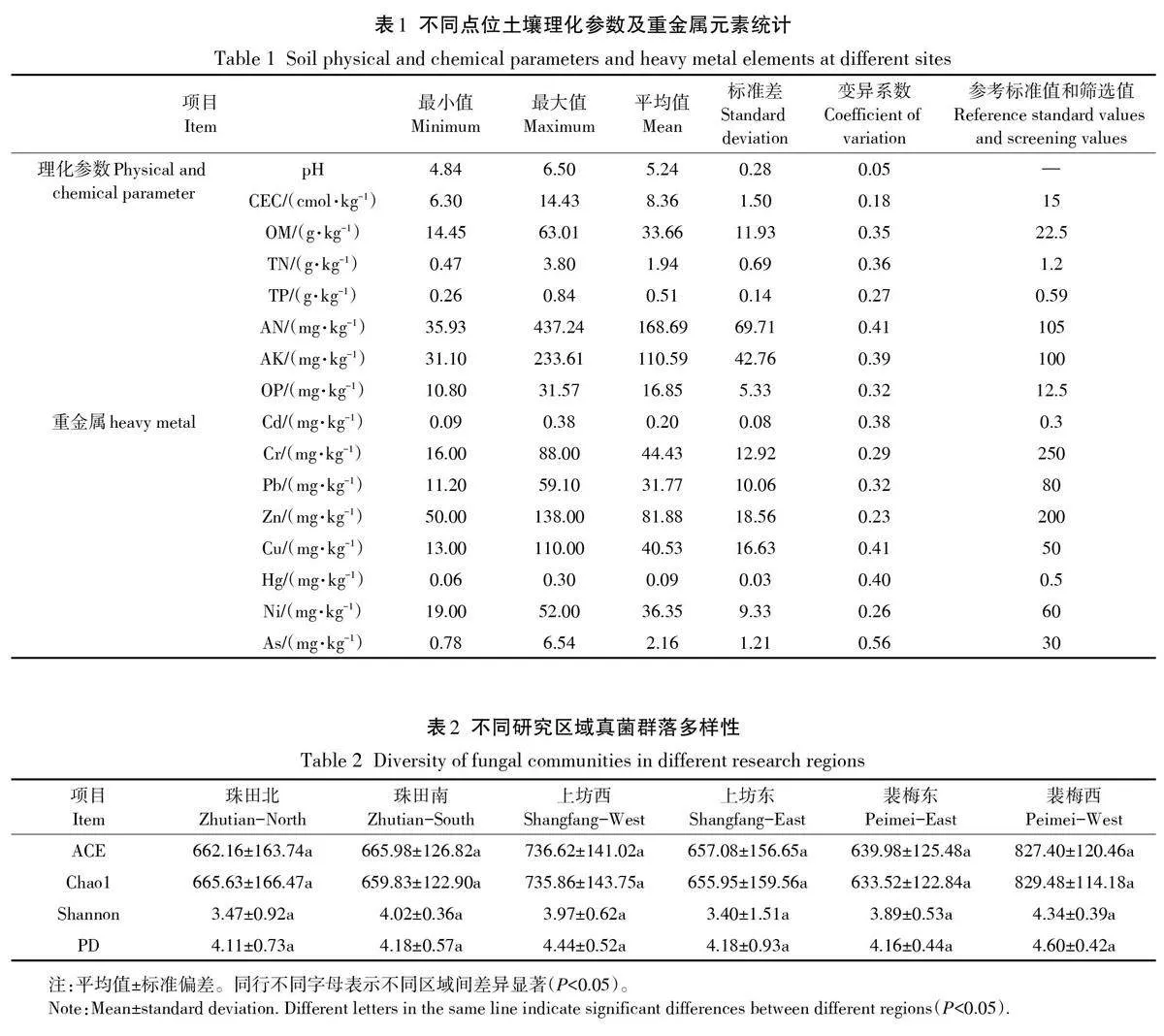
不同點位土壤理化參數及重金屬元素統計結果如表1所示。結果顯示,萬年縣流域稻田均為酸性土壤,pH介于4.84和6.50之間,平均值為5.24。由變異系數可知,不同點位稻田土壤理化指標AN和重金屬Cu、Hg、As含量差異較大,變異系數均大于0.4。依據《南方地區耕地土壤肥力診斷與評價》(NY/T 1749—2009)對測定的土壤理化進行分析,其中所有點位的CEC 均低于土壤肥力參考標準值(15 cmol·kg-1),而OM、TN、AN、AK、OP含量平均值均高于參考標準值。同時,根據《土壤環境質量農用地土壤污染風險管控標準(試行)》(GB 15618—2018)規定的篩選值對稻田土壤重金屬含量進行評估。流域內部分區域土壤Cd和Cu的含量(0.38 mg·kg-1和110 mg·kg-1)高于篩選值(0.3 mg·kg-1和50 mg·kg-1),這表明該流域內土壤存在一定的Cd和Cu污染情況。同時,所測點位中Cr、Pb、Zn、Hg、Ni、As含量均遠低于對應的污染風險篩選值,表明這6種重金屬元素基本不存在污染情況。
2.2 土壤真菌多樣性分析
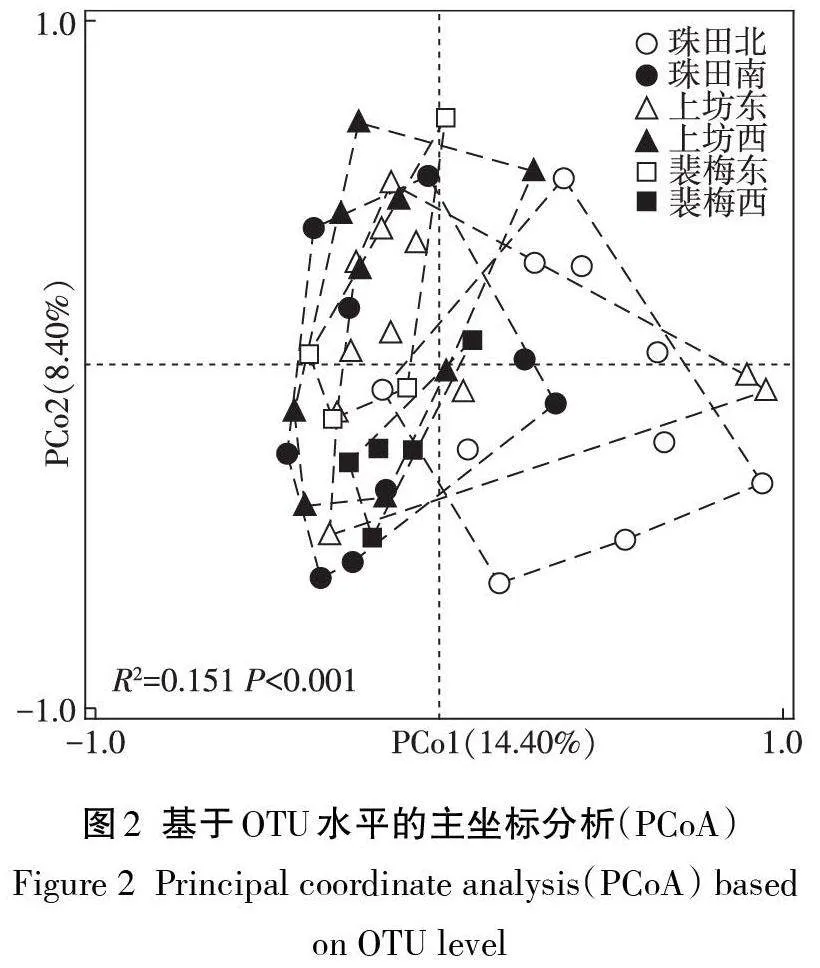
本實驗經質量控制和聚類(97%相似性)分析,共得到6 931個OTU,注釋獲得17個門、62個綱、136個目、300個科、572個屬和901個種,平均覆蓋度均高于99%。以上結果顯示測序數據合理可信,且研究區域內的真菌物種極為豐富。ACE和Chao1指數能夠很好地表征微生物群落的豐富度,Shannon和PD指數主要表達群落多樣性和譜系多樣性。由表2可知,各區域真菌群落Alpha多樣性差異性不顯著(Pgt;0.05),但仍表現出一定的變化趨勢特征。相較于其他研究區域,裴梅西真菌群落表現出最高的Alpha多樣性(包括ACE、Chao1、Shannon和PD指數),而裴梅東樣地真菌群落豐富度最低,上坊東和珠田北樣地真菌物種多樣性最低。此外發現,在上坊東、珠田北和珠田南的一些特定區域,稻田土壤表現出極低的真菌多樣性,遠低于其他點位,這些地方可能面臨微生態環境惡化風險。基于PCoA 分析發現(圖2),裴梅西樣點緊密地聚為一簇,位于研究區域樣點聚類中心位置,珠田南、上坊西和裴梅東樣點較為接近,而珠田北和上坊東樣點則較為分散。這表明了珠田南、上坊西和裴梅東3個區域稻田土壤真菌群落組成具有一定相似性,而珠田北和上坊東區域真菌群落表現出獨特的結構特征,同時珠田北和上坊東區域不同樣點稻田土壤真菌群落組成具有差異性。
2.3 土壤真菌群落組成特征
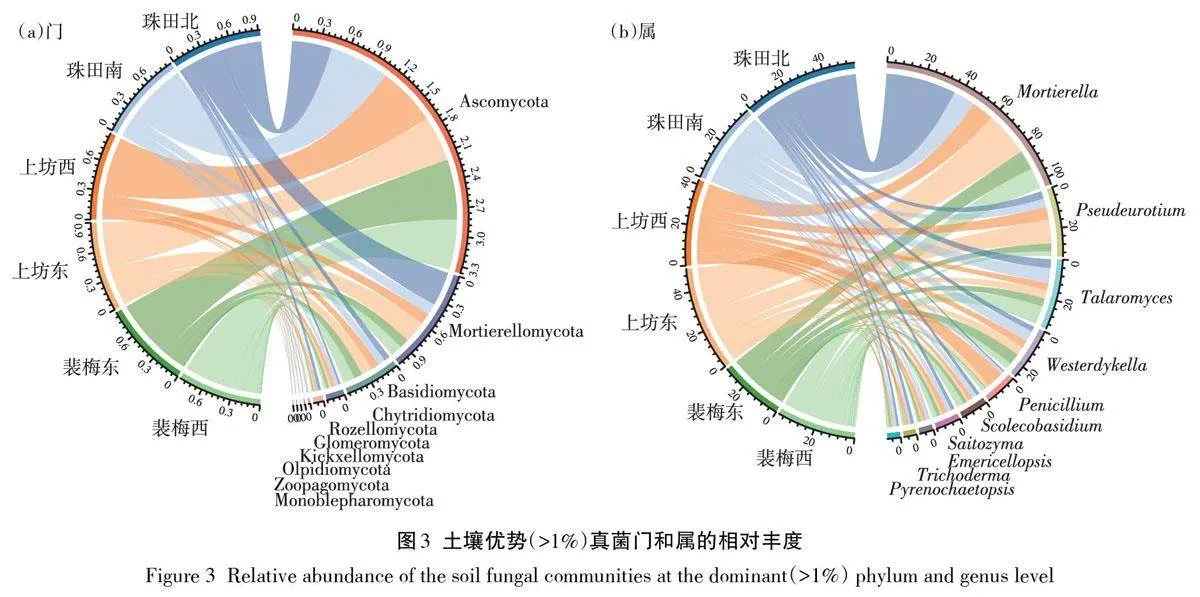
如圖3a所示,真菌優勢門(Top10)平均占比為99.86%,其中子囊菌門(Ascomycota)最為豐富,占比高達54.86%;其次是被孢霉門(Mortierellomycota)、擔子菌門(Basidiomycota)和壺菌門(Chytridiomycota),占比分別為19.55%、9.11% 和3.07%;羅茲菌門(Rozellomycota)、球囊菌門(Glomeromycota)、梳霉門(Kickxellomycota)、油壺菌門(Olpidiomycota)、捕蟲霉門(Zoopagomycota)和單毛壺菌門(Monoblepharomy?cota)的相對豐度較低,占比均低于2%。裴梅東、上坊西和珠田南真菌優勢門組成相似,而其他3個區域真菌優勢門則具有一定差異性,這與PCoA分析結果相同。相對而言,珠田北、上坊東表現出更低豐度的子囊菌門(42.57%、49.77%)和更高的被孢霉門(36.78%、23.51%),裴梅西則表現出更高豐度的擔子菌門(12.35%)。優勢屬(Top10)總占比為66.40%(圖3b),相對豐度從高到低依次是Mortierella(19.24%)、Pseudeurotium(6.33%)、Talaromyces(5.49%)、Wester?dykella(4.19%)、Penicillium(2.79%)、Scolecobasidium(2.10%)、Saitozyma(2.03%)、Emericellopsis(1.13%)、Trichoderma(1.08%)和Pyrenochaetopsis(1.04%)。基于屬水平分析發現,6個區域內真菌優勢屬的相對豐度具有較大差別。對于Mortierella 菌屬,珠田北(36.44%)和上坊東(23.39%)區域的相對豐度較高;而對于Talaromyces 菌屬,則是裴梅西(9.49%)和珠田南(7.72%)區域的相對豐度較高;此外,對于Pseudeu?rotium 菌屬和Penicillium 菌屬,上坊東(12.30%)和上坊西(8.58%)區域分別表現出更高的相對豐度,相對豐度遠高于其他區域。
2.4 影響土壤真菌群落的關鍵環境因素分析
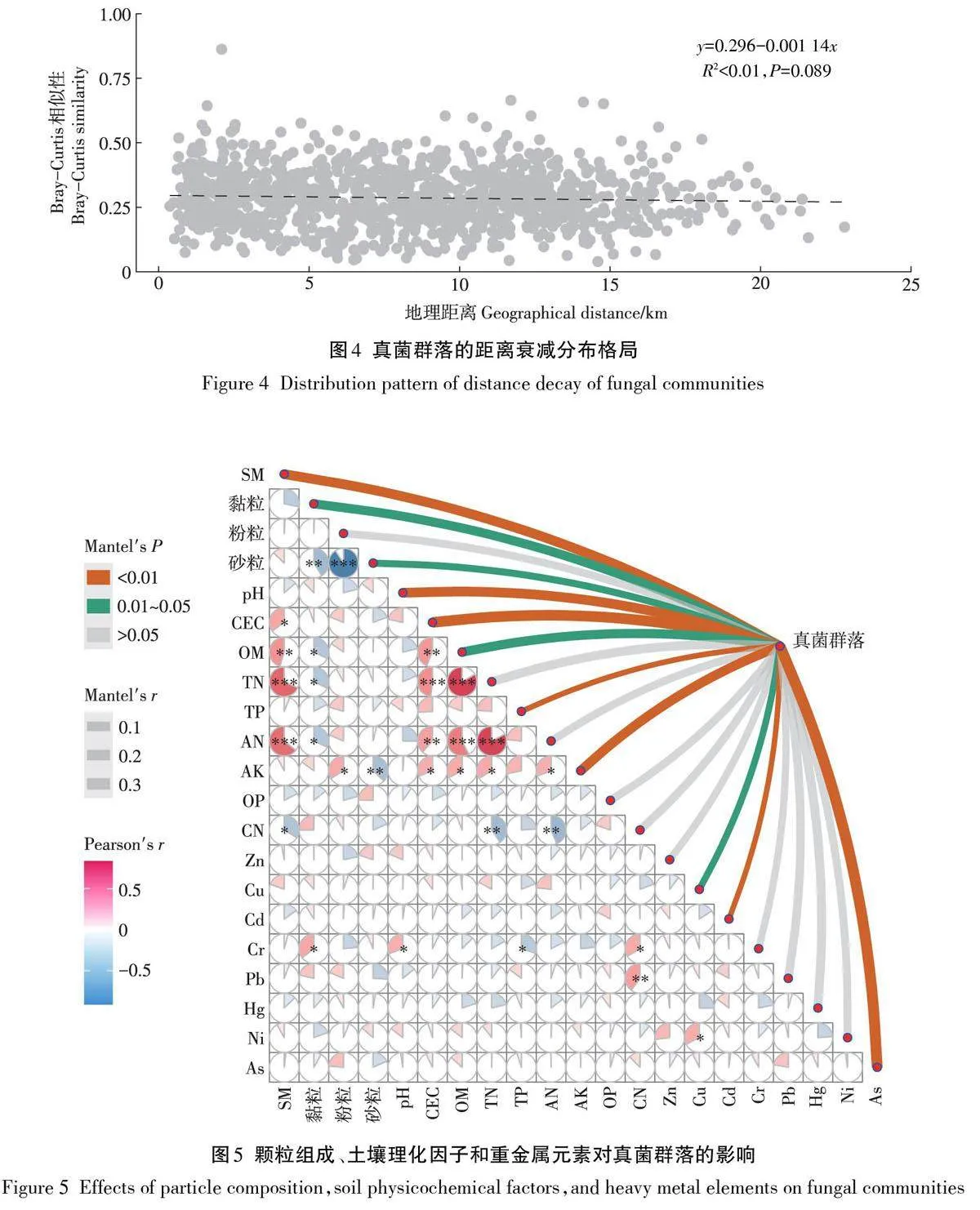
通過樣點地理坐標和樣點真菌群落相似性(Bray-Curtis距離)衰減曲線計算,明確地理距離對真菌群落結構的影響(圖4)。本研究發現樣點之間真菌群落不存在顯著的距離衰減關系(R2lt;0.01,Pgt;0.05),表明萬年縣流域稻田土壤真菌分布特征受地理距離因素影響較小,受土壤理化性質影響可能更多。Mantel分析結果(圖5)表明,樣品真菌群落與土壤理化因子SM(r=0.11)、pH(r=0.11)、CEC(r=0.20)、TP(r=-0.10)、AK(r=0.09)和重金屬元素Cd(r=-0.10)、As(r=0.23)呈極顯著相關性(Plt;0.01),同時真菌群落也會受黏粒(r=0.06)、砂粒(r=-0.06)、OM(r=0.07)和Cu(r=-0.08)的影響(Plt;0.05)。
進一步采用Spearman相關性分析環境因子與門水平和屬水平優勢菌群的關系,結果見圖6。分析結果顯示,土壤理化因子(SM、OM、TN、AN)、顆粒組成(砂粒)和重金屬元素(Cu、Cr)是影響真菌優勢門的主要環境因子。其中SM、OM、TN、AN、砂粒與壺菌門顯著正相關;OM、TN、AN、砂粒與羅茲菌門呈顯著正相關;而SM、OM、TN、AN、Cr與球囊菌門呈顯著負相關。此外,SM、TN、Cu、Ni還與油壺菌門呈顯著負相關。總體來看,壺菌門、羅茲菌門、球囊菌門、油壺菌門與環境因子有很強的耦合關系,能很好地指示環境的變化情況。對屬水平分析顯示,土壤環境因子主要影響Saitozyma 菌屬、Talaromyces 菌屬和Emericellopsis菌屬,不同的環境因子對優勢真菌屬的影響不同。其中土壤理化因子pH與Saitozyma 菌屬呈顯著負相關;CEC和OP分別與Penicillium 菌屬和Mortierella 菌屬、Scolecobasidium 菌屬呈顯著正相關。粉粒與Saitozyma菌屬呈顯著正相關;砂粒與Westerdykella 菌屬呈顯著正相關,同時與Saitozyma 菌屬呈顯著負相關。重金屬元素Cr與Talaromyces 菌屬、Saitozyma 菌屬呈顯著負相關;而Pb和Ni分別與Saitozyma 菌屬和Emericellop?sis 菌屬呈顯著正相關;Hg則與Talaromyces 菌屬呈極顯著正相關,與Emericellopsis菌屬呈顯著負相關。
3討論
3.1土壤理化性質與重金屬元素特征
本研究中萬年縣稻田土壤pH 和CEC 的空間變異性小,其原因是小流域范圍內氣候、海拔和降水量相同導致土壤pH和CEC較為相似[18-19]。同時小流域范圍內整體pH(平均值5.24)和CEC(平均值8.36)較低,CEC 已被證實與黏粒含量和pH 有關,并隨土壤pH的增大而增大,CEC較低可能導致土壤的保肥能力和緩沖性能下降[20]。OM、TN、AN、AK、OP作為衡量土壤肥力的重要指標,也是植物生長的必要養分。OM、TN、AN 等含量差異一方面與地質環境有關,另一方面可能是受不同農戶差異化田間管理措施(施肥)的影響[21-22]。施肥是最有效提高土壤養分的方式之一,但長期過量施肥不僅不能大幅提高作物產量,還會使肥料利用率下降,土壤質量退化[23]。此外,該流域內重金屬Cu、Hg、As空間變異性大,存在重金屬Cd和Cu富集情況。造成土壤重金屬污染及差異的原因,一方面是由于當地有色金屬礦的開采、冶煉和排污,另一方面與含重金屬的農藥和化肥的過量施用密切相關[24]。稻田Cd污染具有長期性和積累性,水稻根系很容易從土壤中吸收Cd并通過體內轉運到可食部分進行積累,從而對人體健康產生危害[25]。Cu雖然是植物生長必需的微量元素,但過量的Cu反而會對生物產生毒性作用[26]。因此,未來需要多關注稻田土壤重金屬污染狀況。
3.2 真菌群落多樣性及群落結構差異
真菌群落多樣性和群落結構能反映當地稻田土壤生態環境變化[27]。研究發現,萬年縣流域大部分稻田區域真菌群落組成相似(珠田南、上坊西和裴梅東)。裴梅西真菌群落表現出最高的Alpha 多樣性(包括ACE、Chao1、Shannon和PD指數),而珠田北和上坊東樣地物種多樣性較低(包括Shannon 和PD 指數),但差異性不顯著(Pgt;0.05)。以上結果表明裴梅西區域土壤可能具有更強的有機分解和轉化能力[28],而珠田北、上坊東區域土壤真菌群落組成表現出一定特異性,例如更低豐度的子囊菌門(42.57%、49.77%)和更高豐度的被孢霉門(36.78%、23.51%)。其中子囊菌門被發現可以更好地承受環境壓力,并利用更多的資源來增加其在土壤中的優勢地位[29],與其他農田生態系統相同,本研究中子囊菌在稻田土壤中占絕對優勢。而被孢霉菌門是一種產油絲狀真菌,能分解木質素和纖維素,但其菌絲會影響其他微生物類群對養分的吸收,并通過影響土壤團聚體改變土壤微生境[30],這可能是導致珠田北和上坊東區域與其他區域真菌群落組成不同的原因。此外,本研究還發現上坊東、上坊西區域優勢菌屬與其他區域具有較大差異,其中上坊東區域中Pseudeurotium菌屬和上坊西區域中Penicillium 菌屬的部分菌株為致病菌,其過多可能會威脅水稻生長和人類健康[31-32]。從真菌多樣性及群落組成綜合看,裴梅西區域稻田土壤更加健康,原因與其處于萬年縣流域上游受人為影響較小,同時保持傳統的耕作方式密不可分。而上坊東區域則需要調整和優化土壤微生態結構從而更有利于水稻的種植。
3.3驅動真菌群落變化的環境因子
土壤微生物對不同環境因子的響應存在差異,同時真菌群落結構和多樣性與土壤的理化性質和重金屬元素有著密切的聯系[33]。本研究中,小流域范圍內稻田真菌群落的變化主要由土壤理化性質(SM、pH、CEC、TP、AK)和重金屬元素(Cd、As)共同驅動,受地理距離影響小。土壤pH值被認為是區域尺度上真菌豐富度和系統發育多樣性的最強預測因子,同時土壤真菌的群落組成與土壤肥力顯著相關[34-35]。土壤微生物可以通過調節群落組成來適應不同環境,本研究中壺菌門和羅茲菌門更適應高養分的環境(高OM、TN、AN),而球囊菌門則更適應低養分土壤環境(低OM、TN、AN)。此外,磷元素同樣是植物和微生物生長的必需元素[36],捕蟲霉門、油壺菌門、被孢霉門、Mortierella 菌屬和Scolecobasidium 菌屬適應較高磷土壤環境,而子囊菌門更適應低磷土壤環境。Liu等[37]發現受重金屬Cd污染的稻田土壤常表現出更低的微生物生物量和真菌豐度,同時表現出更低的碳利用效率。本研究表明,油壺菌門對于稻田土壤中重金屬Ni、Cu具有很好的指示作用;球囊菌門、擔子菌門、Talaromyces 菌屬和Saitozyma菌屬對重金屬Cr的變化較敏感;而Saitozyma 菌屬和Emericellopsis 菌屬分別與Pb和Ni呈顯著正相關關系(Plt;0.05),表明其對相應重金屬具有一定的耐受性。總而言之,壺菌門、羅茲菌門、球囊菌門、油壺菌門、Saitozyma菌屬、Talaromy?ces菌屬和Emericellopsis菌屬對土壤養分情況及土壤重金屬污染有很好的指示作用,可以反映一定的土壤健康狀況,對水稻的種植具有重要意義。
4結論
(1)江西小流域范圍內土壤理化性質接近,僅水解氮和重金屬Cu、Hg、As變異系數較大,同時部分點位存在Cd和Cu重金屬污染情況。未來應多關注周邊礦山開采與重金屬污染對稻田土壤的影響。
(2)江西小流域范圍內稻田土壤真菌群落組成具有差異性。相較而言,裴梅西處于珠溪河上游地區且保持傳統耕作方式,表現出最高豐度的擔子菌門(12.35%)和Talaromyces菌屬(9.49%);珠田北、上坊東則處于珠溪河中下游地區受人為活動影響大,表現出更低豐度的子囊菌門(42.57%、49.77%)和更高豐度的被孢霉門(36.78%、23.51%)。
(3)土壤理化指標(含水量、pH、陽離子交換量、總磷、速效鉀)和重金屬元素(Cd、As)是影響萬年縣稻田土壤真菌類群變化的關鍵環境因子。壺菌門、羅茲菌門、球囊菌門、油壺菌門、Saitozyma 菌屬、Tal?aromyces菌屬和Emericellopsis 菌屬可以很好地反映出稻田土壤肥力和重金屬污染情況,有利于參考并及時采取相應措施保障稻田土壤健康。
