豬流行性腹瀉病毒SCYA2204 株的基因克隆及測序分析
2024-01-21 09:31:02孔晶晶田珺劼任玉鵬陳弟詩
四川畜牧獸醫 2024年1期
孔晶晶,田珺劼,任玉鵬*,陳弟詩,包 昕
(1.西南民族大學畜牧獸醫學院,四川 成都 610041;2.四川省動物疫病預防控制中心,四川 成都 610041)
豬流行性腹瀉(PED)由冠狀病毒科Alpha 冠狀病毒屬的豬流行性腹瀉病毒(PEDV)引起,PEDV 對各日齡豬都具有易感性,對2 周齡內仔豬的致死率可達100%[1]。PEDV 為單股正鏈RNA 病毒,全基因組序列大小約為28 kb,包含5′端帽子結構,3′端poly(A)尾及7 個開放閱讀框(ORF),共編碼纖突蛋白(S蛋白)、包膜蛋白(E蛋白)、膜蛋白(M蛋白)和核衣殼蛋白(N蛋白)等四種結構蛋白,以及ORF1ab 復制酶多聚蛋白和ORF3蛋白等非結構蛋白[2]。S蛋白是PEDV重要的毒力蛋白,它所構建的系統進化樹可將PEDV劃分為G1和G2兩個型,其中G2型包含G2a、G2b和G2c 亞型[3],我國目前流行的毒株主要是G2a、G2b 亞型。監測S 基因的遺傳變異對PED 的防控施策具有重要意義[4]。此外,PEDV的ORF3蛋白是唯一的非結構輔助蛋白。在PEDV連續傳代弱化毒株中,ORF3 基因存在17 個連續氨基酸的穩定缺失,可能與其毒力弱化有關[5]。E 蛋白主要調控病毒組裝、胞內運輸和病毒釋放等過程,還能引起內質網產生應激,并促使宿主細胞產生白細胞介素-8(IL-8)、B-細胞淋巴瘤/白血病-2基因(Bcl-2),造成細胞損傷和誘導細胞凋亡[6]。M 蛋白位于整個細胞質中,通過與S 蛋白和N 蛋白相互作用在病毒核衣殼的組裝過程中發揮重要作用,對細胞增殖和病毒復制具有重要影響。N蛋白是一種高度保守的磷蛋白[7],是PEDV核衣殼的基礎結構,在病毒復制、組裝過程中發揮重要作用。
本研究對四川省雅安地區流行的一株PEDV SCYA2204的S基因、ORF3基因、E基因、M基因和N基因進行基因克隆及測序,分析病毒基因核苷酸序列,并推導氨基酸序列的變異性及遺傳進化特征,以期為PEDV 的有效防控提供理論依據。
1 材料與方法
1.1 主要試劑 Trizol、反轉錄試劑盒、Taq DNA聚合酶、pMD19-T載體、大腸桿菌DH5α感受態細胞、DL2000 DNA Marker、膠回收試劑盒。
1.2 臨床樣本來源與處理 臨床樣本于2022年4 月采自四川省雅安市某規模化豬場,用無菌棉簽采集疑似PEDV 感染豬的肛門棉拭子,低溫保存于DMEM 基礎培養基中。將樣本用適量PBS 稀釋后反復凍融3 次,在高速冷凍離心機上以10 000 r/min 離心15 min,取上清液。樣本由西南民族大學動物醫學實驗室進行RT-PCR 檢測,確定為PEDV陽性樣本,并凍存于-80 ℃冰箱。
1.3 引物設計 PEDV S 基因、ORF3 基因和M 基因的引物均參照文獻[8-9]合成,其中對S基因采用分段重疊法,分4 段(S1、S2、S3、S4)進行測序。E 基因、N 基因的引物以GenBank 中CV777、AJ1102毒株的基因組序列為參考,運用引物設計軟件Primer Primier 5.0設計。所有引物均由上海生工生物工程技術服務有限公司合成(表1)。
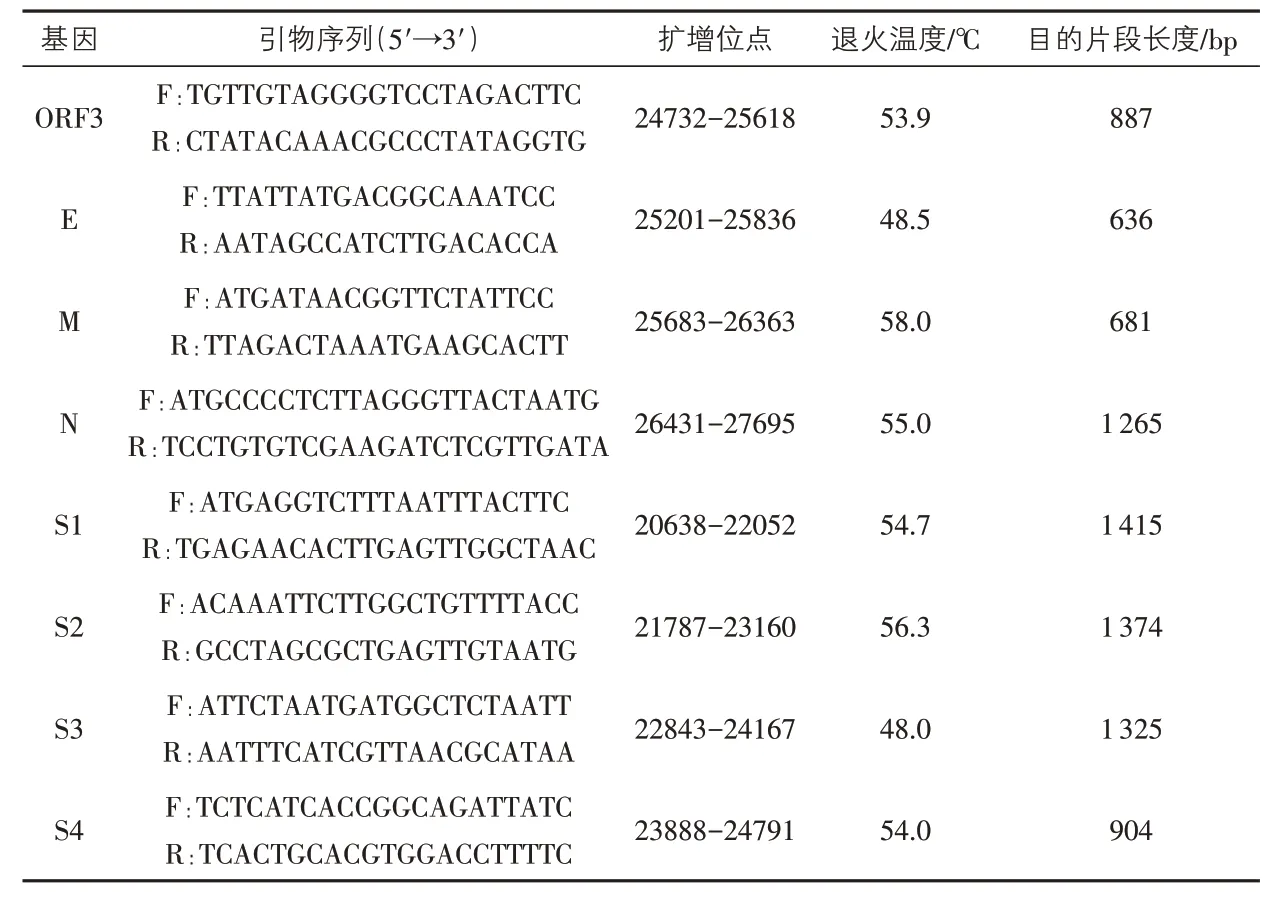
表1 PEDV擴增引物信息
1.4 RNA 提取與反轉錄 使用Trizol 法提取上述樣品的總RNA。參照反轉錄試劑盒說明書將RNA 反轉錄為cDNA,所得cDNA 于-20 ℃凍存備用。
1.5 目的基因的PCR 擴增 25 μL PCR 擴增加樣體系:ddH2O 10.5μL,2×PCR MasterMix 12.5μL,上、下游引物各0.5 μL,cDNA 1 μL。反應條件:94 ℃預變性1 min;94 ℃變性30 s,退火30 s(退火溫度參照表1設置),72 ℃延伸50 s,共35個循環;72 ℃延伸10 min。取5 μL PCR 產物進行1.5%瓊脂糖凝膠電泳。
1.6 基因的克隆及測序 根據Omega 生物技術公司膠回收試劑盒說明書進行回收純化。參照pMDTM19-T Vector Cloning Kit說明書進行連接,并轉化大腸桿菌DH5α感受態細胞,在含氨芐青霉素的牛肉膏蛋白胨培養基(100 μg/mL Amp LB)瓊脂上篩選陽性克隆,送樣至生工生物工程(上海)股份有限公司測序。
1.7 基因序列的生物信息學分析 選取62株國內外PEDV毒株,其中28株國外毒株,包括2株常用疫苗毒株AJ1102、CV777(GenBank登錄號分別為JX188454、AF353511);34株國內毒株,包括14株往年四川地區流行的毒株。用SeqMan 軟件對基因片段進行拼接,將各基因序列在NCBI 中比對,在MegAlign軟件中用Clustal W法計算各基因的核苷酸同源性;用MEGA 11 軟件中Neighbor-Joining法構建系統進化樹;使用Alignment Report分析各基因的變異性及S基因推導的氨基酸的變異性;利用RDP4軟件中的7種不同程序方法對S基因序列的重組信號進行檢測。
2 結果
2.1 PEDV 基因的克隆測序 對PEDV S 基因、ORF3基因、E基因、M基因和N基因進行擴增,獲得與預期大小相符的條帶(圖1)。將PCR產物克隆測序,對S 基因重疊的區域進行拼接,得到PEDV S 基因的總長度為4 161 bp,ORF3 基因、E基因、M 基因和N 基因序列的長度分別為675、231、681、1 326 bp。將此毒株命名為SCYA2204。
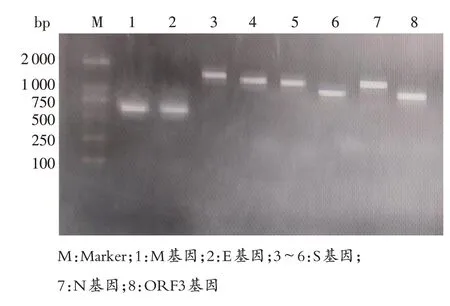
圖1 PEDV基因各片段的RT-PCR擴增產物
2.2 PEDV 基因的同源性分析 與參考毒株相比,SCYA2204 S基因的核苷酸同源性為92.5%~99.2%,其中與四川毒株CH/SCXH/2018的核苷酸同源性最高,為99.2%;與經典疫苗株CV777 S基因的核苷酸同源性為93.2%,與AJ1102 S基因的核苷酸同源性為96.8%,而與四川近年流行毒株(SCDY/05/2020、SCDZ-1/2020、SCXH/2018、SCYB/2018、SC/CY/2022、SC2021 等)的同源性高達98.1%~99.2%。另外,ORF3 基因、E 基因、M基因和N 基因的核苷酸同源性分別為96.0%~100%、97.0%~100%、97.4%~99.7%、95.2%~99.9%。其中,SCYA2204 與美國地區IA1、NPLPEDV/2013和中國地區LYG等毒株ORF3基因的核苷酸同源性最高;與中國地區SC2021、CH/JXJA/2017 和國外地區IA1、ON-018 等毒株E 基因的核苷酸同源性最高,均為100%;與SC2021、HK2021 M基因的核苷酸同源性最高,為99.7%;與HN2021、SD2020、NH-TA2020 N 基因的核苷酸同源性最高達99.9%(表2)。
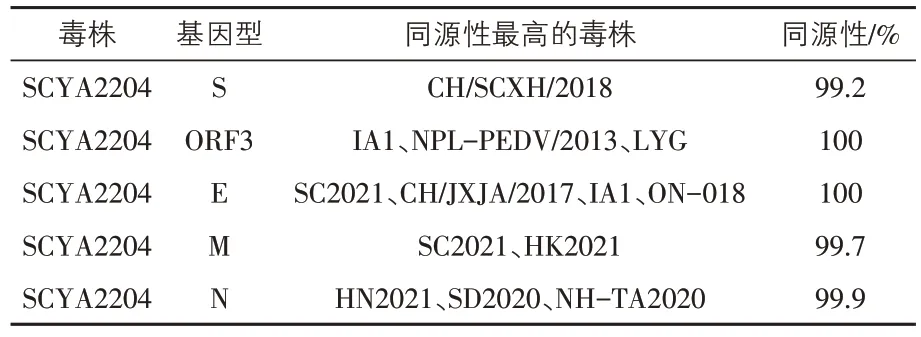
表2 PEDV SCYA2204株各基因同源性分析
2.2 PEDV基因的遺傳進化樹分析 由S基因構建的遺傳進化樹(圖2)將所有PEDV分為G1、G2a和G2b 三個亞型,SCYA2204 與28 株PEDV 國內外流行毒株同屬于G2b亞型。其中SCYA2204與四川往年流行毒株CH/SCYB-1/2018、CH/SCNJ-1/2020、CH/SCXH/2018、SWUN-C6-CH-SCYA-2019、SWUN-Y1-CH-SCCQ-2019 聚成一支,親緣關系密切,與SC2021的親緣關系相對較遠。此外,常用疫苗株AJ1102 屬于G2b 亞型,CV777 和DR3 屬于G1a 亞型,與SCYA2204 的親緣關系都比較遠。

圖2 S基因的遺傳進化樹分析
由ORF3 基因構建的系統發育樹(圖3A)可見,SCYA2204 與CH/SXWS/2018、PEDV-LYG 的親緣關系較近,與四川往年流行毒株SC2021、CH/SCAZ10/2017 的親緣關系均相對較遠。此外,由于CV777、AJ1102 和DR13 為弱毒疫苗株,其ORF3 基因均存在50 bp 堿基缺失,因此,三個毒株的ORF3基因序列與其他毒株的均不在同一分支。由E基因遺傳進化樹(圖3B)可見,河南毒株HN2021、山東毒株SDTA13-2020、四川毒株SC2021 等都與SCYA2204 的親緣關系極其密切。在M基因遺傳進化樹(圖3C)中,四川往年流行毒株SC2021、國內流行毒株HK2021、江西流行毒株JX2020 與SCYA2204 聚成一支,親緣關系極其密切。在N基因遺傳進化樹(圖3D)中,河南毒株HN2021、國內毒株TRS2021與SCYA2204的親緣關系密切,三株山東毒株NH-TA2020、SD2020、SDTA13-2020 與SCYA2204 的親緣關系也較為密切,而四川往年流行毒株SC2021 與SCYA2204的親緣關系則相對較遠。E基因、M基因和N 基因的系統發育樹均提示經典疫苗株CV777、AJ1102與SCYA2204的親緣關系較遠。
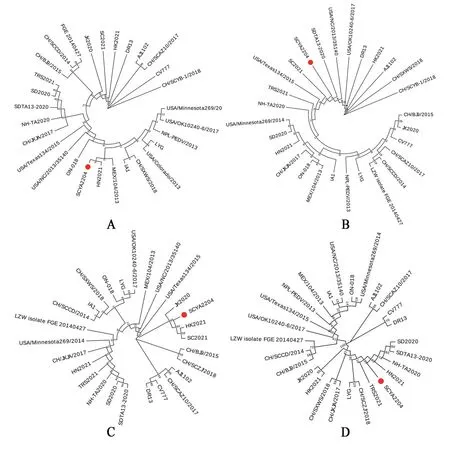
圖3 ORF3、E、M、N基因的遺傳進化樹分析
2.3 PEDV S 蛋白的變異性分析 將SCYA2204毒株S 基因推導的氨基酸序列與常用疫苗毒株CV777、AJ1102 及四川往年流行毒株SC2021、SCDZ-1/2020、SCCY2022、CH/SCDY/05/2020作比較,共有10 處氨基酸(AA)突變,其中存在3 處連續AA 突變,分別為PC700/701LW、QA706/707HY、DDIV712/713/714/715HNTP。與CV777 相比,共有83處AA 突變,其中17 處為連續AA 突變:TP2/3KS、GSTI27/28/29/30SANT、SMNS55/56/57/58IGEN、GTGIE68/69/70/71/72AGQHP、DS86/87RG、DN130/131SI、VN148/149AD、LQD157/158/159HMS、KNI161/162/163EHS、KRS200/201/202SGG、EP229/230QL、DS246/247EP、PC700/701LW、QA706/707HY、DDIV712/713/714/715HNTP、GD1177/1178DE、TY1197/1198NH;在59~63 位存在4 個氨基酸插入(QGVN),在134位存在一個氨基酸插入(N),160位存在一個氨基酸的缺失(G)。與AJ1102 株比較,共有34 處AA 突變,其中3 處為連續突變:PC696/697LW、QA702/703HY、DDIV708/709/710/711HNTP,30處為點突變,1處在1 198位插入(H)。
S 基因編碼的S 蛋白能夠誘導機體產生中和抗體。研究顯示[10],S 蛋白含有的抗原中和表位包括COE(499~638 AA)、SS2(748~755 AA)、SS6(764~771 AA)、2C10(1 368~1 374 AA)。與CV777 比較(圖4),SCYA2204 的中和表位共有7處AA 突變:A521S、F540L、T553S、G598S、Q637E、L768S、D770S,其中COE 區有5處突變:A521S、F540L、T553S、G598S、Q637E,SS6區有2 處突變:L768S、D770S。與AJ1102比較,SCYA2204 的中和表位共有3 處AA 突變:T500I、A521S、F540L,其中COE區A521S、F540L兩處突變為SCYA2204 與AJ1102、CV777 共有的AA 突變。SCYA2204的SS 區 與CV777 和AJ1102 比 較,均無突變。

圖4 S蛋白中和表位的比對分析
2.4 PEDV S 基因的重組分析 運用RDP4軟件中的7個不同程序方法(RDP、MaxChi、GENECONV、BootScan、Chimaera、SiScan 和3Seq)對S 基因序列的重組信號進行檢測,結果發現:SCYA2204與41株參考毒株的S基因之間存在1個潛在重組事件,7種重組檢測方法中有6種皆呈陽性,發生概率較高。主要親本為G2b亞型的CH/SCXH/2018,次要親本基于G1 亞型的LZC株推導,重組區域在S基因的2108~2237 bp。
3 討論
目前PEDV 仍廣泛流行于全國各地。Zhang Hao 等[11]對2011~2021 年華中、華南和華東地區共計149 869 例腹瀉豬組織樣本進行檢測,結果表明PEDV 仍然是過去11 年引起國內豬腹瀉病的最主要病原,流行率為61.8%。雖然當前幾種商品化的弱毒疫苗和滅活疫苗已被廣泛用于PED 的防控,但由于PEDV 基因組的高度變異性導致其基因亞型不斷增加,基因重組事件頻發,造成毒株間的交叉保護效果差,如:研究已證實基于G1 型毒株研制的疫苗(CV777)已無法對G2a和G2b 亞型的PEDV提供良好的保護[12];且不同G2 亞型毒株間的交叉保護效果也有差異[13]。據調查顯示,PEDV 在國內不同地區的優勢基因型有較大差異[14];同時,當前四川地區流行的PEDV 毒株的變異性和基因多樣性在不斷增加。本次四川雅安地區流行的SCYA2204株屬于G2b 亞型,其S 基因發生了較大的變異。與CV777 比較,SCYA2204 有17 處連續AA 突變,且SCYA2204 株S 蛋白存在3 處獨特的連續AA 突變,與國內近幾年流行毒株(SC2021、SCDZ-1/2020、SCCY2022、CH/SCDY/05/2020 等)的S 蛋白氨基酸序列有明顯差異。研究表明,PEDV S 基因包括多個病毒主要的中和表位和受體結合域,它們與病毒的抗原性和吸附入侵密切相關[10],在病毒—宿主細胞膜融合的信號轉導過程中起著重要作用[15],其融合活性也決定了病毒的組織嗜性和細胞感染能力。本研究中,將SCYA2204 株與CV777株的4個當前已鑒定的S蛋白中和表位比較,SCYA2204 株中和表位共有7 處AA 突變;與AJ1102 比較,SCYA2204 株的中和表位有3 處AA突變。其中,COE區的A521S、F540L兩處突變為SCYA2204 與AJ1102、CV777 共有的AA 突變。SCYA2204 株的氨基酸變化,尤其是中和表位區內的變異,可能導致其抗原性發生改變,影響傳統疫苗株的保護效力。
本研究分析了SCYA2204 S、ORF3、E、M、N基因的核苷酸同源性和遺傳進化樹,發現SCYA2204株與四川往年流行毒株(如CH/SCYB-1/2018、CH/SCNJ-1/2020、CH/SCXH/2018、SCDY/05/2020、SC2021)的親緣關系密切,而與常用疫苗株AJ1102 和經典毒株CV777 的親緣關系都較遠;E基因、M基因和N基因的系統發育樹均提示經典疫苗株CV777、AJ1102 與SCYA2204 的親緣關系較遠。進一步說明基于傳統毒株的疫苗對于現在四川流行毒株的保護效力可能會有所下降。已有研究發現PEDV S 基因易發生重組事件,重組可能使高毒力PEDV 毒株在豬群中持續存在和傳播[16-17]。本研究發現PEDV SCYA2204與25株參考毒株的S基因之間存在1個潛在重組事件,6 種重組檢測方法皆呈陽性,說明PEDV 各毒株間發生重組現象的概率較高。
據調查,本次PED 發病豬場已免疫G2b亞型疫苗,但未能較好地交叉保護引發本次疫情的SCYA2204 毒株(G2b 亞型)。提示未來還需進一步篩選新型有效的PEDV 候選疫苗毒株,這對有效防控PED疫情具有重要意義。
4 結論
本研究獲得了PEDV SCYA2204 株的S、ORF3、E、M 基因和N 基因的核苷酸序列,并進行了變異分析和遺傳進化分析。結果得出:SCYA2204的S 基因與經典株CV777 的核苷酸同源性為93.2%,與AJ1102毒株的核苷酸同源性為96.8%,而與四川近年流行毒株的同源性高達98.1%~99.2%。這與本研究所檢測的PEDV 毒株S 基因的核苷酸同源性與系統發育進化樹分析結果一致。與SCYA2204 株ORF3、E、M、N 基因的核苷酸同源性最高的毒株大部分來自不同地區,僅有少數來自四川地區。PEDV 毒株分為G1和G2兩個型,G1 又可以進一步分為2 個亞型,即G1a、G1b,本試驗毒株SCYA2204經基因序列比對與進化樹分析,被劃分為G2b 亞型。和常用疫苗毒株AJ1102 相比,SCYA2204 株的S 基因具有34 個氨基酸位點變化,且發生了重組事件。
