POU1F1基因SNP位點(diǎn)與尼羅羅非魚體質(zhì)量和形態(tài)性狀的相關(guān)性
2024-01-26 08:19:14高風(fēng)英佟延南曹建萌劉志剛衣萌萌可小麗盧邁新
廣東海洋大學(xué)學(xué)報(bào) 2024年1期
高風(fēng)英,佟延南,曹建萌,劉志剛,王 淼,衣萌萌,可小麗,盧邁新,朱 海
(1.中國(guó)水產(chǎn)科學(xué)研究院珠江水產(chǎn)研究所/農(nóng)業(yè)部熱帶亞熱帶水產(chǎn)資源利用與養(yǎng)殖重點(diǎn)實(shí)驗(yàn)室,廣東 廣州 510380;2.海南省海洋與漁業(yè)科學(xué)院,海南 海口 570206)
垂體特異性正轉(zhuǎn)錄因子1(POU1F1)(亦稱PIT-1 或GHF-1)是一種在垂體前葉中表達(dá)的組織特異性轉(zhuǎn)錄因子[1,2],在生長(zhǎng)激素(GH)和催乳素(PRL)基因的轉(zhuǎn)錄調(diào)節(jié)中有一定作用[3,4],還參與激活促甲狀腺激素b 亞基(TSHb)、POU1F1基因[5-6]和生長(zhǎng)激素釋放激素受體基因[7],對(duì)促體細(xì)胞、促乳細(xì)胞以及促甲狀腺細(xì)胞的分化、繁殖和生存亦至關(guān)重要[5],是有育種潛力的候選基因,值得在羅非魚中深入研究。此外,由于POU1F1的下游靶基因和調(diào)控細(xì)胞在物理生長(zhǎng)中起著直接作用,POU1F1中的一些SNP 與畜禽的生長(zhǎng)性狀密切相關(guān)[8-13]。在水產(chǎn)領(lǐng)域,僅杜芳芳等[14]對(duì)大口黑鱸(Micropterus salmoides)POU1F1基因多態(tài)性與生長(zhǎng)相關(guān)性狀關(guān)聯(lián)性進(jìn)行了研究,獲得了生長(zhǎng)相關(guān)基因型。
目前,已廣泛實(shí)施分子標(biāo)記輔助育種(Marker Assisted Selection,MAS),以改善水產(chǎn)動(dòng)物生產(chǎn)性能[15-16]。羅非魚是我國(guó)重要的淡水養(yǎng)殖魚類之一,以尼羅羅非魚為主。據(jù)《2022 中國(guó)漁業(yè)統(tǒng)計(jì)年鑒》,2020 和2021 年我國(guó)羅非魚產(chǎn)量超過世界總產(chǎn)量的30%,在國(guó)民經(jīng)濟(jì)中占有重要地位,羅非魚的選育研究有重要意義。羅非魚的選育目標(biāo)主要是生長(zhǎng)速度。尼羅羅非魚引入我國(guó)后,經(jīng)多年的選育,不同選育群體間因選育技術(shù)不同,生長(zhǎng)速度不同。為繼續(xù)選育尼羅羅非魚,篩選生長(zhǎng)相關(guān)分子標(biāo)記以輔助選育非常必要。POU1F1基因多態(tài)性與家畜的生長(zhǎng)和肉質(zhì)性狀相關(guān),為探索羅非魚中是否存在與體質(zhì)量以及全長(zhǎng)、體長(zhǎng)、頭長(zhǎng)、體高、體寬等形態(tài)性狀相關(guān)的POU1F1多態(tài)位點(diǎn),本研究選取POU1F1基因,在尼羅羅非魚高要親代群體中篩選SNP,并與生長(zhǎng)數(shù)據(jù)相結(jié)合,篩選與體質(zhì)量和形態(tài)性狀相關(guān)的SNP,并將這些SNP在高要子代群體、番禺群體以及海南群體中進(jìn)行普適性驗(yàn)證,旨在為羅非魚以生長(zhǎng)性狀為目標(biāo)的選育提供有效分子標(biāo)記。
1 材料與方法
1.1 材料
篩選SNP 位點(diǎn)樣品:尼羅羅非魚(Oreochromis niloticus)吉富品系,取自珠江水產(chǎn)研究所水產(chǎn)良種基地,40 尾魚的鰭條樣本,置20 ℃無(wú)水乙醇中保存。
體質(zhì)量和形態(tài)性狀關(guān)聯(lián)分析樣品:尼羅羅非魚吉富品系高要親代及子代群體取自珠江水產(chǎn)研究所水產(chǎn)良種基地,“廣特超”新吉富羅非魚(番禺群體)取自廣東羅非魚良種場(chǎng),尼羅羅非魚海南群體取自海南省水產(chǎn)科學(xué)院。高要親代群體為2020 年的秋苗,取1 000 尾同塘養(yǎng)殖越冬至次年7 月;高要子代群體為2021 年用傳統(tǒng)土塘繁殖的春苗,取200尾同水泥池養(yǎng)殖3個(gè)月(經(jīng)激素處理,全雄);番禺群體為2021 年春苗(經(jīng)激素處理,全雄),取200 尾,同水泥池養(yǎng)殖3個(gè)月;海南群體為2021年春苗1萬(wàn)尾,放至1/3 hm2的土塘中養(yǎng)殖3 個(gè)月(未經(jīng)激素處理的正常雌雄群體)。取高要群體162 尾,番禺群體175尾,分別測(cè)量體質(zhì)量、全長(zhǎng)、體長(zhǎng)、頭長(zhǎng)、提高、體寬,海南群體300尾,測(cè)量體質(zhì)量。
1.2 方法
1.2.1 基因組DNA 的提取 采用磁珠法基因組DNA 提取試劑盒(NanoMagBio)提取基因組DNA,具體方法步驟參照試劑盒說(shuō)明書。用瓊脂糖凝膠電泳檢測(cè)基因組DNA質(zhì)量。
1.2.2POU1F1基因SNP 位點(diǎn)的篩查 根據(jù)POU1F1基因(GenBank accession No.XM_019352661,NC_031987)3?區(qū)域序列,利用Primer Primer 5.0 軟件設(shè)計(jì)2 對(duì)上下游引物:POU1F1-3end_1-F(15631-15655)(CGTACAAAATATGCCATTTACCAAG)、POU1F1-3end_1-R(16976-16995)(ACTTTATCAAGGCAGCGACG),POU1F1-3end_2-F(16749-16772)(GCTTTAACCAA ATGAATGTAGCTG)、POU1F1-3end_2-R(18139-18160)(TTTCAGTCTCCACTTTATCCCC),以及測(cè)序引物POU1F1-3end_2-seqf1(GAGTGGTTTTCACTGTGAC)。以上述高要親代群體中40個(gè)個(gè)體的DNA為模板,進(jìn)行PCR 擴(kuò)增。反應(yīng)體系(30μL):2×Taq PCR Master Mix 15μL,基因組DNA 1μL(約20 ng),10 pmol/μL上下游引物各1μL,ddH2O 12μL。反應(yīng)條件:95 ℃5 min;95 ℃30 s、55 ℃30 s、72 ℃45 s,35 個(gè)循環(huán);72 ℃5 min,16 ℃1 min,PCR 產(chǎn)物4 ℃下保存。條帶符合理論目的大小的PCR 產(chǎn)物送Sanger 測(cè)序,測(cè)序平臺(tái)為ABI 公司的3730xl DNA Analyzer 測(cè)序儀。2 對(duì)引物分別做雙向測(cè)序后正反向拼接,且兩段檢測(cè)區(qū)域有重疊,以保證2 對(duì)引物測(cè)通全長(zhǎng)。由于尾端有POLY 結(jié)構(gòu),因此多設(shè)計(jì)一條加測(cè)引物pou1f1-3end_2-seqf1,以避開POLY 結(jié)構(gòu)。將測(cè)序良好的全長(zhǎng)序列在SeqMan 軟件中進(jìn)行多序列比對(duì),篩查POU1F1基因中的SNP位點(diǎn)。
1.2.3 POU1F1 基因SNP 位點(diǎn)的分型分析 采用
1.2.2 中PCR 產(chǎn)物測(cè)序法分析SNP 位點(diǎn)分型。反應(yīng)體系和反應(yīng)條件同1.2.2.。共分析3 個(gè)群體:首先分析高要親代群體213 尾的SNP 位點(diǎn)分型,并分析SNP 位點(diǎn)基因型與體質(zhì)量和形態(tài)性狀的相關(guān)性,將獲得的與體質(zhì)量和形態(tài)性狀相關(guān)的SNP 位點(diǎn)在高要子代群體(162 尾)以及番禺群體(175 尾)中進(jìn)行分型分析,分析SNP 位點(diǎn)基因型(下文基因型中“I”代表TC 插入、CC 插入、TTTG 插入、C 插入、CTC 插入、T 插入;“D”代表缺失)與體質(zhì)量和形態(tài)性狀的相關(guān)性,驗(yàn)證親子代群體間體質(zhì)量和形態(tài)性狀相關(guān)SNP 位點(diǎn)的可重復(fù)性,以及不同群體間的可重復(fù)性。
1.2.4POU1F1基因SNP 位點(diǎn)的分型驗(yàn)證 將與高要親代群體、高要子代群體以及番禺群體體質(zhì)量和形態(tài)性狀相關(guān)的位點(diǎn),在海南群體中進(jìn)行擴(kuò)增,并用SeqMan 軟件統(tǒng)計(jì)基因型,分析SNP 位點(diǎn)基因型與體質(zhì)量和形態(tài)性狀的相關(guān)性,從而進(jìn)一步驗(yàn)證所獲得的體質(zhì)量和形態(tài)性狀相關(guān)SNP 位點(diǎn)的可重復(fù)性。
1.2.5 遺傳信息統(tǒng)計(jì) 利用PIC-CALC 0.6 分析SNP位點(diǎn)的多態(tài)信息含量(polymorphism information content,PIC)。利用Popgene 3.2 進(jìn)行哈迪-溫伯格平衡(Hardy-Weinberg equilibrium,HWE)狀態(tài)的卡方檢驗(yàn),并計(jì)算觀測(cè)雜合度(Ho)、期望雜合度(He))、有效等位基因數(shù)(Ne)、基因頻率及等位基因頻率。采用HaploView 4.2 軟件對(duì)SNP 位點(diǎn)進(jìn)行連鎖不平衡分析和單倍塊構(gòu)建。利用單倍型手動(dòng)分析出雙倍型。
1.3 數(shù)據(jù)的關(guān)聯(lián)分析
采用SPSS17 軟件一般線性模型中的多元方差分析模塊分析SNP 位點(diǎn)不同基因型與全長(zhǎng)、體長(zhǎng)、頭長(zhǎng)、體高、體寬、體質(zhì)量等6個(gè)生長(zhǎng)性狀的相關(guān)性。數(shù)據(jù)以平均值±標(biāo)準(zhǔn)差形式表達(dá),差異顯著性水平α=0.05。
2 結(jié)果分析
2.1 POU1F1基因部分序列SNP位點(diǎn)
將PCR 擴(kuò)增獲得的2 341 bp片段(登錄號(hào):NC_031987)進(jìn)行序列比對(duì),獲得28個(gè)SNP位點(diǎn)(表1)。
2.2 各群體中POU1F1基因SNP位點(diǎn)的遺傳多樣性
2.2.1 高要親代群體 表2 可見:高要親代群體POU1F1基因28 個(gè)SNP 位點(diǎn)的多樣性較為豐富,有效等位基因數(shù)(Ne)、觀測(cè)雜合度(Ho)、期望雜合度(He)平均值分別為1.359 3、0.240 7、0.156 6;多態(tài)信息含量(PIC)平均為0.298,有6 個(gè)位點(diǎn)低度多態(tài)(PIC <0.25),20 個(gè)位點(diǎn)中度多態(tài)(0.25 <PIC <0.50),2 個(gè)位點(diǎn)高度多態(tài)。位點(diǎn)S1(A-198G)、S2(C-201T)、S4(A-469T)、S6(A-881G)、S7(A-888G) 在 群體中符合Hardy-Weinberg 平衡定律(下稱“H-W 平衡”),其余23個(gè)位點(diǎn)均偏離H-W平衡。
2.2.2 高要子代群體 表3 可見:高要子代群體POU1F1基因12 個(gè)SNP 位點(diǎn)的多樣性比較豐富,Ne、Ho、He平均值分別為1.1873、0.1542、0.1556;PIC平均為0.155,12個(gè)位點(diǎn)均低度多態(tài)(PIC <0.25);12個(gè)位點(diǎn)在群體中均符合H-W平衡。

表3 POU1F1基因12個(gè)SNP位點(diǎn)在尼羅羅非魚高要子代群體的遺傳信息Table 3 Genetic information of twelve SNP loci of POU1F1 gene in Gaoyao offspring population of Nile tilapia
2.2.3 番禺群體 表4 可知:番禺群體POU1F1基因12 個(gè)SNP 位點(diǎn)的多樣性比較豐富,Ne、Ho、He平均值分別為1.2643、0.2038、0.2044;PIC 平均為0.182,有10 個(gè)位點(diǎn)低度多態(tài)(PIC <0.25),2 個(gè)位點(diǎn)高度多態(tài);有11 個(gè)位點(diǎn)在群體中均符合H-W 平衡,有1 個(gè)位點(diǎn)偏離H-W平衡。

表4 POU1F1基因12個(gè)SNP位點(diǎn)在尼羅羅非魚番禺群體的遺傳信息Table 4 Genetic information of twelve SNP loci of POU1F1 gene in Panyu population of Nile tilapia
2.2.4 海南群體 表5 表明:海南群體雌性個(gè)體POU1F1基因6 個(gè)SNP 位點(diǎn)的多樣性比較豐富,Ne、Ho、He平均值分別為1.2918、0.1933、0.2201;PIC 平均為0.202,有5 個(gè)位點(diǎn)低度多態(tài)(PIC <0.25);有3個(gè)位點(diǎn)在群體中均符合H-W 平衡,有3 個(gè)位點(diǎn)偏離H-W平衡。
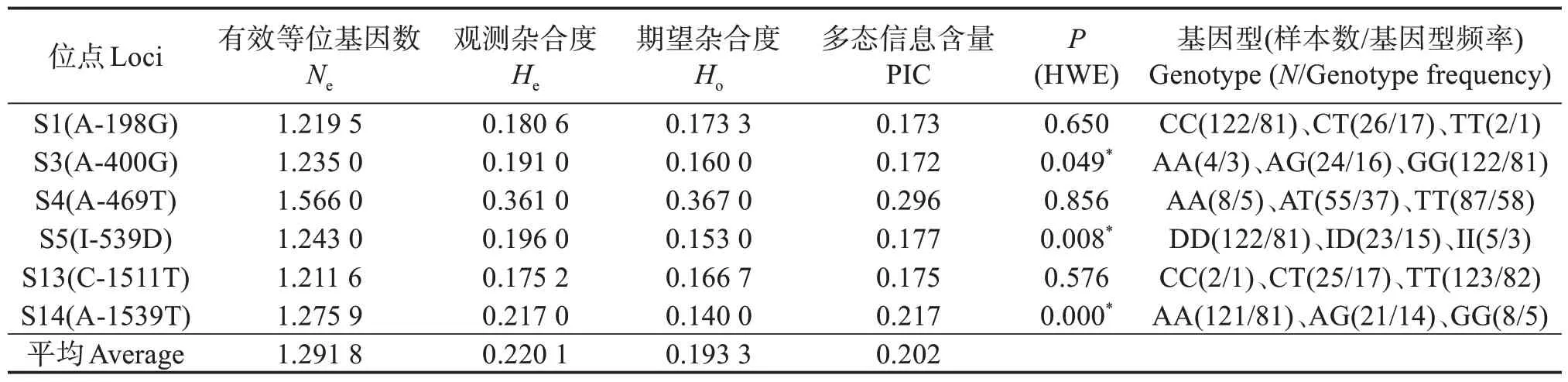
表5 POU1F1基因3個(gè)SNP位點(diǎn)在尼羅羅非魚海南雌性群體的遺傳信息Table 5 Genetic information of 3 SNP loci of POU1F1 gene in Hainan female population of Nile tilapia
表6表明:海南群體雄性個(gè)體POU1F1基因6個(gè)SNP 位點(diǎn)的多樣性比較豐富,Ne、Ho、He平均值分別為1.2434、0.1767、0.1936;PIC 平均為0.185,6 個(gè)位點(diǎn)均低度多態(tài)(PIC <0.25);有5 個(gè)位點(diǎn)在群體中符合H-W平衡(表6),有1個(gè)位點(diǎn)偏離H-W平衡。
2.3 POU1F1基因SNP位點(diǎn)與各群體體質(zhì)量和形態(tài)性狀的相關(guān)性
2.3.1 高要親代群體 表7 表明:POU1F1基因S3(A-400G)位點(diǎn)的AA 型個(gè)體與AG、GG 型個(gè)體在頭長(zhǎng)上有顯著差異。S4(A-469T)位點(diǎn)TT、AT 型與AA型在體質(zhì)量與全長(zhǎng)上均有顯著差異。S5(I-539D)位點(diǎn)DD 型和ID 型與II 型在體寬上有顯著差異。S6(A-881G)位點(diǎn)AA 型與AG、GG 型在頭長(zhǎng)上存在顯著差異。S7(A-888G)位點(diǎn)的GG 型與AA、AG 型在頭長(zhǎng)上存在顯著差異。S12(C-1365T)位點(diǎn)的CT 與TT型在體質(zhì)量和全長(zhǎng)、體高、體寬上存在顯著差異,該位點(diǎn)CC 與TT 型在體長(zhǎng)上有顯著差異,CC 與CT型在頭長(zhǎng)上有顯著差異。差異顯著性水平均為0.05。

表7 POU1F1基因中與尼羅羅非魚高要親代群體體質(zhì)量或形態(tài)性狀顯著相關(guān)的6個(gè)位點(diǎn)Table 7 6 SNP loci in POU1F1 gene significantly associated with body mass or morphological traits of Gaoyao parent population of Nile tilapia
2.3.2 高要子代群體 將高要親代群體中篩選出的與體質(zhì)量和形態(tài)相關(guān)聯(lián)的SNP 位點(diǎn),在高要子代群體中進(jìn)行分型分析,這些SNP 位點(diǎn)各基因型與高要子代群體體質(zhì)量和形態(tài)性狀關(guān)聯(lián)分析表明:S3(A-400G)位點(diǎn)的AA型個(gè)體與AG、GG型個(gè)體間在體質(zhì)量、全長(zhǎng)、體長(zhǎng)、頭長(zhǎng)、體高、體寬方面均有顯著差異;AA 型個(gè)體各指標(biāo)平均值均低于AG、GG 型個(gè)體。S5(I-539D)位點(diǎn)的DD 型個(gè)體與ID、II型個(gè)體間在體質(zhì)量、全長(zhǎng)、體長(zhǎng)、頭長(zhǎng)、體高、體寬方面均存在顯著差異;S11(I-1358D)位點(diǎn)ID 型與II 型在體質(zhì)量和體高方面有顯著差異,ID 型個(gè)體體質(zhì)量和體高顯著高于TT 型;S13(C-1511T)位點(diǎn)CC 型與CT、TT 型個(gè)體間在體質(zhì)量、體長(zhǎng)、頭長(zhǎng)、體高、體寬方面均有顯著差異,CC 型個(gè)體平均值均高于CT、TT 型;S14(A-1539T)位點(diǎn)AT 型與TT 型個(gè)體間在體質(zhì)量和體高方面有顯著差異,AT 型個(gè)體的體質(zhì)量、體高均值均高于TT型(表8)。
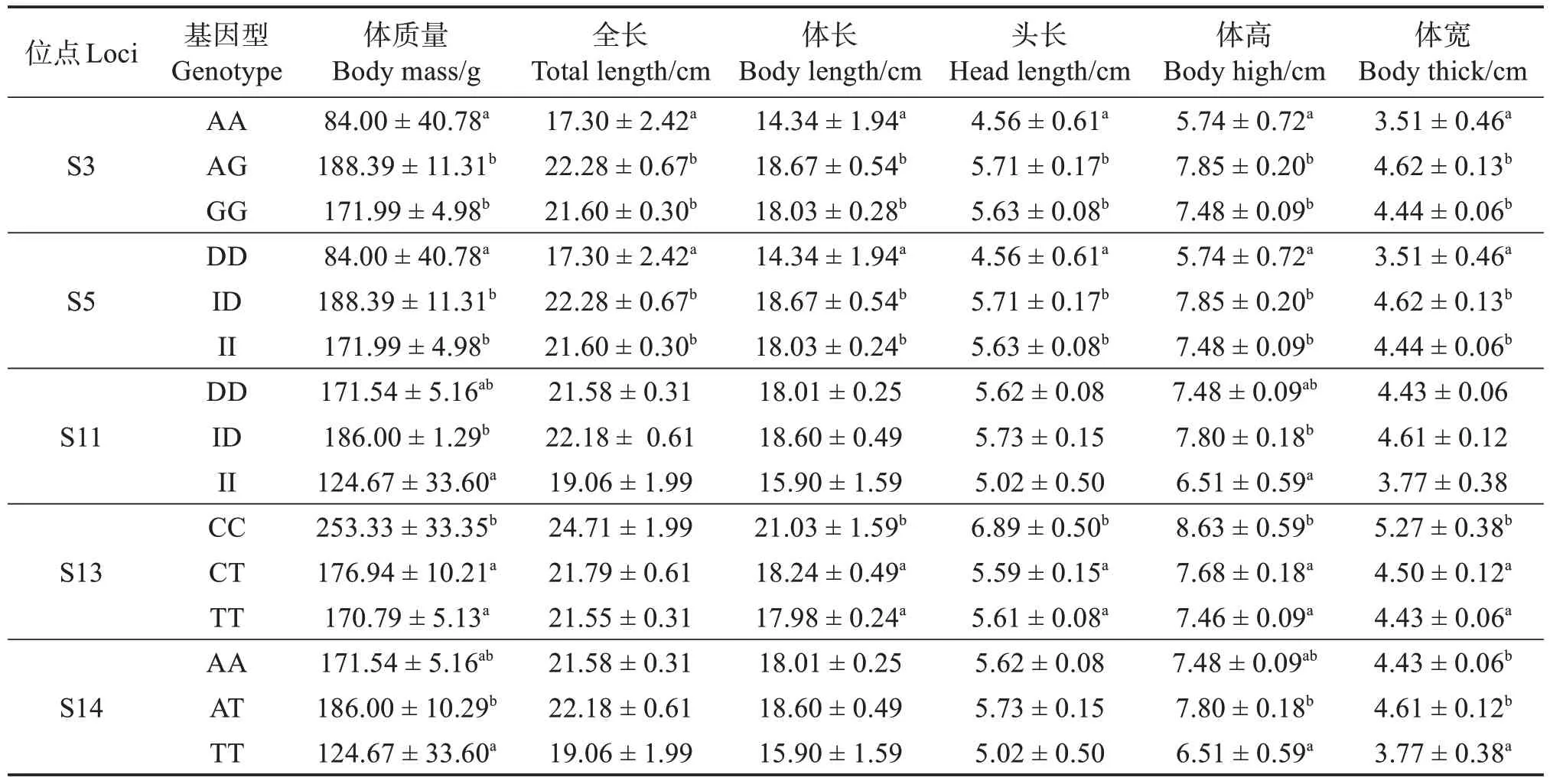
表8 POU1F1基因中與尼羅羅非魚高要子代群體體質(zhì)量或形態(tài)性狀顯著相關(guān)的5個(gè)位點(diǎn)Table 8 5 SNP loci in POU1F1 gene significantly associated with body mass or morphological traits of Gaoyao offspring population of Nile tilapia
2.3.3 番禺群體POU1F1基因SNP 位點(diǎn)與番禺群體體質(zhì)量和形態(tài)性狀關(guān)聯(lián)分析表明,12 個(gè)SNP 位點(diǎn)與番禺群體體質(zhì)量和形態(tài)性狀均無(wú)關(guān)聯(lián)(數(shù)據(jù)略)。
2.3.4 海南群體POU1F1基因SNP 位點(diǎn)與海南雌性群體體質(zhì)量性狀關(guān)聯(lián)分析表明,S4(A-469T)位點(diǎn)與海南雌性群體體質(zhì)量具有相關(guān)性,AA 型個(gè)體體質(zhì)量明顯高于AT、TT型(表9)。

表9 POU1F1基因中各SNP位點(diǎn)與尼羅羅非魚海南雌、雄性群體體質(zhì)量性狀的關(guān)聯(lián)分析Table 9 Association analysis of SNPs in POU1F1 gene with body mass traits of Hainan female and male population of Nile tilapia
表9 表明,海南雌性群體S3(A-400G)和S5(I-539D)位點(diǎn)與體質(zhì)量性狀顯著相關(guān),S3(A-400G)位點(diǎn)的AA 型個(gè)體體質(zhì)量顯著高于AG、GG 型個(gè)體,S5(I-539D)位點(diǎn)的DD型個(gè)體體質(zhì)量顯著高于ID、II型個(gè)體。
2.4 POU1F1基因雙倍型與各群體體質(zhì)量和形態(tài)性狀的關(guān)聯(lián)分析
2.4.1 高要親代群體 將27 個(gè)SNPs 位點(diǎn)進(jìn)行單倍型分析,得出雙倍型(圖1,表10),除去出現(xiàn)頻率低于3%的雙倍型,將其余5種雙倍型與體質(zhì)量和形態(tài)性狀進(jìn)行關(guān)聯(lián)性分析,結(jié)果表明:雙倍型D4、D7 個(gè)體的體寬顯著高于雙倍型D8 的個(gè)體。在實(shí)際的育種應(yīng)用中,應(yīng)選擇D4、D7雙倍型個(gè)體,以增加體寬。

圖1 尼羅羅非魚POU1F1基因SNPs在高要親代群體中的連鎖不平衡分析Fig.1 Map of pair-wise LD between POU1F1 SNPs in Gaoyao parent populations of Nile tilapia

表10 POU1F1基因雙倍型與尼羅羅非魚高要親代群體體質(zhì)量和形態(tài)性狀關(guān)聯(lián)分析Table 10 Correlation analysis between body mass and morphological traits and diplotype in the POU1F1 gene in Gaoyao parent population of Nile tilapia
2.4.2 高要子代群體 將12 個(gè)SNPs 位點(diǎn)進(jìn)行單倍型分析,利用單倍型分析出雙倍型,除去出現(xiàn)頻率低于3%的雙倍型,將其余2種雙倍型與體質(zhì)量和形態(tài)性狀進(jìn)行關(guān)聯(lián)性分析(表11)。結(jié)果表明,2 種雙倍型與高要子代群體體質(zhì)量和形態(tài)性狀均無(wú)關(guān)聯(lián)。

表11 POU1F1基因雙倍型與尼羅羅非魚高要子代群體體質(zhì)量和形態(tài)性狀關(guān)聯(lián)分析Table 11 Correlation analysis between body mass and morphological traits and diplotype in the POU1F1 gene in Gaoyao offspring population of Nile tilapia
2.4.3 番禺群體 將12 個(gè)SNPs 位點(diǎn)進(jìn)行單倍型分析,利用單倍型分析出雙倍型,除去出現(xiàn)頻率低于3%的雙倍型,將其余3種雙倍型與體質(zhì)量和形態(tài)性狀進(jìn)行關(guān)聯(lián)性分析發(fā)現(xiàn),3 種雙倍型與番禺群體體質(zhì)量和形態(tài)性狀均無(wú)關(guān)聯(lián)(表12)。

表12 POU1F1基因雙倍型與尼羅羅非魚番禺群體體質(zhì)量和形態(tài)性狀關(guān)聯(lián)分析Table 12 Correlation analysis between body mass and morphological traits and diplotype in POU1F1 gene in Panyu population of Nile tilapia
2.4.4 海南群體 6 個(gè)SNP 位點(diǎn)在雄性群體中獲得的雙倍型,除去出現(xiàn)頻率低于3%的雙倍型,剩余4種雙倍型。關(guān)聯(lián)性分析發(fā)現(xiàn),4 種雙倍型與海南雄性群體體質(zhì)量性狀均無(wú)關(guān)聯(lián)(表13)。

表13 POU1F1基因雙倍型與尼羅羅非魚海南雄性群體生長(zhǎng)性狀關(guān)聯(lián)分析Table 13 Correlation analysis between body mass and diplotype in POU1F1 gene in Hainan male population of Nile tilapia
6 個(gè)SNP 位點(diǎn)在雌性群體中獲得的雙倍型,除去出現(xiàn)頻率低于3%的雙倍型,剩余5種雙倍型。關(guān)聯(lián)性分析發(fā)現(xiàn),雙倍型為D6 的個(gè)體體質(zhì)量顯著高于雙倍型為D2的個(gè)體(表14,圖2)。

圖2 尼羅羅非魚POU1F1基因SNPs在海南雌性群體中的連鎖不平衡分析Fig.2 Map of pair-wise LD between POU1F1 SNPs in Hainan female population of Nile tilapia
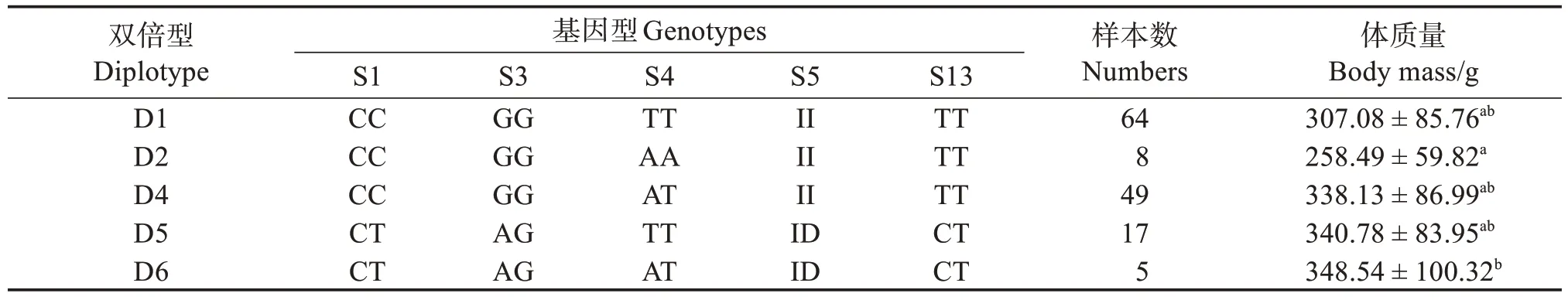
表14 POU1F1基因雙倍型與尼羅羅非魚海南雌性群體體質(zhì)量性狀關(guān)聯(lián)分析Table 14 Correlation analysis between body mass traits and diplotype in the POU1F1 gene in Hainan female population of Nile tilapia
3 討論
3.1 POU1F1多態(tài)性
單基因的遺傳多樣性可反映某群體的遺傳多樣性,某個(gè)群體的遺傳多樣性比較低,則在某個(gè)基因上可反映出來(lái)[17],本研究從單基因多態(tài)性方面分析群體遺傳多樣性,有一定實(shí)用性。
等位基因數(shù)、雜合度和多態(tài)信息含量等遺傳參數(shù)均可反映直接反映群體的基因豐富度和遺傳多樣性[18]。本研究的高要子代群體Ne、Ho兩個(gè)多樣性指標(biāo)與親代群體相當(dāng),但He和PIC 低于親代群體。可能由子代群體分析中所采用的SNP 位點(diǎn)數(shù)少于親代群體導(dǎo)致,子代群體分析只采用了與親代群體生長(zhǎng)相關(guān)的位點(diǎn)。高要親代群體PIC 均值為0.298,番禺群體為0.182,海南雌性群體為0.202,均稍低于王春曉等2015 年對(duì)這兩個(gè)群體在GHSR基因中檢測(cè)到的多樣性(0.278,0.310)[19]。這可能是因?yàn)楸狙芯克玫母咭后w和番禺群體經(jīng)過幾年的選育后多樣性有所下降,也可能是因?yàn)椴煌虻腟NP 多樣性有所不同。本研究表明,各羅非魚群體具有較高的遺傳多樣性,可在此基礎(chǔ)上繼續(xù)進(jìn)行良種選育。
3.2 生長(zhǎng)相關(guān)位點(diǎn)的篩選
本研究篩選生長(zhǎng)相關(guān)SNP 位點(diǎn)所用的區(qū)域均位于POU1F1基因的3?端非編碼區(qū)。這些位于非編碼區(qū)的SNP 位點(diǎn),不參與蛋白編碼,也不能改變蛋白序列及結(jié)構(gòu),因此這些基因座對(duì)性狀的影響可能不直接,但這些位于非編碼區(qū)的突變位點(diǎn)可能與負(fù)責(zé)改變蛋白序列及結(jié)構(gòu)的位點(diǎn)連鎖不平衡(具有連鎖不平衡的位點(diǎn)可影響動(dòng)物的生產(chǎn)性狀[20])。單倍型或雙倍型與性狀進(jìn)行關(guān)聯(lián)分析,解決了單個(gè)標(biāo)記分子存在的位點(diǎn)檢測(cè)和統(tǒng)計(jì)效率低等問題,從而能更真實(shí)反映所分析位點(diǎn)與性狀的關(guān)聯(lián)性[21]。王春曉等[19]在尼羅羅非魚GHSR基因中篩選的3 個(gè)SNP位點(diǎn)均與生長(zhǎng)性狀無(wú)關(guān)聯(lián),但3 個(gè)位點(diǎn)組成的雙倍型與生長(zhǎng)性狀顯著相關(guān)。本研究在高要親代群體中獲得體寬相關(guān)雙倍型2 個(gè),在海南雌性群體中獲得與體質(zhì)量相關(guān)的雙倍型1個(gè);但在高要子代群體、海南雄性群體中獲得與生長(zhǎng)性狀相關(guān)的SNP 位點(diǎn),而未獲得與生長(zhǎng)性狀相關(guān)雙倍型:SNP 位點(diǎn)之間出現(xiàn)了拮抗。這與李紅霞等[22]對(duì)建鯉(Cyprinus carpiovar.Jian)鳥氨酸脫酶基因多態(tài)性與生長(zhǎng)性狀的關(guān)聯(lián)分析結(jié)果類似,不同位點(diǎn)之間可能存在拮抗或協(xié)同的互作。
3.3 生長(zhǎng)相關(guān)SNP位點(diǎn)在各群體中的普適性
本研究在高要親代群體中獲得的體質(zhì)量和形態(tài)相關(guān)位點(diǎn),與子代中篩選到的相關(guān)位點(diǎn)部分重合,但是親子代群體與重合位點(diǎn)關(guān)聯(lián)的體質(zhì)量和形態(tài)性狀有所不同,如:親代群體位點(diǎn)S3 與頭長(zhǎng)顯著相關(guān),S4 與體質(zhì)量、全長(zhǎng)相關(guān),S5 與體寬顯著相關(guān),S11 與頭長(zhǎng)顯著相關(guān);但在子代群體中,位點(diǎn)S3 和S5與體質(zhì)量、全長(zhǎng)、體長(zhǎng)、頭長(zhǎng)、體高、體寬均顯著相關(guān),S11 與體質(zhì)量和體高顯著相關(guān)。這可能是親代群體與子代群體生長(zhǎng)狀態(tài)不同導(dǎo)致的;也可能是因?yàn)轸~類生長(zhǎng)性狀是受多種基因控制的數(shù)量性狀,主效基因和微效基因在不同世代中會(huì)出現(xiàn)分離或整合現(xiàn)象,從而導(dǎo)致篩選的生長(zhǎng)相關(guān)SNP 位點(diǎn)在不同世代之間的適用性較差[23];還可能是本研究子代群體采樣數(shù)量少于親本導(dǎo)致的,增加子代采樣數(shù)量,有可能提高親代子代之間生長(zhǎng)相關(guān)位點(diǎn)的契合度。本研究在高要子代群體中獲得的SNP 位點(diǎn)與番禺群體中相關(guān)SNP 位點(diǎn)沒有重疊,在番禺群體未篩選到體質(zhì)量和形態(tài)性狀相關(guān)位點(diǎn)。高要子代中與生長(zhǎng)性狀顯著相關(guān)的SNP 位點(diǎn),在番禺群體中各基因型的體質(zhì)量數(shù)據(jù)亦有部分差異,只是在統(tǒng)計(jì)學(xué)上未達(dá)到顯著性水平。這可能由取樣數(shù)量導(dǎo)致。影響標(biāo)記-性狀連鎖分析準(zhǔn)確性的一個(gè)重要因素是群體數(shù)量;但目前對(duì)用于標(biāo)記-性狀連鎖分析的群體數(shù)量須達(dá)到什么標(biāo)準(zhǔn)篩選出的標(biāo)記才可信,尚無(wú)明確的依據(jù),因而并不能說(shuō)在另外一個(gè)群體中未得到驗(yàn)證的關(guān)聯(lián)標(biāo)記不可信[24]。與高要子代群體生長(zhǎng)性狀相關(guān)的這些位點(diǎn)在海南群體中進(jìn)行進(jìn)一步驗(yàn)證表明:僅S4 位點(diǎn)與海南雌性群體體質(zhì)量顯著相關(guān),S3 和S5 位點(diǎn)與雄性群體體質(zhì)量相關(guān),其余位點(diǎn)與體質(zhì)量性狀均不具有相關(guān)性。這可能是因?yàn)椴煌后w的遺傳背景差異會(huì)導(dǎo)致篩選到的生長(zhǎng)相關(guān)SNP 位點(diǎn)在不同群體中的關(guān)聯(lián)性急劇下降。
本實(shí)驗(yàn)獲得的與高要親代群體生長(zhǎng)相關(guān)位點(diǎn),在高要親代與子代之間普適性較弱,可能是因?yàn)槿訑?shù)量不夠大,導(dǎo)致親代、子代之間生長(zhǎng)相關(guān)位點(diǎn)有些差異;這些與高要親代群體生長(zhǎng)相關(guān)SNP 位點(diǎn)在不同群體間的普適性較差,可能是不同群體間遺傳背景的差異導(dǎo)致,也可能是因?yàn)槿訑?shù)量不夠大導(dǎo)致。擴(kuò)大群體取樣數(shù)量,這些生長(zhǎng)相關(guān)SNP 標(biāo)記在不同群體之間的普適性可能就會(huì)提高。
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
電子制作(2018年18期)2018-11-14 01:48:24
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
汽車觀察(2016年3期)2016-02-28 13:16:26
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
中國(guó)質(zhì)量與標(biāo)準(zhǔn)導(dǎo)報(bào)(2014年1期)2014-02-28 22:21:28
