煙草未授粉子房離體培養(yǎng)誘導(dǎo)胚狀體的影響因素研究
2024-01-30 01:13:16曹景林程君奇李亞培吳成林張俊杰蔡長(zhǎng)春
中國(guó)煙草科學(xué) 2023年6期
關(guān)鍵詞:煙草
曹景林,王 欣,程君奇,李亞培,吳成林,張俊杰,蔡長(zhǎng)春
煙草未授粉子房離體培養(yǎng)誘導(dǎo)胚狀體的影響因素研究
曹景林,王 欣,程君奇*,李亞培,吳成林,張俊杰,蔡長(zhǎng)春
(湖北省煙草科學(xué)研究院,武漢 430030)
源于未授粉子房的雌性單倍體在煙草育種利用價(jià)值上優(yōu)于源于花藥或小孢子的雄性單倍體,開(kāi)展煙草未授粉子房的離體培養(yǎng)對(duì)煙草單倍體育種具有重要意義。為了建立未授粉子房離體培養(yǎng)技術(shù)體系,分別以不同的雜種F1代為試材,采用HW培養(yǎng)基探討了消毒方法、接種方式、接種密度、無(wú)機(jī)鹽濃度、外源激素配比和瓊脂濃度6個(gè)關(guān)鍵因素對(duì)胚狀體誘導(dǎo)的影響。結(jié)果表明:花蕾不需剝開(kāi)花瓣而直接放入0.1%升汞中浸泡8 min,剝開(kāi)子房壁并橫切子房后接種,每個(gè)125 mL三角瓶中接種2塊子房,培養(yǎng)基中無(wú)機(jī)鹽濃度額外增加60%、附加IAA 0.5 mg/L和6-BA 2 mg/L或者附加KT 2.0 mg/L和2,4-D 0.5 mg/L、瓊脂濃度9 g/L有助于對(duì)胚狀體的誘導(dǎo)。據(jù)此進(jìn)一步完善了煙草未授粉子房離體培養(yǎng)過(guò)程中的技術(shù)參數(shù)。
煙草;未授粉子房;離體培養(yǎng);胚狀體;影響因素
通過(guò)誘導(dǎo)單倍體再加倍能夠快速縮短育種時(shí)間,是目前作物快速、高效的育種途徑之一[1]。單倍體植株通常可以由花藥或小孢子體外培養(yǎng)誘導(dǎo)產(chǎn)生,稱作雄性單倍體;也可以由子房或胚珠離體培養(yǎng)誘導(dǎo)產(chǎn)生,稱作雌性單倍體[1-2]。與花藥或小孢子培養(yǎng)相比,經(jīng)未授粉子房或未受精胚珠離體培養(yǎng)誘導(dǎo)的煙草雌性單倍體植株至少有3個(gè)獨(dú)特的優(yōu)點(diǎn):(1)煙草雄性單倍體經(jīng)染色體加倍后往往產(chǎn)量和品質(zhì)性狀劣變嚴(yán)重,而雌性雙單倍體不會(huì)造成產(chǎn)量和品質(zhì)的劣變,若采用雜交種未受精的卵細(xì)胞培養(yǎng),則獲得的雙二倍體品系與常規(guī)自交相似,因而比雄性單倍體更具有育種利用價(jià)值[3-5];(2)由于未授粉子房或未受精胚珠離體培養(yǎng)不牽涉花粉,因而能夠適用于雄性不育植物[6],即使單倍體植株經(jīng)染色體加倍后得不到種子,也可利用組培技術(shù)繁殖種苗;(3)培養(yǎng)F1代的未授粉子房或未受精胚珠,可迅速獲得高純度種質(zhì)資源,對(duì)提高種質(zhì)資源利用率和育種效率具有理論研究和實(shí)際應(yīng)用價(jià)值[7]。基于此,國(guó)內(nèi)外學(xué)者越來(lái)越重視煙草未授粉子房或未受精胚珠的離體培養(yǎng),并在此方面做了許多探索性工作。
Burk等[8]于1979年開(kāi)始煙草未授粉子房培養(yǎng)研究,并成功獲得了單倍體植株。祝仲純等[9-10]采用H培養(yǎng)基(附加生長(zhǎng)素IAA 0.5 mg/L和細(xì)胞分裂素KT 2 mg/L)對(duì)普通煙草品種Copusyeusuheku No.4、NC2326和革新五號(hào)的未授粉子房進(jìn)行了離體培養(yǎng),采用N6培養(yǎng)基(附加生長(zhǎng)素IAA 0.5 mg/L、細(xì)胞分裂素KT 6 mg/L、肌醇100 mg/L和蔗糖8 g/L)對(duì)大葉黃煙的未授粉子房進(jìn)行了離體培養(yǎng)[11],均獲得了單倍體植株。吳伯驥等[12]采用改良的H培養(yǎng)基(附加生長(zhǎng)素IAA 0.5 mg/L和細(xì)胞分裂素KT 2 mg/L)對(duì)普通煙草品種柳葉煙、金星煙、黃苗榆和黃花煙草品種甘肅黃花煙的未授粉子房進(jìn)行離體培養(yǎng),也獲得了單倍體植株。潘莉等[13-14]曾以H培養(yǎng)基為基礎(chǔ),探討了基因型、外源激素、蔗糖、活性炭、椰胚乳、光照強(qiáng)度等因子對(duì)單倍體誘導(dǎo)率的影響。本研究組也以H培養(yǎng)基為基礎(chǔ),探討了未受精胚珠離體培養(yǎng)適宜的子房發(fā)育時(shí)期和培養(yǎng)基類型[3]。為了建立穩(wěn)定的煙草未授粉子房離體培養(yǎng)技術(shù)體系,提高胚狀體誘導(dǎo)頻率,本研究對(duì)前人研究[3, 9-14]中尚未涉及的消毒方法、接種方式、接種密度、無(wú)機(jī)鹽濃度、其他外源激素配比、瓊脂濃度等技術(shù)因子對(duì)胚狀體誘導(dǎo)率的影響進(jìn)行了探討,以期進(jìn)一步完善煙草未授粉子房離體培養(yǎng)過(guò)程中的技術(shù)因素參數(shù)。
1 材料與方法
1.1 試驗(yàn)材料
選用云煙87×吉煙9號(hào),K346×6388和Coker371Gold×龍江911這3個(gè)雜種F1代的未授粉子房為材料。為了保證整個(gè)試驗(yàn)期間均能得到子房,將3個(gè)雜交組合F1代分三期于溫室內(nèi)播種和盆栽,間隔60 d,采用的栽煙盆內(nèi)徑為30 cm,每次盆栽100株。
1.2 試驗(yàn)方法
根據(jù)本研究組先前的研究結(jié)果,選取花冠即將露出花萼時(shí)期的子房作為外植體[3],以由吳伯驥等[12]改良的H培養(yǎng)基(用HW表示)為基本培養(yǎng)基,附加IAA 0.5 mg/L、6-BA 2 mg/L、瓊脂8 g/L 和蔗糖20 g/L。在未授粉子房的離體培養(yǎng)過(guò)程中,設(shè)置外植體不同消毒方法、接種方式和接種密度,以及培養(yǎng)基不同無(wú)機(jī)鹽濃度、外源激素配比和瓊脂濃度對(duì)胚狀體誘導(dǎo)影響的試驗(yàn)。培養(yǎng)基中所有激素和氨基酸在滅菌前加入,并用40 g/L NaOH調(diào)節(jié)pH至5.5,高溫高壓滅菌15 min。采用125 mL三角瓶進(jìn)行子房培養(yǎng),每個(gè)三角瓶中倒入約30 mL培養(yǎng)基。胚狀體誘導(dǎo)培養(yǎng)在光照培養(yǎng)室中進(jìn)行,培養(yǎng)溫度為(28±2)℃,光照強(qiáng)度為1500 lx,光照時(shí)間為12 h/d。培養(yǎng)過(guò)程中時(shí)常觀察子房發(fā)育狀況,培養(yǎng)25 d左右統(tǒng)計(jì)試驗(yàn)結(jié)果。各試驗(yàn)過(guò)程均在無(wú)菌條件下進(jìn)行。
1.2.1 外植體不同消毒方法對(duì)胚狀體誘導(dǎo)的影響 以Coker371Gold×龍江911為材料,將所采集的花蕾分別采用以下4種方式進(jìn)行消毒:(X1)不剝開(kāi)花瓣,將花放入70%酒精浸泡30 s,0.1%升汞滅菌8 min;(X2)不剝開(kāi)花瓣,將花放入0.1%升汞浸泡8 min;(X3)剝開(kāi)花瓣,將子房放入70%酒精浸泡30 s,0.1%升汞滅菌8 min;(X4)剝開(kāi)花瓣,將子房放入0.1%升汞浸泡8 min。接著用無(wú)菌水沖洗 3 次,對(duì)于采用方式X1和X2進(jìn)行消毒的花蕾,在墊有濾紙的無(wú)菌培養(yǎng)皿上將花蕾基部橫切,用鑷子將子房從底部擠出,輕輕剝開(kāi)子房壁,橫切子房后接種至HW培養(yǎng)基;對(duì)于采用方式X3和X4進(jìn)行消毒的子房,用鑷子輕輕剝開(kāi)子房壁,橫切子房后接種至HW培養(yǎng)基。每個(gè)三角瓶中接種2塊子房,每個(gè)處理接種5瓶。
1.2.2 外植體不同接種方式對(duì)胚狀體誘導(dǎo)的影響 以K346×6388為材料,將所采集的花蕾首先按X1消毒方式進(jìn)行消毒處理,然后,在墊有濾紙的無(wú)菌培養(yǎng)皿上將消毒后的花蕾基部橫切,用鑷子把子房從底部擠出,分別按以下5種方式接種至HW培養(yǎng)基上進(jìn)行培養(yǎng):(S1)不剝開(kāi)子房壁,直接接種至培養(yǎng)基;(S2)不剝開(kāi)子房壁,橫切子房再接種至培養(yǎng)基;(S3)剝開(kāi)子房壁,直接接種至培養(yǎng)基;(S4)剝開(kāi)子房壁,橫切子房再接種至培養(yǎng)基;(S5)剝開(kāi)子房壁,切碎子房后接種至培養(yǎng)基。每個(gè)三角瓶中接種2塊子房,每個(gè)處理接種5瓶。
1.2.3 外植體不同接種密度對(duì)胚狀體誘導(dǎo)的影響 以云煙87×吉煙9號(hào)為材料,將所采集的花蕾先按X1消毒方式進(jìn)行消毒處理,然后采用S4接種方式將子房按以下5個(gè)不同密度分別接種至HW培養(yǎng)基上進(jìn)行培養(yǎng):(D1)1塊子房/瓶;(D2)2塊子房/瓶;(D3)3塊子房/瓶;(D4)4塊子房/瓶;(D5)5塊子房/瓶。每個(gè)處理接種20塊子房。
1.2.4 培養(yǎng)基不同無(wú)機(jī)鹽濃度對(duì)胚狀體誘導(dǎo)的影響 以云煙87×吉煙9號(hào)、K346×6388、Coker371Gold×龍江911為材料,將所采集的花蕾先按X1消毒方式進(jìn)行消毒處理,然后采用S4接種方式將子房分別接種至如表1所示的3種培養(yǎng)基上進(jìn)行培養(yǎng)。每個(gè)三角瓶中接種2塊子房,每個(gè)培養(yǎng)基接種20瓶。
1.2.5 培養(yǎng)基不同外源激素配比對(duì)胚狀體誘導(dǎo)的影響 以云煙87×吉煙9號(hào)為材料,將所采集的花蕾先按X1消毒方式進(jìn)行消毒處理,然后采用S4接種方式將子房分別接種于附加有不同外源激素配比的HW培養(yǎng)基上。共設(shè)置如表2所示的16種外源激素配比。每個(gè)三角瓶中接種2塊子房,每個(gè)處理接種10瓶。

表1 培養(yǎng)基不同濃度無(wú)機(jī)鹽配方
1.2.6 培養(yǎng)基不同瓊脂濃度對(duì)胚狀體誘導(dǎo)的影響 以云煙87×吉煙9號(hào)為材料,將所采集的花蕾先按X1消毒方式進(jìn)行消毒處理,然后采用S4接種方式將子房分別接種于瓊脂濃度不同的HW培養(yǎng)基上進(jìn)行培養(yǎng)。瓊脂濃度設(shè)置4個(gè)不同的梯度:(A1)7 g/L;(A2)8 g/L;(A3)9 g/L;(A4)10 g/L。每個(gè)三角瓶中接種2塊子房,每個(gè)處理接種10瓶。
2 結(jié) 果
2.1 外植體不同消毒方法對(duì)胚狀體誘導(dǎo)的影響
外植體的不同消毒方法對(duì)胚狀體的誘導(dǎo)效果有所差異。從表3可以看出,4種消毒方法在子房污染率上差異不大,在2種不剝開(kāi)花瓣處理X1和X2以及2種剝開(kāi)花瓣處理X3和X4中,升汞滅菌前是否先用酒精浸泡對(duì)胚狀體誘導(dǎo)率也無(wú)太大影響。但是否剝開(kāi)花瓣對(duì)胚狀體誘導(dǎo)率則有明顯影響,2種不剝開(kāi)花瓣滅菌處理的胚狀體誘導(dǎo)率接近或達(dá)到90%;而2種剝開(kāi)花瓣滅菌處理的胚狀體誘導(dǎo)率則明顯下降,分別為55.56%和77.78%。綜合來(lái)看,在4種消毒方法中,不剝開(kāi)花瓣,也不用酒精浸泡,而將花蕾直接放入0.1%升汞中浸泡8 min為最優(yōu)。
2.2 外植體不同接種方式對(duì)胚狀體誘導(dǎo)的影響
外植體接種方式不同,其對(duì)胚狀體誘導(dǎo)的影響也不同(表4)。不剝開(kāi)子房壁而直接接種至培養(yǎng)基的方式未觀察到胚狀體產(chǎn)生,雖然接種5 d后子房有明顯膨大,但同時(shí)子房壁增厚,逐漸呈現(xiàn)翠綠色的玻璃化狀態(tài),胚珠發(fā)育受到抑制,最后褐化死亡。不剝開(kāi)子房壁,但橫切子房接種至培養(yǎng)基,5 d后子房膨脹,在切口處有胚狀體產(chǎn)生,但子房壁仍然變綠增厚、玻璃化嚴(yán)重,抑制胚珠發(fā)育。剝開(kāi)子房壁,將整個(gè)子房接種至培養(yǎng)基,5 d后子房緩慢膨脹,胚珠雖有發(fā)育但比較緩慢,10 d后子房底部褐化嚴(yán)重。剝開(kāi)子房壁,并將子房切碎成小塊接種至培養(yǎng)基,5 d后大部分外植體褐化死亡,胚狀體誘導(dǎo)率僅為30%。相比較而言,剝開(kāi)子房壁,并橫切子房的誘導(dǎo)效果最好,接種5 d后,子房膨脹,胚珠也有明顯發(fā)育。接種10 d后,大部分子房能產(chǎn)生胚狀體,誘導(dǎo)率達(dá)到90%。
2.3 外植體不同接種密度對(duì)胚狀體誘導(dǎo)的影響
外植體的接種密度對(duì)胚狀體的誘導(dǎo)也有影響,統(tǒng)計(jì)結(jié)果(表5)表明,子房接種密度由1塊/瓶增加到2塊/瓶時(shí),胚狀體誘導(dǎo)率由82.35%增加到90.00%。而后,隨著接種密度的增大,胚狀體誘導(dǎo)率開(kāi)始逐漸下降,當(dāng)接種密度增加到5塊/瓶時(shí),胚狀體誘導(dǎo)率下降到52.94%。可見(jiàn),采用2塊子房/瓶的接種密度最優(yōu)。
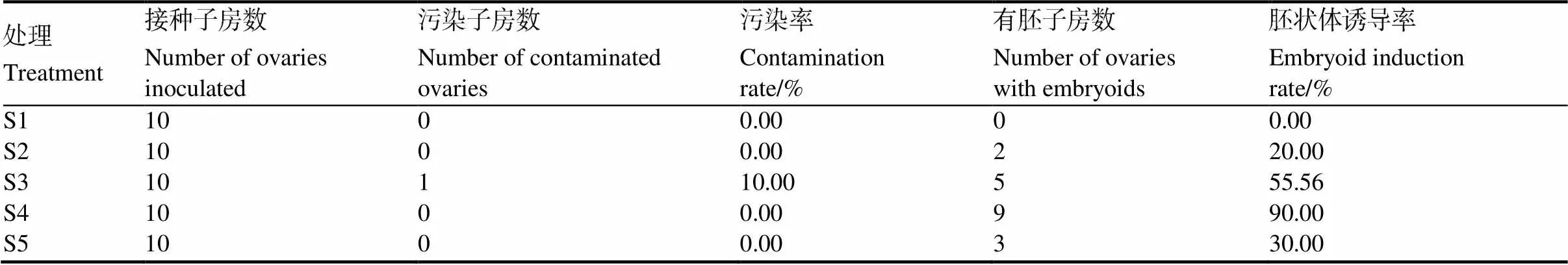
表4 子房不同接種方式對(duì)胚狀體誘導(dǎo)的影響
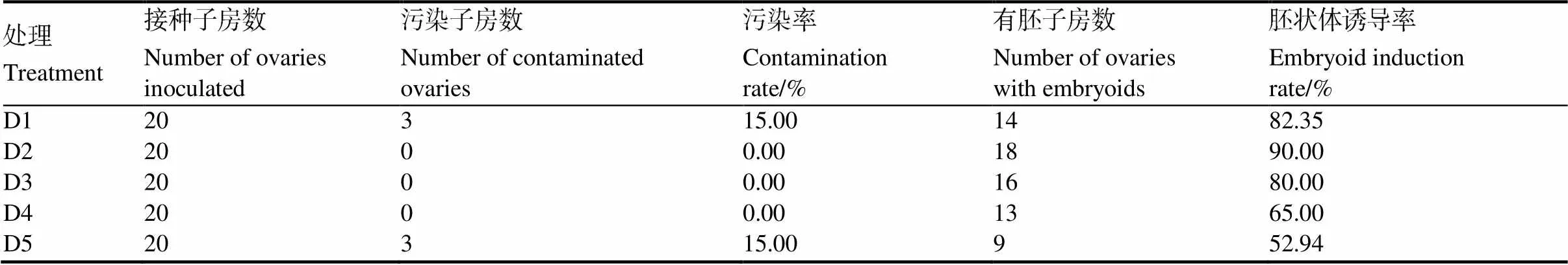
表5 子房不同接種密度對(duì)胚狀體誘導(dǎo)的影響
2.4 培養(yǎng)基不同無(wú)機(jī)鹽濃度對(duì)胚狀體誘導(dǎo)的影響
培養(yǎng)基中的無(wú)機(jī)鹽濃度對(duì)胚狀體的誘導(dǎo)影響很大。從表6可知,3個(gè)雜種F1代的未授粉子房在3種培養(yǎng)基上培養(yǎng)的表現(xiàn)趨勢(shì)是一致的。當(dāng)培養(yǎng)基中不另加無(wú)機(jī)鹽(即M1)時(shí),3個(gè)雜種F1代的未授粉子房接種后很快白化,約20 d后大部分褐化死亡,胚狀體誘導(dǎo)率都較低,平均僅有7.88%。當(dāng)培養(yǎng)基中的無(wú)機(jī)鹽濃度額外增加60%(即M2)時(shí),3個(gè)雜種F1代未授粉子房離體培養(yǎng)的情況均有明顯好轉(zhuǎn)。子房接種5 d后就開(kāi)始膨脹,15 d后能膨脹到直徑15 mm左右。胚狀體誘導(dǎo)率平均達(dá)到90.28%。當(dāng)培養(yǎng)基中的無(wú)機(jī)鹽濃度加倍(即M3)時(shí),3個(gè)雜種F1代未授粉子房接種5 d后均表現(xiàn)為膨脹,但10 d后部分子房褐化死亡,胚狀體誘導(dǎo)率較培養(yǎng)基M2均有所下降,平均為65.82%。綜上可見(jiàn),HW培養(yǎng)基中的無(wú)機(jī)鹽濃度額外增加60%有助于對(duì)胚狀體的誘導(dǎo)。
此外,3個(gè)雜種F1代的未授粉子房在3種無(wú)機(jī)鹽濃度不同的培養(yǎng)基上進(jìn)行培養(yǎng),胚狀體誘導(dǎo)效果因未授粉子房供體材料不同而有所差異,K346× 6388 F1未授粉子房在不同培養(yǎng)基上的胚狀體誘導(dǎo)率除M1培養(yǎng)基外均最高,平均誘導(dǎo)率也最高,達(dá)58.89%,而Coker371Gold×龍江911 F1未授粉子房在不同培養(yǎng)基上的胚狀體誘導(dǎo)率均最低,平均為51.32%。
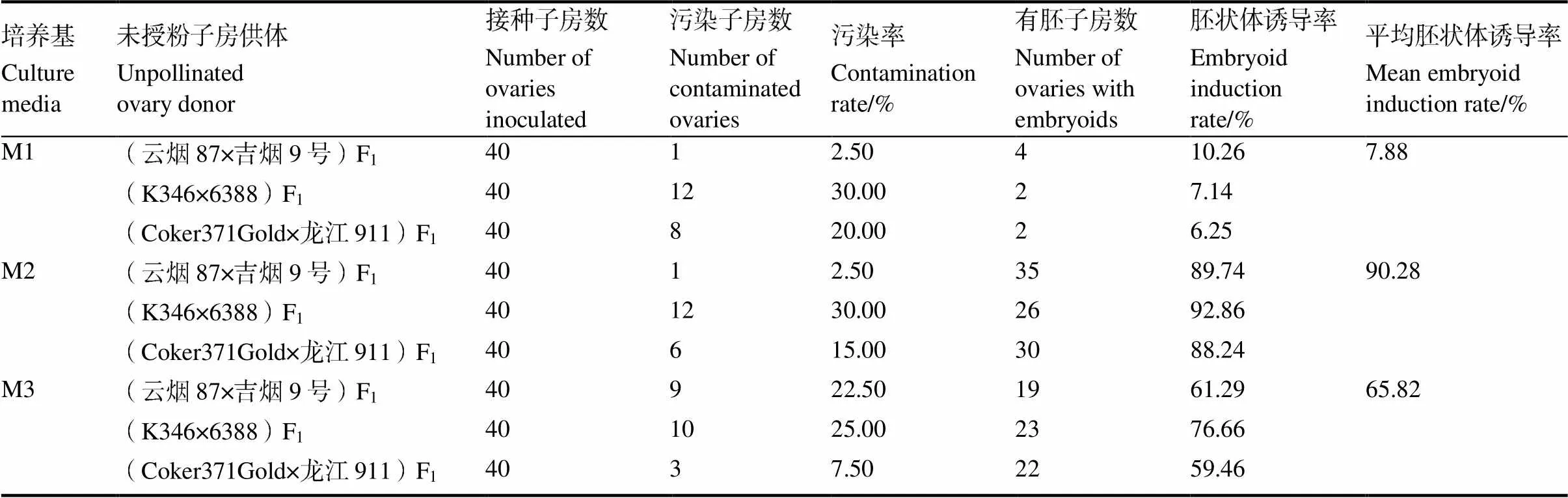
表6 不同無(wú)機(jī)鹽濃度對(duì)胚狀體誘導(dǎo)的影響
2.5 培養(yǎng)基不同外源激素配比對(duì)胚狀體誘導(dǎo)的影響
培養(yǎng)基中外源激素種類和配比不同,對(duì)胚狀體的誘導(dǎo)效果有很大差異。統(tǒng)計(jì)結(jié)果(表7)表明,在16種不同外源激素配比中,外源激素配比為IAA 0.5 mg/L、6-BA 2 mg/L和配比為KT 2.0 mg/L、2,4-D 0.5 mg/L的培養(yǎng)基誘導(dǎo)胚狀體的效果最好,誘導(dǎo)率均幾乎達(dá)到90%;其次是外源激素配比為IAA 0.5 mg/L、2,4-D 0.5 mg/L和配比為KT 2.0 mg/L、6-BA 2 mg/L,胚狀體誘導(dǎo)率在85%左右;而外源激素配比為IAA 1.0 mg/L、2,4-D 1.0 mg/L,配比為IAA 1.0 mg/L、2,4-D 0.5 mg/L和配比為KT 2.0 mg/L、6-BA 1 mg/L的培養(yǎng)基誘導(dǎo)胚狀體的效果最差,誘導(dǎo)率均不超過(guò)50%。可見(jiàn),在用HW培養(yǎng)基對(duì)煙草未授粉子房進(jìn)行離體培養(yǎng)時(shí),培養(yǎng)基中以附加IAA 0.5 mg/L和6-BA 2 mg/L或者附加KT 2.0 mg/L和2,4-D 0.5 mg/L為佳。
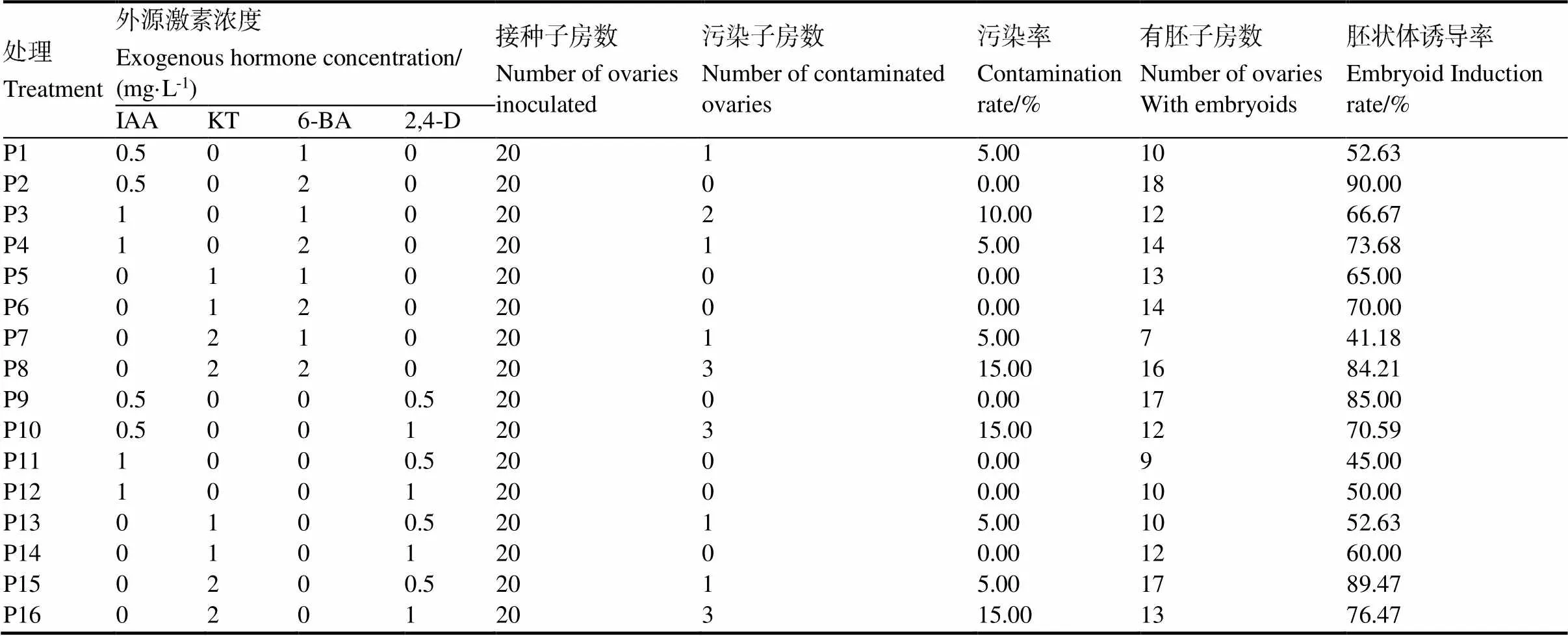
表 7 不同外源激素配比胚狀體誘導(dǎo)的影響
2.6 不同瓊脂濃度對(duì)胚狀體誘導(dǎo)的影響
培養(yǎng)基中瓊脂濃度不同對(duì)胚狀體誘導(dǎo)也有影響。從表8中可知,培養(yǎng)基中瓊脂濃度為7 g/L的情況下,子房玻璃化率達(dá)到90.00%,因而胚狀體誘導(dǎo)率較低,僅有50.00%;隨著瓊脂濃度的增加,子房玻璃化現(xiàn)象逐漸減輕,胚狀體誘導(dǎo)率逐漸提高,當(dāng)瓊脂濃度增加到9 g/L時(shí),胚狀體誘導(dǎo)率最高,達(dá)89.47%;但隨著瓊脂濃度增加到10 g/L時(shí),子房玻璃化現(xiàn)象雖然大幅減輕,但有相當(dāng)部分子房很快褐化死亡,胚狀體誘導(dǎo)率反而下降到72.22%。綜合考慮,培養(yǎng)基中的瓊脂濃度應(yīng)以9 g/L為宜。
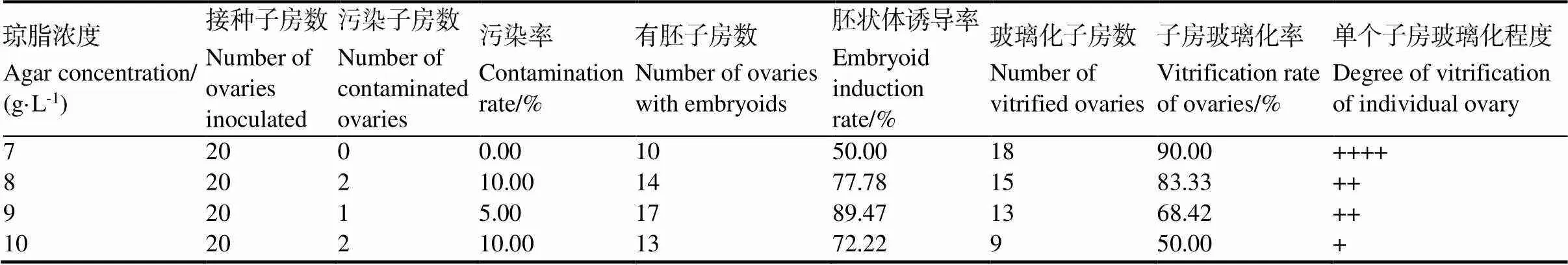
表8 不同瓊脂濃度對(duì)子房玻璃化的影響
3 討 論
外植體的消毒影響著子房在培養(yǎng)過(guò)程中可能受污染的程度以及胚狀體誘導(dǎo)率的高低。在本研究條件下,各種消毒方法均有較好的滅菌效果,但是剝開(kāi)花瓣滅菌處理會(huì)致使胚狀體誘導(dǎo)率下降,這說(shuō)明滅菌處理溶液直接浸泡子房,可能會(huì)對(duì)子房的分化能力有一定的傷害。雖然升汞滅菌前先用酒精浸泡與不用酒精浸泡相比,對(duì)胚狀體誘導(dǎo)率無(wú)太大影響,但增加了滅菌步驟和成本。可見(jiàn),一種好的消毒方法應(yīng)該步驟簡(jiǎn)單,既能有效滅掉外植體表面的雜菌,又不妨礙外植體的生長(zhǎng)。
子房的接種包括接種方式和接種密度2個(gè)方面。祝仲純等[9-11]、吳伯驥等[12]和潘莉等[13-14]采用煙草子房直接接種能夠誘導(dǎo)出胚狀體,但本研究采用子房直接接種,很少有胚狀體產(chǎn)生,這主要是由于在培養(yǎng)過(guò)程中子房壁增厚,玻璃化嚴(yán)重,抑制了胚珠的發(fā)育。而去除子房壁后接種,則能觀察到許多胚狀體。這些結(jié)果與先前的研究[3]是一致的。本研究結(jié)果顯示,去除子房壁并橫切子房后接種,能獲得較好的胚狀體誘導(dǎo)效果,但若將子房切碎接種則胚狀體誘導(dǎo)率下降,這可能是由于外植體受損嚴(yán)重,毒性酚類物質(zhì)或其氧化物醌類物質(zhì)在培養(yǎng)基中積累并抑制了其他酶的活性[15],致使外植體褐化死亡而導(dǎo)致的。關(guān)于煙草子房培養(yǎng)中的密度效應(yīng)迄今未見(jiàn)報(bào)道。在本研究中,子房接種密度由1塊/瓶增加到2塊/瓶,可能更有助于培養(yǎng)基的條件化,致使子房組織釋放到培養(yǎng)基中的某些物質(zhì)能夠促進(jìn)子房多細(xì)胞結(jié)構(gòu)的形成,進(jìn)而誘導(dǎo)胚狀體的產(chǎn)生[16]。而當(dāng)接種密度增加后,可能由于培養(yǎng)基營(yíng)養(yǎng)供應(yīng)不足,同時(shí)子房產(chǎn)生的有害次生代謝物增多,而致使子房褐化嚴(yán)重,胚狀體誘導(dǎo)率下降[15]。
培養(yǎng)基是未授粉子房離體培養(yǎng)成敗的關(guān)鍵。培養(yǎng)基中無(wú)機(jī)鹽成分是植物生長(zhǎng)發(fā)育所必需的。無(wú)論前人采用H培養(yǎng)基或HW培養(yǎng)基,無(wú)機(jī)鹽成分及其濃度基本上沒(méi)有變化[9-14]。而本研究采用無(wú)機(jī)鹽濃度未改變的HW培養(yǎng)基時(shí),很少有胚狀體產(chǎn)生,這可能是無(wú)機(jī)鹽營(yíng)養(yǎng)不足所致。當(dāng)增大培養(yǎng)基中無(wú)機(jī)鹽濃度時(shí),情況明顯好轉(zhuǎn),這說(shuō)明在HW培養(yǎng)基中,采用更高的無(wú)機(jī)鹽濃度有利于對(duì)胚狀體的誘導(dǎo)。但無(wú)機(jī)鹽濃度額外增加到2倍時(shí),胚狀體誘導(dǎo)率下降,這主要是由于鹽離子濃度過(guò)高而導(dǎo)致了部分子房褐化死亡[15]。培養(yǎng)基中添加外源激素在未授粉子房離體培養(yǎng)過(guò)程中也有利于胚狀體誘導(dǎo)[1,7,13,17]。在煙草子房培養(yǎng)中,常用的外源激素包括生長(zhǎng)素類激素IAA、2,4-D和細(xì)胞分裂素類激素6-BA和KT[9-14]。祝仲純等[9-11]和吳伯驥等[12]采用IAA 0.5mg/L與KT 2 mg/L或KT 6 mg/L配合使用對(duì)煙草子房進(jìn)行了離體培養(yǎng)。潘莉等[13]又探討了2,4-D和6-BA的不同濃度配比對(duì)煙草子房培養(yǎng)的作用。為了進(jìn)一步篩選培養(yǎng)基中附加的有助于提高胚狀體誘導(dǎo)率的外源激素及其濃度,本研究在前人研究[9-14,17]的基礎(chǔ)上,又另外探討了IAA與6-BA,2,4-D與KT,以及IAA與2,4-D,KT與6-BA的不同濃度配比對(duì)子房培養(yǎng)的作用,結(jié)果顯示,生長(zhǎng)素類激素同類配合或者細(xì)胞分裂素類激素同類配合使用對(duì)胚狀體的誘導(dǎo)效果總體上不如細(xì)胞分裂素類與生長(zhǎng)素類激素的配合使用,這與前人的研究是一致的[7,9-14]。本研究中,IAA與6-BA以及2,4-D與KT采用適當(dāng)?shù)臐舛扰浜鲜褂茫兄^好的胚狀體誘導(dǎo)效果。這進(jìn)一步說(shuō)明不同種類激素合理的濃度搭配,對(duì)胚狀體的誘導(dǎo)是非常重要的[1,13,17]。培養(yǎng)基中的瓊脂作為支持物,利于各種外植體的培養(yǎng)。在本研究中,隨著瓊脂濃度的增加,玻璃化程度逐漸減輕,但瓊脂濃度過(guò)高時(shí),胚狀體誘導(dǎo)率反而下降。這可能是由于瓊脂濃度低,使培養(yǎng)基內(nèi)水勢(shì)過(guò)高,造成培養(yǎng)器皿內(nèi)空氣濕度大,導(dǎo)致子房玻璃化,而增加瓊脂濃度,能夠降低培養(yǎng)基的水勢(shì),減少培養(yǎng)基中子房可獲得的水分,因而可明顯減輕子房的玻璃化現(xiàn)象[18]。但瓊脂濃度過(guò)高時(shí),會(huì)致使培養(yǎng)基太硬而不利于外植體吸收營(yíng)養(yǎng)[18],進(jìn)而導(dǎo)致胚狀體誘導(dǎo)率下降。
4 結(jié) 論
以不同的雜種F1代為試材,分別建立了煙草未授粉子房離體培養(yǎng)過(guò)程中的消毒方法、接種方式、接種密度、無(wú)機(jī)鹽濃度、外源激素配比和瓊脂濃度6個(gè)因素的最佳技術(shù)參數(shù)。在進(jìn)行煙草未授粉子房的離體培養(yǎng)過(guò)程中,花蕾不剝開(kāi)花瓣而直接采用0.1%升汞浸泡8 min進(jìn)行消毒,再剝開(kāi)子房壁將子房橫切后接種,每個(gè)125 mL三角瓶中接種2塊子房,培養(yǎng)基采用改良的H培養(yǎng)基額外增加60%無(wú)機(jī)鹽,附加IAA 0.5 mg/L和6-BA 2 mg/L,或者附加KT 2 mg/L和2,4-D 0.5 mg/L,瓊脂濃度9 g/L,能夠獲得較高的胚狀體誘導(dǎo)率。
[1] 陳海強(qiáng),劉會(huì)云,王軻,等. 植物單倍體誘導(dǎo)技術(shù)發(fā)展與創(chuàng)新[J]. 遺傳,2020,42(5):466-482.
CHEN H Q, LIU H Y, WANG K, et al. Development and innovation of haploid induction technologies in plants[J]. Hereditas (Beijing), 2020, 42(5): 466-482.
[2] 馮紅玉,姚碧嬌,陳媚,等. 單倍體育種技術(shù)的應(yīng)用進(jìn)展[J]. 中國(guó)農(nóng)學(xué)通報(bào),2021,37(30):1-6.
FENG H Y, YAO B J, CHEN M, et al. Haploid breeding techniques research progress[J]. Chinese Agricultural Science Bulletin, 2021, 37(30): 1-6.
[3] MUNA A, CHEN L, CAO J L, et al. An optimized culture system of haploid plants from unfertilized ovules of male-sterile tobacco[J]. Asian Journal of Plant Science and Research, 2022, 12(7): 303. DOI:10.36648/2249-7412.12.7.303.
[4] WERNSMAN E A, MATZINGER D F, RUFTY R C. Androgenetic vs. gynogenetic doubled haploids of tobacco[J]. Crop Science, 1989, 29(5): 1151-1155.
[5] 任學(xué)良,李繼新,李明海. 美國(guó)煙草育種進(jìn)展簡(jiǎn)況[J]. 中國(guó)煙草學(xué)報(bào),2007,13(6):57-64.
REN X L, LI J X, LI M H. Recent advances in tobacco breeding in the US[J]. Acta Tabacaria Sinica, 2007, 13(6): 57-64.
[6] 王文和. 未授粉子房和胚珠離體培養(yǎng)誘導(dǎo)植物雌核發(fā)育研究進(jìn)展[J]. 植物學(xué)通報(bào),2005,22(S1):108-117.
WANG W H. Advances in induction of gynogenesisby unpollinated ovary and ovule culture[J]. Chinese Bulletin of Botany, 2005, 22(S1): 108-117.
[7] 王文和,吳祿平,趙玉萍. 草莓未受精子房的離體培養(yǎng)[J]. 中國(guó)農(nóng)學(xué)通報(bào),2011,27(13):177-182.
WANG W H, WU L P, ZHAO Y P. Cultureof unfertilized ovary in strawberry (.)[J]. Chinese Agricultural Science Bulletin, 2011, 27(13): 177-182.
[8] BURK L G, GERSTEL D U, WERNSMAN E A. Maternal haploids ofLfrom seed[J]. Science, 1979, 206(4418): 585.
[9] 祝仲純,吳海珊. 從未授粉的小麥及煙草子房培養(yǎng)出單倍體植株[J]. 遺傳學(xué)報(bào),1979,6(2):181-183.
ZHU Z C, WU H S.production of haploid plantlets from the unpollinated ovaries ofand[J]. Acta Genetica Sinica, 1979, 6(2): 181-183.
[10] 祝仲純,吳海珊. 從煙草單倍體植株未傳粉子房又培養(yǎng)出單倍體植株[J]. 遺傳學(xué)報(bào),1981,8(1):63-65.
ZHU Z C, WU H S. Induction of haploid plantlets from unpollinated ovaries ofcultured[J]. Acta Genetica Sinica, 1981, 8(1): 63-65.
[11] 祝仲純,吳海珊,安慶坤. 大葉黃煙未傳粉子房培養(yǎng)及其細(xì)胞學(xué)研究[J]. 遺傳學(xué)報(bào),1984,11(4):281-287.
ZHU Z C, WU H S, AN Q K. Study on the culture of unpollinated ovaries ofand its cytology[J]. Acta Genetica Sinica, 1984, 11(4): 281-287.
[12] 吳伯驥,鄭國(guó)锠. 從未授粉煙草子房誘導(dǎo)單倍體植株的細(xì)胞學(xué)和胚胎學(xué)研究[J]. 植物學(xué)報(bào),1982,24(2):125-129.
WU B J, ZHENG G C. Cytological and embryological studies on haploid plant production from cultured unpollinated ovaries ofL.[J]. Acta Botanica Sinica, 1982, 24(2): 125-129.
[13] 潘莉,劉宗才,喜進(jìn)安. 外源激素對(duì)煙草未授粉子房培養(yǎng)的作用[J]. 河南農(nóng)業(yè)大學(xué)學(xué)報(bào),1998,32(2):167-170,175.
PAN L, LIU Z C, XI J A. The effects of exogenous hormone on cultured unpollinated ovaries ofL.[J]. Acta Agriculturae Universitatis Henanensis, 1998, 32(2): 167-170, 175.
[14] 潘莉,鄭奇君. 煙草未授粉子房單倍體誘導(dǎo)及影響因素的研究[J]. 河南農(nóng)業(yè)大學(xué)學(xué)報(bào),1999,33(1): 48-52.
PAN L, ZHENG Q J. Pathways and factors affecting haploid plant production from unpollinated ovaries ofL.[J]. Acta Agriculturae Universitatis Henanensis, 1999, 33(1): 48-52.
[15] 鄒甜,李涵,閔子揚(yáng),等. 蔬菜類植物組織培養(yǎng)過(guò)程中的褐變問(wèn)題及控制措施[J]. 長(zhǎng)江蔬菜,2015(8):1-4.
ZOU T, LI H, MIN Z Y, et al. Browning in plant tissue culture of vegetables and related control measures[J]. Journal of Changjiang Vegetables, 2015 (8): 1-4.
[16] 許智宏,SUNDERLAND N. 大麥花藥培養(yǎng)中的密度效應(yīng)[J]. 中國(guó)科學(xué)(B輯),1982(9):807-814.
XU Z H, SUNDERLAND N. Density effect in anther culture of barley[J]. Science in China (Series B), 1982(9): 807-814.
[17] 韓磊,汪旭東,吳先軍,等. 植物組織培養(yǎng)技術(shù)及其應(yīng)用研究進(jìn)展[J]. 種子,2005,24(1):38-43.
HAN L, WANG X D, WU X J, et al. Research progress of plant tissue culture technology and its application[J]. Seed, 2005, 24(1): 38-43.
[18] 李婭莉,張健,潘遠(yuǎn)智. 觀賞植物組織培養(yǎng)過(guò)程中玻璃化現(xiàn)象與解決措施進(jìn)展[J]. 四川農(nóng)業(yè)大學(xué)學(xué)報(bào),2004,22(3):278-282.
LI Y L, ZHANG J, PAN Y Z. Vitrification of shoots in tissue culture and advances in its prevention and cure[J]. Journal of Sichuan Agricultural University, 2004, 22(3): 278-282.
Study on the Factors Influencing the Induction of Embryoids byCulture of Unpollinated Ovary in Tobacco
CAO Jinglin, WANG Xin, CHENG Junqi*, LI Yapei, WU Chenglin, ZHANG Junjie, CAI Changchun
(Tobacco Research Institute of Hubei Province, Wuhan 430030, China)
The utilization value of female haploids derived from unpollinated ovaries is superior to male haploids derived from anthers or microspores in tobacco breeding. Therefore,culture of unpollinated ovary is of great significance for tobacco haploid breeding. In order to establish the technique system forculture of unpollinated ovaries, different F1 hybrids were used as test materials to explore the effects of six key factors, including disinfection method, inoculation method, inoculation density, inorganic salt concentration, exogenous hormone ratio, and agar concentration, on embryoid induction in HW media. The results showed that the measures that contributed to the induction of embryoids were as follows: the buds were immersed in 0.1% HG solution for 8 min without peeling the petals, the ovary walls were peeled off and the ovaries were transected before inoculation, 2 ovaries were inoculated in each 125 mL triangular flask, and the inorganic salt concentration in HW media was increased by an additional 60%, with additional IAA 0.5 mg/L and 6-BA 2 mg/L, or KT 2.0 mg/L and 2,4-D 0.5 mg/L, and agar concentration of 9 g/L. Based on this, the technical parameters forculture of unpollinated tobacco ovaries were further improved.
tobacco; unpollinated ovary;culture; embryoid; influencing factor
S572.01
A
1007-5119(2023)06-0084-07
中國(guó)煙草總公司湖北省公司科技重點(diǎn)項(xiàng)目(027Y2022-021)
曹景林(1967-),男,博士,副研究員,主要從事煙草育種研究。E-mail:caojinglin670425@sohu.com。*通信作者,E-mail:424653@qq.com
2023-07-06
2023-11-01
10.13496/j.issn.1007-5119.2023.06.012
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
中國(guó)煙草學(xué)報(bào)(2019年5期)2019-11-14 07:54:12
首都公共衛(wèi)生(2019年5期)2019-05-21 01:08:34
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
自動(dòng)化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學(xué)林(2013年2期)2013-11-12 12:58:54
中國(guó)煙草學(xué)報(bào)(2012年5期)2012-04-12 06:21:18
