擬南芥磷脂酸熒光探針的構(gòu)建及應(yīng)用
2024-02-25 12:18:26馬疏言鄭月萍鄭志富
浙江農(nóng)林大學(xué)學(xué)報 2024年1期
關(guān)鍵詞:植物
馬疏言,鄭月萍,2,鄭志富,2
(1.浙江農(nóng)林大學(xué) 現(xiàn)代農(nóng)學(xué)院,浙江 杭州 311300;2.浙江農(nóng)林大學(xué) 油料作物種質(zhì)創(chuàng)新與利用研究所,浙江 杭州 311300)
磷脂酸(phosphatidic acid,PA)是一種常見的細(xì)胞甘油磷脂,也是最簡單的膜脂之一[1]。幾乎所有生物體內(nèi)都會產(chǎn)生PA,包括酵母[2]、動物[3]和植物[4]等。在植物中PA 的合成主要包括2 種方式:第1 種是從頭合成途徑,3-磷酸甘油先后在3-磷酸甘油酰基轉(zhuǎn)移酶和溶血磷脂酸脂酰基轉(zhuǎn)移酶的作用下發(fā)生連續(xù)2 步酰化反應(yīng)生成PA,這也是PA 合成的主要途徑[5];第2 種是磷脂的降解途徑,磷脂降解又可分為2 種,一種是磷脂酶D 通過水解某些結(jié)構(gòu)磷脂直接生成PA[6-9],另一種是磷脂酶C 通過水解磷脂酰肌醇生成二脂酰甘油[10],繼而經(jīng)二脂酰甘油激酶磷酸化生成PA[11]。PA 作為一種結(jié)構(gòu)膜脂,磷酸基團(tuán)頭部能夠以“靜電/氫鍵開關(guān)模型”的方式與蛋白質(zhì)結(jié)合[12]。在該模型中,PA 結(jié)合蛋白中帶正電荷的堿性氨基酸殘基[13-17],可以吸引磷脂雙分子層上的負(fù)電荷,并通過靜電作用影響膜環(huán)境,繼而與PA 頭部的磷酸基團(tuán)形成氫鍵,產(chǎn)生一個穩(wěn)定的蛋白質(zhì)結(jié)合位點。PA 這種攜帶負(fù)電荷并且在生理條件下呈圓錐體的獨特分子構(gòu)型[18],使得PA 結(jié)合蛋白的結(jié)構(gòu)域能夠特異性的與其結(jié)合,保證了生物膜結(jié)構(gòu)的穩(wěn)定和功能的發(fā)揮。除了作為膜脂的一個結(jié)構(gòu)成分以及甘油脂合成的前體物質(zhì),PA 還是一個關(guān)鍵信號分子,在多種生物學(xué)過程中發(fā)揮重要作用。正常條件下,行使信號功能的PA 含量甚微;在植物應(yīng)答各種生物與非生物脅迫過程中PA 水平則會迅速升高,但這種積累往往是短暫的。這說明生物體擁有一套嚴(yán)格控制信號分子PA 含量的機(jī)制,這對調(diào)節(jié)PA 信號傳遞強(qiáng)度、維持細(xì)胞物質(zhì)與能量代謝的動態(tài)平衡至關(guān)重要[19-22]。
盡管PA 擁有多種功能,但目前對于細(xì)胞內(nèi)PA 含量的動態(tài)變化仍知之甚少,這主要受PA 檢測手段的限制。過去主要采用諸如薄層層析、高效液相色譜和放射性同位素標(biāo)記等理化手段對其含量進(jìn)行測定[23]。近年來,質(zhì)譜分析技術(shù)的應(yīng)用使得對包括PA 在內(nèi)的各種脂質(zhì)的測定更準(zhǔn)確和高效[24-25]。然而,這些理化手段都只能對組織器官中PA 總量與種類進(jìn)行定量或定性分析,無法捕捉細(xì)胞內(nèi)PA 的動態(tài)與瞬時變化信息。通常認(rèn)為,內(nèi)質(zhì)網(wǎng)上的PA 含量相對較高,這有助于促進(jìn)磷脂和三酰甘油的合成[26-27];而質(zhì)膜和細(xì)胞器中行使信號功能的PA 含量甚微。顯然,采用理化測定會掩蓋胞內(nèi)PA 的真實變化。因此,亟需開發(fā)一種能可視化分析細(xì)胞內(nèi)PA 的工具。
大量研究表明:生物體中PA 結(jié)合蛋白種類繁多,這些蛋白中的PA 結(jié)合域(phosphatidic acid binding domains,PABD)結(jié)構(gòu)亦存在差異。其中一些PABD 因與PA 結(jié)合專一性強(qiáng),被用來開發(fā)多種PA 探針[28-33]。酵母蛋白Spo20p 中的PA 結(jié)合域已被證實具有極強(qiáng)的專一性[33],只結(jié)合PA 而不結(jié)合其他磷脂。目前基于這一PABD 所開發(fā)的PA 探針在醫(yī)學(xué)研究中應(yīng)用廣泛,但在植物中的應(yīng)用卻有限[34]。因此,本研究旨在構(gòu)建基于Spo20p 中的PABD 的植物PA 熒光探針,為剖析植物早期應(yīng)答逆境脅迫的細(xì)胞分子機(jī)制提供新工具。
1 材料與方法
1.1 植株培養(yǎng)
1.1.1 土壤基質(zhì)的培養(yǎng) 將擬南芥Arabidopsisthaliana種子置于含有濕潤濾紙的培養(yǎng)皿中,4 ℃冰箱內(nèi)避光放置3 d 后將其點播在濕潤的土壤基質(zhì)上(V營養(yǎng)土∶V蛭石∶V珍珠巖=3∶1∶1)。將播種后的盆栽用透明塑料蓋覆蓋,置于24~25 ℃室溫、50%~60%濕度、14 h 光照/10 h 黑暗的植物培養(yǎng)箱中培養(yǎng),7 d 后揭去塑料蓋間苗。
1.1.2 培養(yǎng)基的培養(yǎng) 將擬南芥種子進(jìn)行消毒,加入適量滅菌過的蒸餾水,在4 ℃冰箱內(nèi)避光放置3 d,點播在1/2 MS 培養(yǎng)基上,置于植物培養(yǎng)間中培養(yǎng)。
1.2 PA 熒光探針的構(gòu)建
1.2.1 pSY06-GFP-PABD載體的構(gòu)建 酵母蛋白Spo20p 中存在一個高度專一的PA 結(jié)構(gòu)域(PABD),該PABD 由40 個氨基酸殘基構(gòu)成,只結(jié)合PA 而不結(jié)合其他磷脂[33-34],因此,本研究選擇該PABD 與熒光蛋白融合,構(gòu)建PA 熒光探針。根據(jù)PAPD 核苷酸序列[33]設(shè)計引物(表1)進(jìn)行PCR 擴(kuò)增,獲得與該PABD 相對應(yīng)的DNA 片段;同時PCR 擴(kuò)增獲得GFP基因編碼區(qū)序列。隨后,利用無縫克隆試劑盒(生工),將GFP和PABD片段克隆至植物表達(dá)載體pSY06 的限制性內(nèi)切酶位點KpnI 和PstI 之間,獲得重組質(zhì)粒pSY06-GFP-PABD。PABD連接至GFP基因編碼區(qū)的3′端,構(gòu)成GFP-PABD融合基因,該融合基因的表達(dá)由組成型啟動子UBQ10 驅(qū)動。重組質(zhì)粒中的插入片段經(jīng)菌落PCR、KpnI 和PstI 雙酶切以及測序驗證正確后,采用熱激法將質(zhì)粒轉(zhuǎn)化至農(nóng)桿菌AgrobacteriumtumefaciensGV3101,用于后續(xù)擬南芥遺傳轉(zhuǎn)化。

表1 相關(guān)引物序列Table 1 Sequence of related primers
1.2.2 擬南芥的遺傳轉(zhuǎn)化、篩選及鑒定 以哥倫比亞野生型擬南芥(WT)為植物材料,利用農(nóng)桿菌花序侵染法轉(zhuǎn)化擬南芥。將獲得的T1代擬南芥種子進(jìn)行表面消毒,均勻鋪在含有草銨膦(10 mg·L-1)和特美汀(50 mg·L-1)的1/2 MS 篩選培養(yǎng)基上進(jìn)行篩選。將具有除草劑抗性的陽性苗移栽到土壤中繼續(xù)培養(yǎng),3 周后提取葉片DNA 并設(shè)計引物(表1)進(jìn)行PCR 鑒定,對T2和T3代轉(zhuǎn)基因株系進(jìn)行除草劑抗性的遺傳分析,最終篩選得到純合單插入位點轉(zhuǎn)基因株系。
1.3 轉(zhuǎn)基因擬南芥中目的基因表達(dá)量分析
以在1/2 MS 培養(yǎng)基中生長7 d 的轉(zhuǎn)基因擬南芥根系作為測定材料,使用高純度RNA 提取試劑盒(全式金)提取RNA 并對其進(jìn)行純度檢測和濃度測定。取等量RNA,使用反轉(zhuǎn)錄試劑盒(翌圣)進(jìn)行單鏈cDNA 的合成。選擇擬南芥肌動蛋白基因Actin為內(nèi)參基因,設(shè)計引物(表1)進(jìn)行實時熒光定量PCR(RT-qPCR),并對數(shù)據(jù)采用2-ΔΔCt法進(jìn)行分析。
1.4 PA 熒光探針轉(zhuǎn)基因擬南芥的表型觀察
將野生型和轉(zhuǎn)基因株系種子點播在1/2 MS 方形培養(yǎng)基上,豎直培養(yǎng)6 d 后,挑選長勢均勻一致的幼苗(每個基因型,20 株),分別轉(zhuǎn)移至含有0 和50 mmol·L-1NaCl 的1/2 MS 固體培養(yǎng)基中繼續(xù)豎直培養(yǎng),7 d 后觀察表型。
1.5 PA 熒光探針靈敏度分析
將在1/2 MS 方形培養(yǎng)基上豎直培養(yǎng)6 d 的幼苗分別浸泡在含有0、2 和10 μmol·L-1PA 的1/2 MS 液體培養(yǎng)基中,依次處理5、10、15 和20 min 后,利用熒光顯微鏡(ZEISS,Axio Imager 2)對擬南芥根尖分生區(qū)PA 的分布進(jìn)行觀察并拍照。
1.6 鹽堿脅迫下根尖PA 的檢測
將含PA 熒光探針的擬南芥轉(zhuǎn)基因株系種子點播在1/2 MS 方形培養(yǎng)基上,豎直培養(yǎng)6 d,選擇均勻一致的幼苗分別浸泡在含有100 mmol·L-1NaCl、10 mmol·L-1NaHCO3和兼含兩者的1/2 MS 液體培養(yǎng)基中,處理5、10 和15 min 后用熒光顯微鏡觀察根尖分生區(qū)PA 的分布。
2 結(jié)果與分析
2.1 PA 熒光探針的設(shè)計及其轉(zhuǎn)基因擬南芥的獲得
如圖1 所示:該載體以pSY06 為骨架,由35S 啟動子驅(qū)動植物選擇標(biāo)記基因——草銨膦抗性基因(bar)的表達(dá),而GFP-PABD融合基因的表達(dá)則由UBQ10 啟動子驅(qū)動。

圖1 pSY06-GFP-PABD 載體圖Figure 1 Vector diagram of pSY06-GFP-PABD
采用農(nóng)桿菌花序侵染法將GFP-PABD融合基因轉(zhuǎn)入擬南芥,獲得T1代轉(zhuǎn)基因種子。對T1代幼苗進(jìn)行草銨膦(10 mg·L-1)抗性篩選與PCR 鑒定,獲得56 個含融合基因的獨立轉(zhuǎn)基因株系。隨后,對相應(yīng)株系的T2代幼苗(每個株系約300 株)繼續(xù)進(jìn)行草銨膦除草劑抗性篩選。卡方測驗顯示:符合3(陽性苗數(shù))∶1(陰性苗數(shù))分離規(guī)律的株系共8 個,這些株系為含單插入位點的轉(zhuǎn)基因株系。再對T2代各個株系不同植株的自交后代(T3代)進(jìn)行除草劑抗性篩選,最終獲得7 個純合、單插入位點轉(zhuǎn)基因擬南芥株系,分別為8-1、11-1、13-12、25-5、42-4、48-1 和53-1。
2.2 不同轉(zhuǎn)基因株系PA 熒光探針表達(dá)量的分析
為了明確不同轉(zhuǎn)基因株系PA 熒光探針表達(dá)量的差異,對上述7 個轉(zhuǎn)基因株系進(jìn)行了目的基因的表達(dá)量測定。RT-qPCR 結(jié)果(圖2)顯示:當(dāng)將轉(zhuǎn)基因株系8-1 中融合基因的相對表達(dá)量設(shè)為參考值“1”,其余6 個株系(11-1、13-12、25-5、42-4、48-1 和53-1)的相對表達(dá)量分別為1.74、15.91、0.72、1.06、1.69 和1.52。株系13-12 的PA 探針的表達(dá)量最高,是其他株系的9~22 倍,為PA 探針高表達(dá)轉(zhuǎn)基因株系;相應(yīng)地,將株系48-1 作為PA 探針低表達(dá)材料。

圖2 不同轉(zhuǎn)基因擬南芥株系中GFP-PABD 融合基因的相對表達(dá)量Figure 2 Relative expression level of the GFP-PABD fusion gene in transgenic A.thaliana lines
2.3 PA 熒光探針對擬南芥生長發(fā)育的影響
為了明確PA 熒光探針的表達(dá)是否會對植物生長發(fā)育產(chǎn)生影響,比較分析了野生型擬南芥和不同轉(zhuǎn)基因株系在整個生長發(fā)育周期的表型差異。土培生長實驗顯示:野生型與轉(zhuǎn)基因擬南芥株系的葉片生長無明顯差異,高表達(dá)轉(zhuǎn)基因株系(13-12)與低表達(dá)轉(zhuǎn)基因株系(48-1)之間亦無可見表型差異(圖3A)。另外,在含有0 和50 mmol·L-1NaCl 的1/2 MS 培養(yǎng)基中生長時,野生型與不同轉(zhuǎn)基因株系之間也未表現(xiàn)出生長速率的差異(圖3B)。這些結(jié)果說明:在一定范圍內(nèi)PA 熒光探針的表達(dá)不會影響擬南芥的正常生長發(fā)育,這一特性有助于合理解釋特定條件下胞內(nèi)PA 變化對植物生長發(fā)育產(chǎn)生的影響。

圖3 野生型擬南芥與3 個PA 熒光探針轉(zhuǎn)基因株系的表型比較Figure 3 Phenotypic comparison between wild type and three transgenic lines of A.thaliana bearing PA fluorescent probe
2.4 熒光探針監(jiān)測PA 的靈敏度
隨著熒光探針表達(dá)量的增加,細(xì)胞內(nèi)的熒光強(qiáng)度也會增強(qiáng),這會導(dǎo)致PA 檢測的信噪比下降;相反,熒光探針表達(dá)量過低則會影響PA 檢測的靈敏度。為了獲得檢測靈敏度和信噪比都較高的PA 熒光探針,用不同濃度的外源PA 處理熒光探針表達(dá)量存在差異的轉(zhuǎn)基因株系,進(jìn)而在熒光顯微鏡下觀察根尖PA 的積累情況。結(jié)果(圖4~6)顯示:野生型擬南芥在有、無外源PA 處理下,根尖細(xì)胞自發(fā)熒光的強(qiáng)度無明顯差別。另外,在不同濃度外源PA 處理下,熒光探針低表達(dá)的轉(zhuǎn)基因株系(48-1)的根尖分生區(qū)的PA 含量變化未能被捕捉到。相反,對于熒光探針高表達(dá)的轉(zhuǎn)基因株系(13-12),2 和10 μmol·L-1PA 分別處理10 和5 min 后,其根尖分生區(qū)細(xì)胞的胞內(nèi)或質(zhì)膜上的PA 積累清晰可見(圖5)。當(dāng)外源PA 濃度低時,隨著處理時間延長,細(xì)胞中PA 積累減少(圖5),這很可能是因為作為甘油脂合成前體物質(zhì)的PA 已被轉(zhuǎn)化為其他物質(zhì)。而當(dāng)外源PA 濃度升至10 μmol·L-1時,處理10、20 min 后依然可見細(xì)胞中的PA 呈點狀分布(圖6)。這些結(jié)果說明:熒光探針表達(dá)量較高的轉(zhuǎn)基因株系(13-12)可被用于監(jiān)測細(xì)胞中PA 含量的變化。
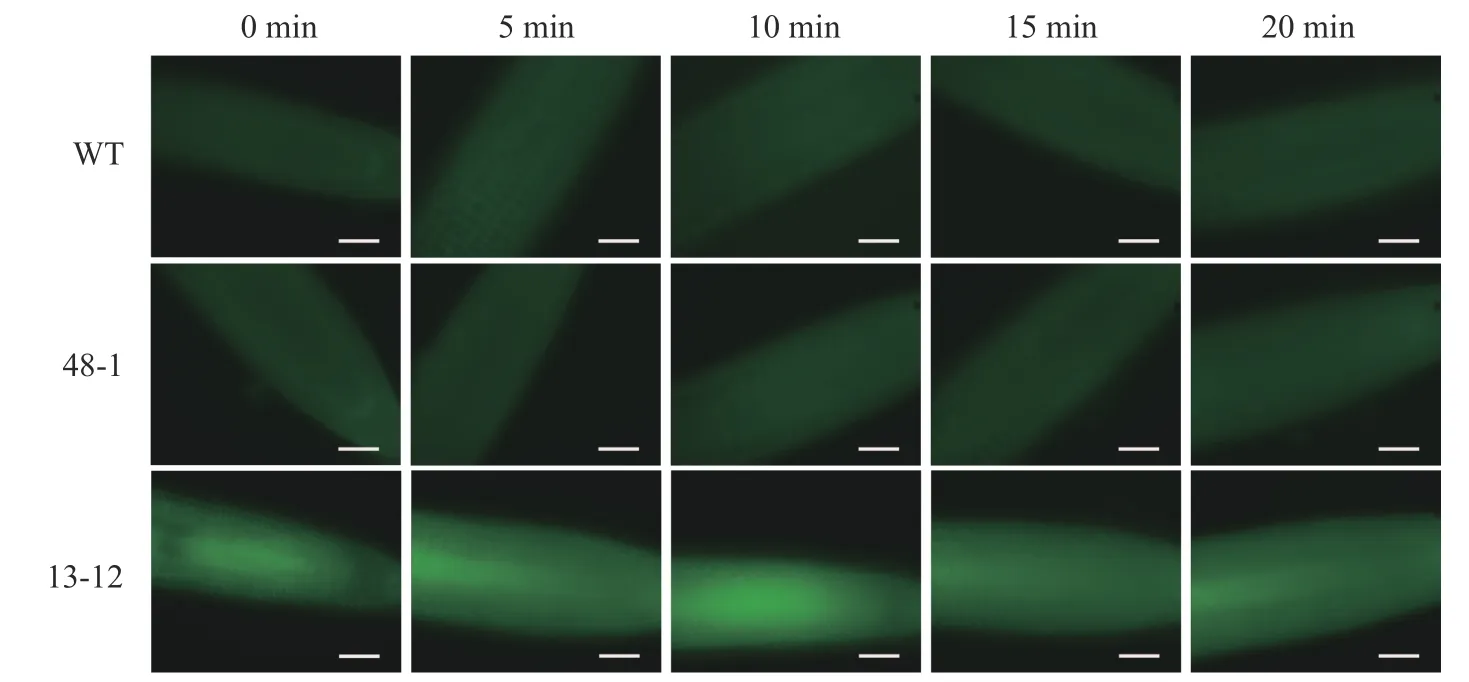
圖4 無外源PA 處理下PA 探針在野生型和2 個轉(zhuǎn)基因擬南芥株系根尖中熒光強(qiáng)度的差異分析Figure 4 Analysis of discrepancy in fluorescent intensity of PA probe between wild type and two transgenic lines of A.thaliana

圖5 外源PA(2 μmol·L-1)處理下PA 熒光探針對PA 監(jiān)測靈敏度的分析Figure 5 Sensitivity analysis of PA fluorescent probe monitoring PA under treatment with 2 μmol·L-1 exogenous PA

圖6 外源PA (10 μmol·L-1)處理下PA 熒光探針對PA 監(jiān)測靈敏度的分析Figure 6 Sensitivity analysis of PA fluorescent probe monitoring PA under treatment with 10 μmol·L-1 exogenous PA
2.5 熒光探針檢測鹽堿脅迫下擬南芥根尖PA 的含量變化
為了進(jìn)一步驗證 PA 熒光探針的實用性,利用熒光探針表達(dá)量較高的轉(zhuǎn)基因株系(13-12)檢測鹽堿脅迫條件下根尖細(xì)胞中PA 含量的變化。結(jié)果顯示:鹽、堿單獨或混合處理轉(zhuǎn)基因株系,5 min 后即可在根尖分生區(qū)細(xì)胞中觀察到PA 的點狀分布(圖7)。因此推斷,本研究構(gòu)建的PA 熒光探針可用于監(jiān)測逆境條件下植物細(xì)胞中PA 含量的變化。另外,鹽堿脅迫可快速誘導(dǎo)PA 積累這一現(xiàn)象表明在植物早期逆境響應(yīng)中PA 發(fā)揮某種重要作用。

圖7 單獨或混合鹽堿脅迫下PA 在轉(zhuǎn)基因擬南芥根尖分生組織中的分布Figure 7 Distribution of PA in the root tip meristem of a transgenic A.thaliana line bearing PA sensor under saline and alkaline stresses, alone or in combination
3 討論
已知PA 在植物生長發(fā)育與逆境響應(yīng)過程中發(fā)揮重要作用,但由于缺乏PA 的可視化分析工具,對細(xì)胞中PA 含量動態(tài)變化的了解十分有限。這一方面限制了PA 作用機(jī)制的研究,另一方面給評估基因工程手段操控植物細(xì)胞PA 含量變化所產(chǎn)生的實際應(yīng)用效果帶來困難。因此,本研究將酵母蛋白Spo20p 中對PA 高度專一的結(jié)合域與熒光蛋白融合,成功構(gòu)建了可對細(xì)胞內(nèi)PA 進(jìn)行可視化檢測的熒光探針。
為了使熒光探針適用于監(jiān)測整個生長周期不同組織細(xì)胞中的PA 水平,選擇組成型表達(dá)啟動子UBQ10 驅(qū)動GFP-PABD融合基因的表達(dá)。研究發(fā)現(xiàn):植物細(xì)胞中PA 的檢測靈敏度與PA 熒光探針的表達(dá)量密切相關(guān),表達(dá)量較高的熒光探針可有效檢測到外源PA 處理帶來的胞內(nèi)PA 水平的變化,而表達(dá)量較低的探針檢測效果則相反。如13-12 株系中PA 熒光探針的表達(dá)量高于其他株系,該株系經(jīng)2 μmol·L-1外源PA 處理10 min,其根尖細(xì)胞中PA 的點狀分布即可被觀察到,這也說明外源PA 能夠被擬南芥根尖快速吸收,繼而與PA 熒光探針結(jié)合。鑒于已有研究中常用濃度高于10 μmol·L-1的外源PA 檢測PA 探針的靈敏度[35],本研究中構(gòu)建的熒光探針可有效監(jiān)測到2 μmol·L-1外源PA 處理所帶來的細(xì)胞內(nèi)PA 含量的變化,推測基于13-12 轉(zhuǎn)基因株系的PA 熒光探針具有較高的PA 檢測靈敏度。
需要指出的是,PA 熒光探針使用過程中,樣品處理時間的延長和觀察光源的增強(qiáng)均會使植物細(xì)胞造成某種損傷,導(dǎo)致細(xì)胞產(chǎn)生自發(fā)熒光,從而降低PA 檢測的信噪比。因此在實驗過程中,需盡量縮短植物樣本的制片時間及熒光觀察的時間,以提高觀察結(jié)果的真實性。
鹽堿地中鹽和堿是2 種共存但不同的非生物脅迫,共同影響植物的正常生長發(fā)育。利用本研究構(gòu)建的PA 熒光探針發(fā)現(xiàn):鹽脅迫處理5 min 就會造成PA 在細(xì)胞中的積累,這與已有的研究結(jié)果一致[22,36]。同時,堿脅迫和鹽堿混合脅迫也會快速誘導(dǎo)PA 的積累。因此,為了全面了解植物早期逆境響應(yīng)機(jī)制,有必要監(jiān)測早期細(xì)胞中PA 含量的動態(tài)變化。本研究開發(fā)的PA 熒光探針為這些方面的研究提供了重要技術(shù)支撐。
4 結(jié)論
本研究成功構(gòu)建了一種基于酵母蛋白Spo20p 中PA 結(jié)合域的熒光探針,可有效監(jiān)測植物細(xì)胞中PA 含量變化。應(yīng)用該探針發(fā)現(xiàn)鹽堿脅迫可快速誘導(dǎo)擬南芥根尖細(xì)胞質(zhì)膜上和胞內(nèi)PA 的積累,這一結(jié)果表明PA 在植物對鹽堿脅迫早期應(yīng)答過程中發(fā)揮重要作用。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
