
嗜熱鏈球菌c-di-AMP 合成酶的結構預測
2024-02-29 07:36:52范藝周駱思羽朱浩偉艾連中熊智強
工業微生物 2024年1期
關鍵詞:結構
范藝周,駱思羽,朱浩偉,薛 珍,艾連中,熊智強
上海理工大學健康科學與工程學院,上海食品微生物工程技術研究中心,上海 200093
嗜熱鏈球菌(Streptococcus thermophilus)是一種革蘭氏陽性菌,也是一種公認的食品安全菌株,被廣泛應用于酸奶和奶酪等傳統乳制品的生產過程中[1-2]。為適應乳制品發酵過程中的酸性環境,感知細胞外的信號并用信使分子來傳遞信號,對嗜熱鏈球菌的生存尤為重要。第二信使分子c-di-AMP 廣泛存在于細菌、古生菌尤其是厚壁菌門中,與脂肪酸代謝、鉀離子穩態、滲透壓調節、細胞毒力以及生物膜形成等多種生理功能密切相關[3-8]。為闡明c-di-AMP 在嗜熱鏈球菌中的調控機制,研究c-di-AMP合成酶的結構與功能非常有必要。細菌c-di-AMP合成酶DAC 包含一個高度保守的DAC/DisA_N 結構域,可催化ATP 或ADP 合成c-di-AMP。DAC 蛋白活性與保守的DGA(Asp-Gly-Ala)和RHR(Arg-His-Arg)基序密切相關[9-12]。DAC 催化c-di-AMP 合成需要輔因子二價金屬離子,用DAC 三級結構可以預測可能的輔因子和關鍵結合位點[13]。
本實驗室在實驗前期在傳統乳制品中篩選出了一株高產胞外多糖的嗜熱鏈球菌菌株S-3,并對其進行了全基因組測序[14]。結合HMMER 預測和NCBI數據庫比對在嗜熱鏈球菌S-3 基因組中預測出的c-di-AMP 合成酶(命名為StDAC),對其跨膜區和二級結構進行預測,并通過SWISS-MODEL 和Modeller 進行三級結構模擬,為后續探究StDAC 的結構與功能奠定基礎。
1 方法
1.1 二級結構預測
將StDAC 氨基酸序列提交至蛋白質序列網絡分析在線軟件(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.plpage=/NPSA/npsa_server.html),選擇其中9 個有代表性的二級結構預測方法:SOPM,HNN,MLRC,DPM,DSC,GOR Ⅰ,GOR Ⅲ,PHD,Predator 對StDAC 二級結構進行預測[15-16]。其中HNN,MLRC,DSC,GORⅢ,PHD,Predator 為三態定義法,“h”代表α-螺旋、“e”代表β-折疊、“c”代表無規則卷曲;SOPM,DPM 和GORⅠ為四態定義法,“h”代表α-螺旋、“e”代表β-折疊、“c”代表無規則卷曲、“t”代表β-轉角。為統一標準采用三態定義法對二級結構進行比較分析。
1.2 SWISS-MODEL 單模板建模
將StDAC 氨基酸序列提交至SWISS-MODEL蛋白分子建模服務器(https://swissmodel.expasy.org/)進行單模板建模,返回GMQE 值小于1 且接近1、QMENA 值接近0、氨基酸同源性大于30%模板的pdb 格式。建立的模型通過Discovery studio 可視化即可模擬StDAC 三級結構[17]。建模模板來自于美國國家生物技術信息中心(National Center of Biotechnology Information,NCBI)的Structure 數據庫。
1.3 Modeller 多模板建模
將StDAC 氨基酸序列提交至Blast(https://blast.ncbi.nlm.nih.gov/Blast.cgi)、Phyre2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id =index)和HHpred(https://toolkit.tuebingen.mpg.de/tools/hhpred)在線數據庫,搜索與StDAC 具有同源性的模板。將氨基酸同源性與軟件評分較高的單模板依次通過腳本salign.py,align2d_mult.py 和model_mult.py 進行多模板建模,返回的pdb 文件通過Discovery studio可視化即可模擬StDAC 三級結構。
1.4 模型評估
將預測的模型提交至SAVES 在線服務器(https://saves.mbi.ucla.edu/), 根據提示分別對Verify-3D,Errat 和PROCHECK 中Ramachandran 圖對模型的3D 結構、晶體學和生理化學參數進行評估。
2 結果分析與討論
2.1 c-di-AMP 合成酶預測
利用HMMER 預測和NCBI 數據庫比對在嗜熱鏈球菌S-3 全基因組中發現的一個編碼DisA_N 結構域的c-di-AMP 合成酶基因(orf0696),將其編碼的蛋白命名為StDAC。StDAC 分別與已報道的枯草芽孢桿菌c-di-AMP 合成酶CdaA 和DisA 擁有47%和33%的氨基酸序列相似性[10,18]。用PROTTER在線分析軟件對StDAC 跨膜區進行預測,結果顯示StDAC 的第7~52 個氨基酸為跨膜區,第1~6 個氨基酸和第53~235 個氨基酸位于膜內側。跨膜區預測結果為StDAC 異源表達提供了借鑒,可通過切除第7~52 個氨基酸提高可溶性蛋白含量。如圖1 所示。
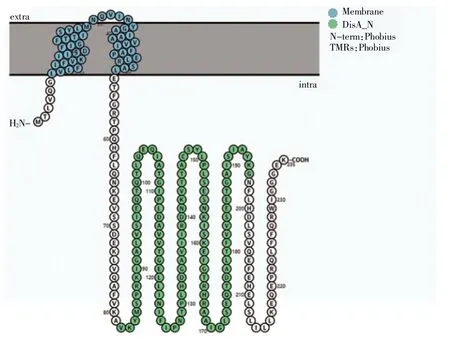
圖1 PROTTER 預測StDAC 跨膜區
2.2 StDAC 二級結構預測
二級結構預測方法通常基于統計經驗算法(MLRC、DPM、GOR Ⅰ、GOR Ⅲ)、物理化學算法(SOPM、DSC、Predator)和神經網絡算法(HNN、PHD),這些方法預測的α-helix 的準確性高于90%[19-20]。綜合9 種預測方法得到的StDAC 二級結構有42.98%(共101 個氨基酸)為α-螺旋,15.32%(共36 個氨基酸)為β-折疊,38.30%(共90 個氨基酸)(圖2)為無規則卷曲。用物理化學算法DSC 和神經網絡算法PHD 預測的二級結構更接近綜合預測結果。如表1所示。
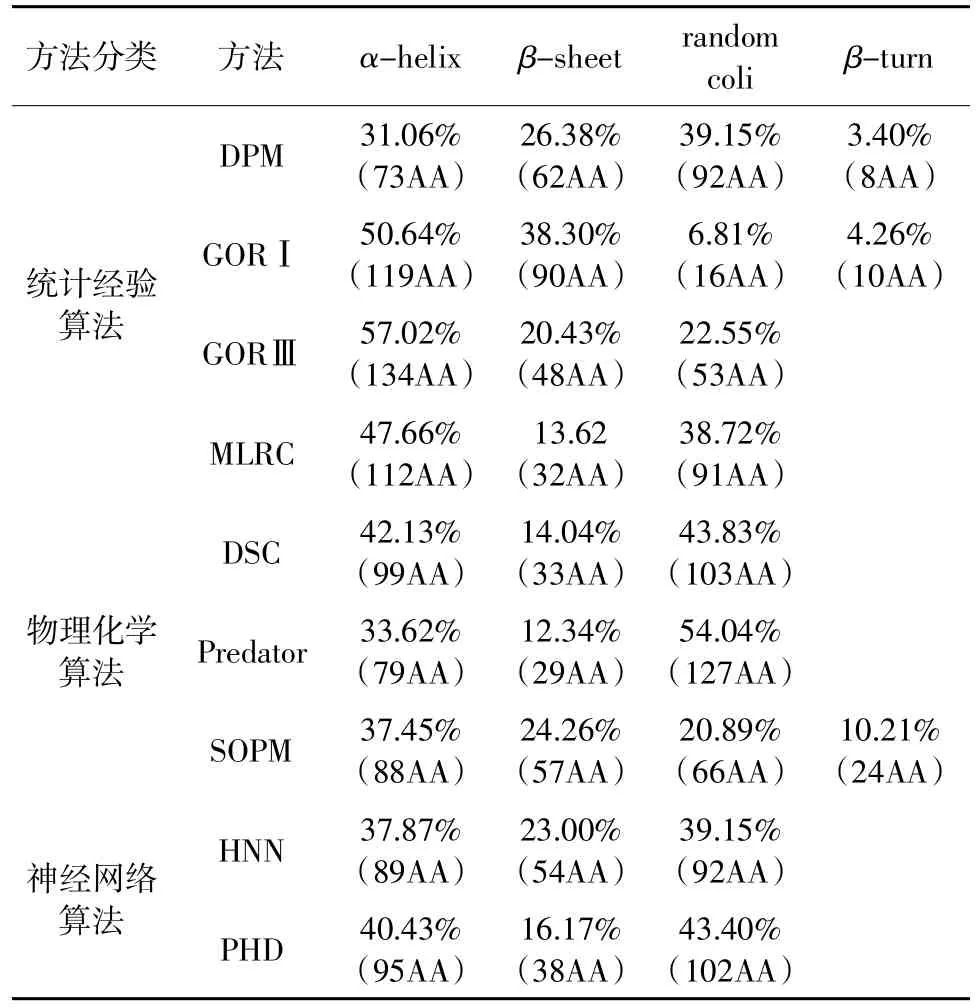
表1 StDAC 不同算法二級結構預測結果
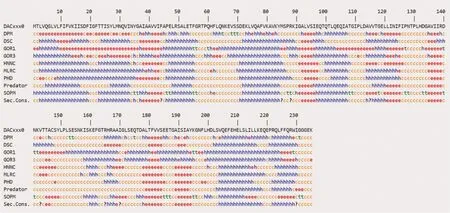
圖2 StDAC 二級結構預測
2.3 StDAC 三級結構建模
SWISS-MODEL 在線服務器選擇用氨基酸同源性最高和覆蓋率最廣的模板6hvm 對StDAC 進行單模板建模(圖3A)。由于模板6hvm 的晶體結構沒有跨膜區,因此SWISS-MODEL 要對含DisA_N 結構域的StDAC 從第70~228 個氨基酸序列(共158 個氨基酸)進行三維建模。利用Modeller 進行多模板建模,用NCBI、Phyre2 和HHpred 在線數據庫搜索StDAC 建模模板,以氨基酸同源性高、軟件評分高和氨基酸覆蓋率廣為標準,以6gyw、6huw 和d2fb5a1為模板對StDAC 進行多模板建模,如圖3B 所示。用SWISS-MODEL 和Modeller 兩種方法建立的三級結構都包含5 個α-螺旋區域和7 個β-折疊區域,但Modeller 建立的三級結構包括跨膜區域。
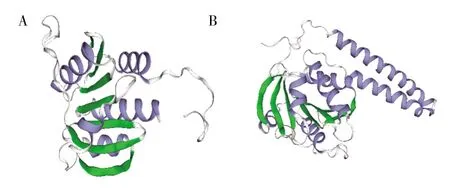
圖3 StDAC 的SWISS-MODEL 和Modeller 建模A:SWISS-MODEL;B:Modeller
2.4 StDAC 三級結構評估
Ramachandran 圖為最常見三級結構評估方法,圖中的紅色區域為“最適區域”,黃色區域為“允許區域”,其他區域的紅點均為構象不合理的氨基酸,合理的三級結構須有90%以上的氨基酸落在紅色區域[21]。用SWISSMODEL 建立的StDAC 三級結構有92.0%的氨基酸落在最適區域,8.0%的氨基酸落在允許區域。用Modeller 建立的三級結構有92.4%的氨基酸落在最適區域,7.6%的氨基酸落在允許區域。因此, 兩種方法建立的三級結構在Ramachandran 圖評估中都不存在氨基酸構象不合理的情況。如圖4 所示。
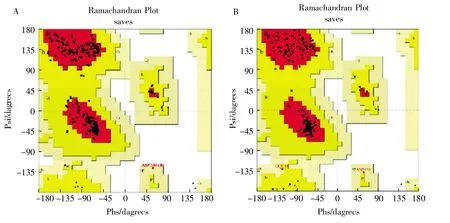
圖4 StDAC 三級結構Ramachandran 圖A:SWISS-MODEL;B:Modeller
Errat 通過統計和分析0.35 nm 半徑內,不同類型原子之間非鍵相互作用的數據,是通過晶體學方法評估蛋白質模型,軟件評分大于85%的模型才具有可靠性,圖中的黃線和紅線越多表示三級結構的可靠性越差[22]。由SWISSMODEL 建立的StDAC 三級結構Errat 的評分為98.98%,模型通過評估。而Modeller 建立的三級結構Errat 的評分為78.28%,其中跨膜區和第180~230 個氨基酸處出現了較多的黃線和紅線,構象不合理導致模型評分低,如圖5所示。
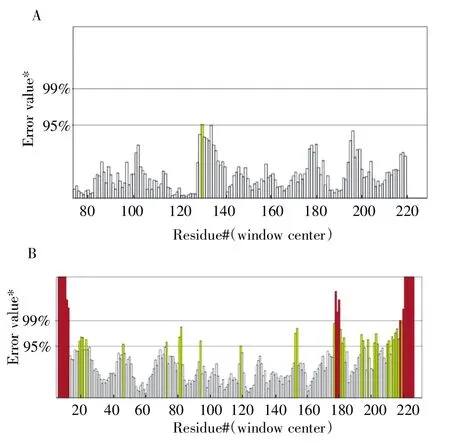
圖5 StDAC 三級結構Errat 評分A:SWISS-MODEL;B:Modeller
Verify-3D 是基于統計分析結果來評估蛋白三維結構與氨基酸序列的相容性的,合理的三級結構通常有80%以上氨基酸的相容性評分高于0.2[23]。由SWISSMODEL建立的StDACVerify-3D評分為84.06%,通過評估。而Modeller 建立的StDAC 三級結構Verify-3D 的評分為54.04%,沒有通過評估,與Errat的評估結果相同,跨膜區和第180~230 個氨基酸結構不合理導致整體結構評分較低,如圖6 所示。
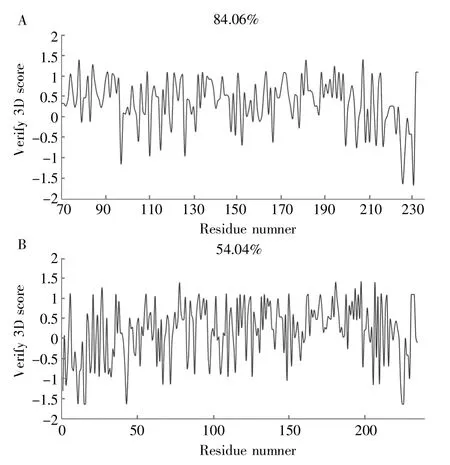
圖6 StDAC 三級結構Verify-3D 分析A:SWISS-MODEL;B:Modeller
通過Verify-3D、Errat 和PROCHECK 中的Ramachandran圖對建立的StDAC 三級結構進行評估,評估結果表明由SWISS-MODEL 建立的StDAC三級結構通過了Ramachandran、Errat 和Verify-3D的評估,蛋白結構較為合理;由Modeller 建立的全長StDAC 三級結構僅通過了Ramachandran 評估,三級結構評分低。因此,可通過SWISS-MODEL 模擬的三級結構對StDAC 的結構與功能進行研究。
3 結論
基于嗜熱鏈球菌S-3 全基因組測序結果,結合HMMER 預測和NCBI 數據庫比對篩選出c-di-AMP 合成酶StDAC。利用統計經驗算法、物理化學算法和神經網絡算法共9 種方法預測StDAC 的二級結構42.98%為α-螺旋、15.32%為β-折疊、38.30%為無規則卷曲。分別通過SWISS-MODEL 和Modeller 模擬StDAC 三級結構,其中SWISSMODEL 單模板建模的StDAC 三級結構在PROCHECK、Verify-3D 和Errat 三個分析軟件中評分最高,為研究c-di-AMP 合成酶StDAC 的結構與功能提供了可靠的模型,并為進一步闡明嗜熱鏈球菌中c-di-AMP 的調控機制奠定了基礎。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
