飼料混合植物油替代魚油對不同品種異育銀鯽生長性能和脂肪代謝的影響
2024-03-02 06:38:08李紅燕巫麗云金俊琰朱曉鳴劉昊昆楊云霞解綬啟
水生生物學報 2024年4期
李紅燕 巫麗云 金俊琰 韓 冬 朱曉鳴 劉昊昆 楊云霞 解綬啟
(1. 中國科學院水生生物研究所淡水生態與生物技術國家重點實驗室, 武漢 430072;2. 中國科學院大學現代農業科學學院, 北京 100049)
水產養殖為人類消費提供了近2/3的動物蛋白來源, 而水產養殖業的發展高度依賴于水產飼料業的發展。脂類是水產飼料中重要的營養素, 也是維持生物體細胞膜和亞細胞膜完整性的必需成分。脂肪作為激素的前體, 參與許多重要的生物過程,可以促進脂溶性維生素的吸收[1]。魚油因其富含必需長鏈多不飽和脂肪酸(LC-PUFAs)、二十碳五烯酸(EPA)和二十二碳六烯酸(DHA), 在魚體脂肪代謝和抗氧化等生理生化過程中發揮重要作用, 被認為是水產飼料中的優質脂肪源[2]。然而, 目前全球魚油產量可能不足以滿足未來水產養殖的需求, 導致了飼料成本的增加[3]。另外, 魚油的生產造成野生魚類資源和水資源環境的破壞, 阻礙了水產養殖業的快速發展。因此, 尋找適宜的飼料脂肪源來替代魚油, 對水產養殖可持續發展具有重要意義[4]。
與魚油相比, 植物油具備廉價易得、產量豐富及較易儲存等優勢, 具有替代魚油的潛能[5]。研究表明, 大豆油、棕櫚油、菜籽油和亞麻油等多種植物油, 已被用來部分或完全替代魚油而被廣泛應用于水產飼料中[6—8]。在大黃魚(Larimichthys crocea)中, 飼料亞麻籽油和豆油完全替代魚油, 魚體增重率和特定生長率顯著降低[6]。大豆油、菜籽油和亞麻油分別完全替代魚油導致軍曹魚(Rachycentron canadum)生長遲緩、腹部和肝臟脂肪沉積[7,8]。植物油替代對魚體造成不同的影響可能是不同脂肪源中脂肪酸組成差異引起的。因此, 通過多種植物油混合達到平衡的脂肪酸模式可能是提高植物脂肪源替代魚油效果的潛在方式。
異育銀鯽(Carassius gibelio)是我國重要的大宗淡水魚養殖品種, “中科3號”和“中科5號”是目前廣泛推廣的兩個重要品種。前期研究表明, 胰島素對不同品種異育銀鯽脂肪代謝的調控存在顯著差異[9]。此外, 不同品種異育銀鯽在飼料不同糖、脂水平下,對脂肪代謝響應存在品種差異性[10]。因此, 我們推測異育銀鯽“中科3號”和“中科5號”對飼料脂肪源的利用可能存在差異。本實驗以異育銀鯽“中科3號”和“中科5號”為研究對象, 參考魚油的脂肪酸組成配制混合植物油飼料, 評估不同飼料脂肪源對不同品種異育銀鯽生長性能、魚體基本組分、肌肉脂肪酸組成、血漿生化及脂肪代謝(脂肪合成、脂肪分解、脂肪酸β氧化和脂肪酸轉運)的影響。本研究可為植物脂肪源替代魚油提供數據支撐, 并為高效利用植物脂肪源的魚類遺傳選育提供理論基礎。
1 材料與方法
1.1 實驗飼料
本實驗以魚油為基礎參照, 將菜籽油、大豆油、棕櫚油按照55%∶25%∶20%的比例調配至與魚油相似的脂肪酸組成。分別以魚油(FO)和混合植物油(BVO)為脂肪源, 配制兩種等氮(35%)等脂(9%)飼料, 實驗飼料配方見表1。所有原料粉碎后經40目篩網過篩, 按配方比例充分混勻, 再加水攪拌后用制粒機(SLP-45, 上海漁業機械設備研究所,中國)制備成2 mm的飼料顆粒。為防止飼料中的脂肪酸氧化, 以上飼料在烘箱50℃烘干后于-20℃冰柜中保存備用。飼料脂肪酸組成見表2。

表1 飼料配方和化學組成(% 干物質)Tab. 1 Formulation and chemical composition of the experimental diets (% dry matter)

表2 飼料脂肪酸組成(% 總脂肪酸)Tab. 2 Fatty acid composition of the experimental diets (% of total fatty acids)
1.2 實驗魚及飼養
實驗所需的異育銀鯽“中科3號”(A strain)和“中科5號”(F strain)幼魚均由中國科學院水生生物研究所(湖北, 武漢)提供。在正式實驗前, 所有的魚都在室內循環水養殖系統中暫養2周以適應養殖環境,期間投喂2種飼料的等量混合物。將實驗魚饑餓24h后, 隨機挑取規格相近、外觀健康的不同品種異育銀鯽[“中科3號”: (5.01±0.05) g和“中科5號”:(5.02±0.05) g], 稱重后放入養殖缸中進行養殖實驗。每缸25尾, 每個處理3個平行。養殖實驗持續70d, 每天表觀飽食投喂3次(8:30、13:30和18:30)。實驗期間的水溫為(28.99±0.47)℃, 溶氧>7 mg/L, 氨氮<0.1 mg/L, pH為6.5—7.0。光照周期為12L∶12D(8:00—20:00光亮), 水面光照強度為2.79—3.32 μmol/(s·m2)。
1.3 樣品采集
在養殖實驗結束后, 將魚饑餓8h后取樣。用麻醉劑MS-222 (60 mg/L, Sigma, USA)將各處理缸中的實驗魚麻醉后計數稱重, 隨后進行取樣。每缸隨機挑選2尾魚, 用肝素鈉抗凝劑潤過的無菌注射器從尾靜脈取血并放入1.5 mL滅菌離心管中, 3000×g離心10min后得到血漿, 保存于-80℃用于后期分析。同時, 在冰浴條件下解剖實驗魚, 快速取得肝臟和肌肉樣品, 將其放入液氮中速凍并保存在-80℃。另外, 取肌肉組織樣品, 經冷凍干燥處理后用于測定脂肪酸組成。
1.4 樣品分析方法
參照AOAC方法[11]分析實驗飼料和魚樣的生化組成(水分、灰分和脂肪)。干物質在烘箱(電熱恒溫干燥箱, 精宏, 中國上海)于105℃干燥至恒重,通過失重法測定; 樣品在550℃下的馬弗爐(馬弗爐,中國湖北)中燃燒3h至恒重, 采用失重法測定灰分;粗脂肪分析以乙醚作為抽提液, 在索氏抽提儀(Soxtec System HT6, Tecator, Haganas, Sweden)中進行。參照Folch等[12]方法, 使用氯仿∶甲醇=2∶1混合試劑提取飼料和肌肉組織總脂, 用氣相色譜儀來測定脂肪酸的組成和含量(7890A-5975C, Agilents Technologies Inc., Santa Clara, CA, USA)。血漿葡萄糖、甘油三酯和膽固醇含量采用商品試劑盒(Fujifilm,Wako Pure Chemical Corporation, Osaka, Japan)進行測定。血漿低密度脂蛋白膽固醇(LDL-C)和高密度脂蛋白膽固醇(HDL-C)分別采用南京建成商品試劑盒A113-1-1和A112-1-1檢測(南京建成生物工程研究所, 中國南京)。
1.5 基因表達分析
使 用TRIzol試 劑(Invitrogen, Carlsbad, California, USA)從肝臟和肌肉組織中提取總RNA, 通過逆轉錄試劑盒M-MLV First-Strand Synthesis Kit (Invitrogen, USA)將其逆轉錄為cDNA。實時熒光定量在Light Cycle 480 Ⅱ(Roche, Basel, Switzerland)儀器 上 進 行, 采 用SYBR?Green Ⅰ Master (Roche,Basel, Switzerland)熒光染色劑測定肝臟和肌肉中脂肪代謝相關基因的相對表達量。選擇β-actin為內參基因, 基因相對表達量分析的計算方法參照Pfaffl[13], 引物信息見表3。

表3 本實驗所用引物序列表Tab. 3 Primers used in the present study
1.6 數據分析
所有數據使用統計軟件SPSS 18.0 (SPSS Inc.,Chicago, IL, USA)進行統計分析, 結果以均值±標準誤(mean±SE,n=6)表示。采用雙因素方差分析(Twoway ANOVA)來比較飼料(魚油vs.混合植物油)和品種(中科3號vs.中科5號)兩個因素的作用。當存在交互作用時(P<0.05), 用Student-Newman-Keuls多重比較分析各組間差異, 用小寫字母a、b或c來表示各組之間差異。當不存在交互作用時(P>0.05), 用大寫字母A和B表示品種間的差異, 大寫字母X和Y表示飼料間的差異。
2 結果
2.1 混合植物油替代魚油對不同品種異育銀鯽生長和飼料利用的影響
如表4所示, 異育銀鯽攝食混合植物油的攝食率均顯著高于魚油組, 但飼料效率顯著降低(P<0.05)。與“中科3號”相比, “中科5號”攝食率顯著降低, 但飼料效率顯著增加(P<0.05)。攝食魚油飼料組“中科5號”特定生長率顯著高于其他處理組(P<0.05)。
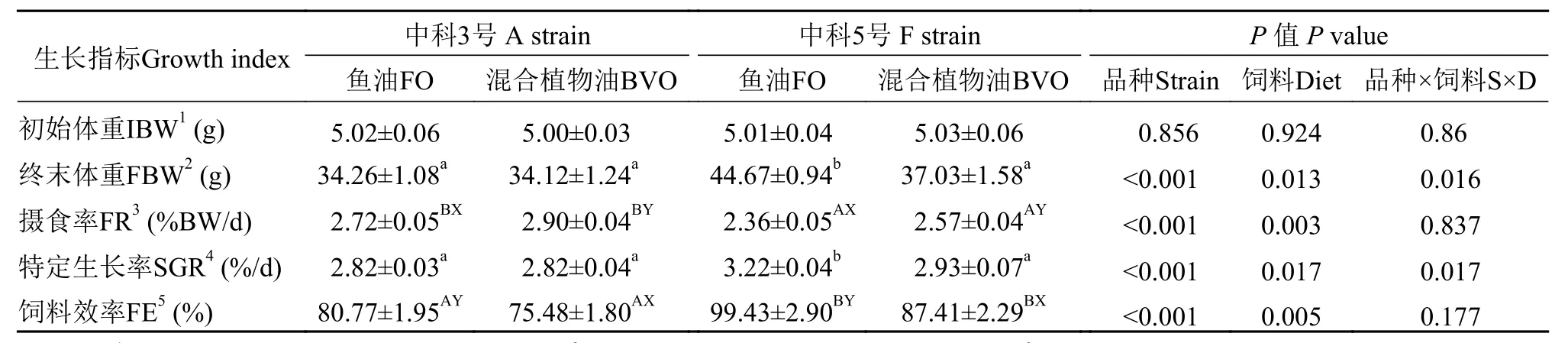
表4 飼料魚油和混合植物油對不同品種異育銀鯽生長和飼料利用的影響Tab. 4 Effects of dietary fish oil and blended vegetable oil on growth and feed utilization of two strains of gibel carp
2.2 混合植物油替代魚油對不同品種異育銀鯽魚體生化和肌肉脂肪酸組成的影響
如表5所示, 異育銀鯽攝食混合植物油飼料組魚體粗脂肪含量顯著高于魚油組(P<0.05)。異育銀鯽“中科3號”粗脂肪含量顯著高于“中科5號”, 而水分含量顯著低于“中科5號”(P<0.05)。灰分含量在各處理間無顯著性差異(P>0.05)。

表5 飼料魚油和混合植物油對不同品種異育銀鯽魚體基本組分的影響Tab. 5 Effects of dietary fish oil and blended vegetable oil on body composition of two strains of gibel carp
如表6所示, 與混合植物油組相比, 不同品種異育銀鯽攝食魚油飼料組Σn-6 PUFA相對含量顯著降低, 而Σn-3 PUFA相對含量顯著增加(P<0.05)。

表6 飼料魚油和混合植物油對不同品種異育銀鯽肌肉脂肪酸組成的影響Tab. 6 Effects of dietary fish oil and blended vegetable oil on fatty acids composition in the muscle of two strains of gibel carp
2.3 混合植物油替代魚油對不同品種異育銀鯽血漿生化指標的影響
如表7所示, 不同脂肪源飼料對“中科3號”血糖含量無顯著性影響, 而“中科5號”攝食混合植物油飼料組血糖含量顯著高于“中科3號”(P<0.05)。“中科3號”血漿低密度脂蛋白膽固醇含量顯著高于“中科5號”(P<0.05)。不論攝食何種飼料, 不同品種異育銀鯽血漿甘油三酯、膽固醇和高密度脂蛋白膽固醇含量無顯著性差異(P>0.05)。
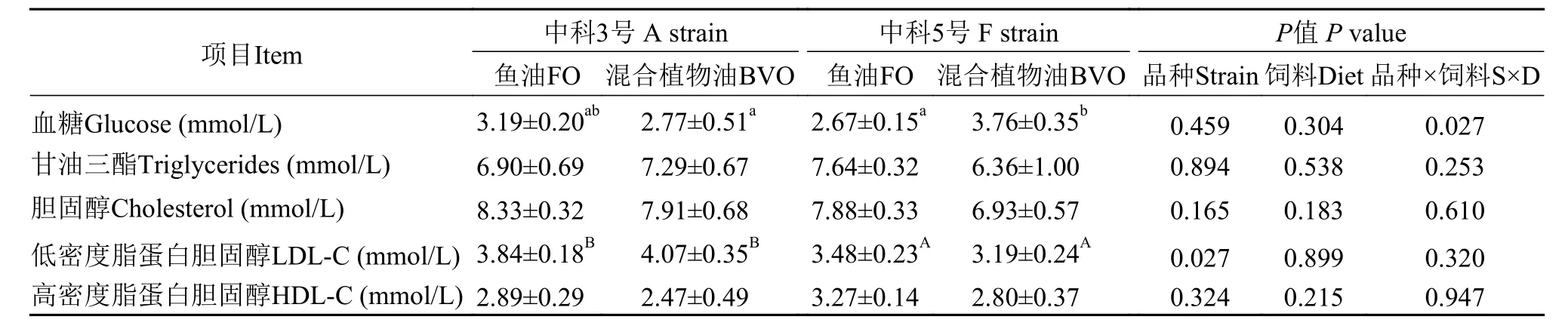
表7 飼料魚油和混合植物油對兩個品種異育銀鯽血漿生化指標的影響Tab. 7 Effects of dietary fish oil and blended vegetable oil on plasma biochemical indices of two strains of gibel carp
2.4 混合植物油替代魚油對不同品種異育銀鯽肝臟和肌肉脂肪代謝相關基因表達的影響
不同品種異育銀鯽肝臟和肌肉脂肪合成關鍵調控元件pparγ、srebp-1c及下游靶基因acly、acc和fas的基因表達量無顯著性差異(圖1A)。不同品種異育銀鯽攝食混合植物油飼料組肌肉srebp-1c和acc基因相對表達量顯著高于魚油組(P<0.05, 圖1B)。
不同品種異育銀鯽攝食魚油和混合植物油飼料后肝臟(圖2A)和肌肉(圖2B)脂肪分解代謝相關基因表達結果顯示, 魚油組肝臟脂肪酸β氧化關鍵基因pparα表達量顯著高于混合植物油組(P<0.05)。異育銀鯽“中科5號”肌肉脂肪分解相關基因hsl表達量顯著高于“中科3號”, 且脂肪酸β氧化相關基因aco3在肝臟和肌肉中的表達量均顯著高于“中科3號”(P<0.05)。脂肪分解代謝相關基因lpl(肝臟、肌肉)和cpt1a(肝臟、肌肉)、hsl(肝臟)和pparα(肌肉)的表達量均不受品種和飼料脂肪源的影響(P>0.05)。

圖2 飼料魚油(FO)和混合植物油(BVO)對“中科3號”(空白柱)和“中科5號”(斜線柱)肝臟(A)和肌肉(B)脂肪分解和脂肪酸β氧化相關基因相對表達量的影響Fig. 2 Effects of dietary fish oil (FO) and blended vegetable oil (BVO) on the mRNA levels of gene involved in lipolysis and fatty acids β oxidation in the liver (A) and muscle (B) of gibel carp A strain (blank bars) and F strain (bias bars)
不論攝食何種脂肪源飼料, 異育銀鯽“中科5號”肝臟脂肪酸轉運相關基因fabp1b和fatp1的表達量均顯著高于“中科3號”(圖3A)。飼料脂肪源和品種對異育銀鯽肌肉fabp1b和fatp1的表達均無顯著性影響(圖3B)。

圖3 飼料魚油和混合植物油對“中科3號”(空白柱)和“中科5號”(斜線柱)肝臟(A)和肌肉(B)脂肪酸轉運相關基因相對表達量的影響Fig. 3 Effects of dietary fish oil (FO) and blended vegetable oil (BVO) on the mRNA levels of gene involved in fatty acids transportation in the liver (A) and muscle (B) of gibel carp A strain (blank bars) and F strain (bias bars)
3 討論
3.1 飼料脂肪源作用
本實驗結果顯示, 異育銀鯽“中科3號”和“中科5號”攝食混合植物油飼料組攝食率顯著高于魚油組, 但飼料效率顯著低于魚油組, 表明異育銀鯽對魚油的利用優于混合植物油, 這與在鮭鱒魚類中的研究結果一致[14,15]。陳家林等[16]的研究結果表示,與全魚油組相比, 魚油分別與椰子油、玉米油、亞麻油1∶1的混合替代顯著提高異育銀鯽的生長性能,但對飼料效率無顯著影響。李新等[17]發現, 吉富羅非魚(Oreochromis niloticus)攝食棕櫚油飼料組攝食率顯著高于棉籽油組, 但飼料效率無顯著差異。飼料不同脂肪源對細鱗鮭(Brachymystax lenok)攝食率無顯著性影響, 其中魚油組的飼料效率最高[2]。紫蘇油、菜籽油、葵花油和棕櫚油完全替代魚油對紅羅非魚(Oreochromissp.)生長性能無顯著影響,棕櫚油組的飼料轉化率顯著高于魚油組[18]。因此, 魚類對植物油的利用差異與魚體本身(品種和規格)、脂肪源的種類和養殖條件等多種因素有關。不同品種異育銀鯽攝食混合植物油飼料攝食率顯著高于魚油組, 可能是通過增加攝食來滿足自身的能量需求。
在本研究中, 不同脂肪源對不同品種異育銀鯽血漿代謝物水平(血糖、甘油三酯、膽固醇、低密度脂蛋白膽固醇和高密度脂蛋白膽固醇)無顯著性影響。飼料脂肪酸組成影響魚類的脂肪合成、分解和轉運等脂肪代謝過程, 肝臟作為魚類代謝調節的中樞器官在脂肪代謝過程中發揮重要作用。在虹鱒(Oncorhynchus mykiss)中, 植物脂肪源完全替代魚油增強了肝臟脂肪合成相關基因的表達[19]。不同品系大西洋鮭(Salmo salar)攝食魚油和混合植物油飼料后, 肝臟長鏈多不飽和脂肪酸代謝存在顯著差異, 表現為Lean品系肝臟脂肪合成關鍵基因脂肪酸合成酶的表達顯著高于Fat品系[20]。混合植物油(菜籽油∶棕櫚油∶亞麻籽油=55%∶30%∶15%)完全替代魚油對虹鱒肝臟脂肪合成無顯著性影響[21]。由此可見, 魚的種類和飼料脂肪酸組成影響魚體的脂肪代謝過程。在本實驗中, 攝食混合植物油飼料組異育銀鯽肌肉脂肪合成關鍵基因srebp1和acc的表達量顯著上調, 且魚體脂肪含量顯著增高, 表明不同品種異育銀鯽攝食混合植物油飼料表現出更高的脂肪合成潛能。同樣, 不同品系大西洋鮭在攝食混合植物油飼料后, Lean品系大西洋鮭脂肪酸合成酶的基因表達水平較魚油組顯著上調[22]。脂肪合成的增加可能和飼料中n3/n6值有關, n-3 PUFA和n-6 PUFA比例的不平衡可能是造成肝臟脂肪含量增加的原因[23]。另外, 魚體的脂肪酸組成可以反映飼料脂肪酸組成[24,25]。在本實驗中, 不同品種異育銀鯽攝食魚油飼料組肌肉中n-3 PUFA的含量顯著高于攝食混合植物油飼料組, 而肌肉中n-6 PUFA的含量結果相反, 表現出與飼料中脂肪酸含量的一致性, 驗證了肌肉脂肪酸體現飼料脂肪酸組成這一結論。
3.2 品種效應
無論攝食何種脂肪源飼料, “中科5號”攝食率顯著低于“中科3號”, 而飼料效率顯著高于“中科3號”, 表現出較好的飼料利用性能。“中科5號”攝食不同脂肪源飼料體脂肪含量顯著低于而水分含量顯著高于“中科3號”, 與不同品種異育銀鯽攝食不同水平碳水化合物飼料和不同蛋白源飼料的結果一致[26,27]。機體脂肪積累是一個復雜的生理過程, 包含脂肪的轉運、攝取、合成和分解代謝過程等。在脂肪合成方面, 不同品種異育銀鯽肝臟和肌肉脂肪合成相關基因均無顯著差異。“中科5號”肌肉脂肪分解相關基因hsl和脂肪酸β氧化酶基因aco3的相對表達量顯著高于“中科3號”, 表明“中科5號”通過脂肪分解和脂肪酸β氧化過程導致了較低的體脂肪含量。該研究結果與不同品種異育銀鯽攝食不同飼料糖、脂水平的研究結果一致, 表明“中科5號”脂肪分解和脂肪酸氧化潛力優于“中科3號”[10]。另外, “中科5號”肝臟中脂肪酸轉運結合蛋白fabp1b和負責脂肪酸跨膜轉運的脂肪酸轉運蛋白1 (fatp1)的基因表達量均顯著高于“中科3號”,表明“中科5號”肝臟組織中脂肪酸的轉運攝取的潛力較強。綜上所述, 異育銀鯽“中科5號”對脂肪的轉運、分解、脂肪酸β氧化水平均顯著高于“中科3號”, 表現出對飼料脂肪更好的利用能力。
3.3 脂肪源和品種的交互作用
在本實驗中, 飼料脂肪源和品種對異育銀鯽特定生長率的影響存在交互作用。然而, 混合植物油完全替代魚油對“中科3號”的特定生長率無顯著性負面影響, 且“中科5號”攝食混合植物油可以達到與“中科3號”攝食魚油組一致的生長性能, 表明在“中科5號”飼料中使用植物性脂肪源存在巨大潛能。血液生化指標能夠反映魚體的生理狀態。在本研究中, 異育銀鯽血糖水平受到飼料脂肪源和品種的交互作用影響。與魚油組相比, 異育銀鯽“中科5號”混合植物油組的血糖含量顯著高于“中科3號”, 可能是由于“中科5號”傾向于利用脂肪酸氧化分解來為機體供能, 而非依賴于血糖的分解代謝供能。另外, 品種和飼料脂肪源的交互作用也體現在肌肉脂肪酸組成上, “中科5號”攝食魚油組n-3/n-6 PUFA顯著高于其他各組, 表明“中科5號”在利用魚油時體現出的更優質的肌肉脂肪酸組成。
4 結論
本實驗探究了飼料混合植物油完全替代魚油對不同品種異育銀鯽生長和脂肪代謝的影響。與魚油組相比, 混合植物油組顯著提高了不同品種異育銀鯽的攝食率, 但飼料效率有所降低, 表明植物油脂肪源對不同品種異育銀鯽的飼料利用存在一定的負面影響。在飼料脂肪源效應上, 混合植物油飼料通過增強肌肉組織脂肪合成相關基因srebp-1c和acc的表達來促進體脂肪的累積。在品種效應上, 不同品種異育銀鯽在體脂含量和肌肉脂肪代謝上表現出品種特異性。“中科5號”體脂肪含量顯著低于“中科3號”, 可能是其具有較高的肌肉脂肪分解、脂肪酸氧化及脂肪酸轉運能力導致的。飼料脂肪源和品種對異育銀鯽生長性能的影響存在交互作用, 但“中科5號”攝食混合植物油飼料可以達到與“中科3號”攝食魚油組相似的特定生長率, 且具有較高的肌肉n-3/n-6 PUFA水平, 表明混合植物油在“中科5號”飼料中具有廣泛應用的潛力。
