云南火焰蘭的花形態(tài)和花藥發(fā)育的胚胎學(xué)特征
2024-03-10 15:33:52王樂(lè)騁陶凱鋒朱永張錦羅艷李璐
廣西植物 2024年1期
王樂(lè)騁 陶凱鋒 朱永 張錦 羅艷 李璐
摘 要: ???關(guān)于珍稀瀕危植物火焰蘭屬的研究資料較少。該文以云南火焰蘭為研究對(duì)象,通過(guò)顯微鏡和石蠟切片技術(shù)觀察了云南火焰蘭的花形態(tài)和花藥發(fā)育胚胎學(xué)特征,并結(jié)合現(xiàn)有資料探討了其分類(lèi)學(xué)意義。結(jié)果表明:(1)具有分類(lèi)學(xué)意義的花形態(tài)特征為唇瓣三裂且花距不明顯,合蕊柱圓柱形,花藥帽紫色具黃條紋,花粉團(tuán)一對(duì)且由粘盤(pán)及粘盤(pán)柄連接。(2)花藥原基分化出一對(duì)側(cè)生花藥室,每個(gè)花藥室的小孢子囊中央分化出一條偏離中軸的不育隔膜組織,發(fā)育為一對(duì)不等大的次生小孢子囊;花藥成熟時(shí),因不育隔膜組織降解而形成2個(gè)深裂花粉團(tuán)。(3)發(fā)育完整的花藥壁有5~6層,為“多層型”花藥壁,包括表皮、2~3層藥室內(nèi)壁、中層和絨氈層;絨氈層細(xì)胞為單核、腺質(zhì)型。花藥成熟時(shí),藥室內(nèi)壁發(fā)生纖維狀加厚,花藥室在遠(yuǎn)軸端開(kāi)裂散粉。(4)小孢子母細(xì)胞經(jīng)同時(shí)型胞質(zhì)分裂,形成正四面體和左右對(duì)稱(chēng)排列的小孢子四分體,小孢子保持在四分體內(nèi),經(jīng)有絲分裂發(fā)育為2-細(xì)胞型的四合花粉,排列緊密,形成堅(jiān)固的花粉團(tuán);在花藥發(fā)育過(guò)程中未觀察到花粉敗育現(xiàn)象。該研究結(jié)果為火焰蘭屬的分類(lèi)學(xué)和保護(hù)生物學(xué)提供了新資料。
關(guān)鍵詞: ?火焰蘭屬, 形態(tài)學(xué), 花粉團(tuán)發(fā)育, 多層型花藥壁, 小孢子發(fā)生, 雄配子體發(fā)育
中圖分類(lèi)號(hào): ??Q944. 58
文獻(xiàn)標(biāo)識(shí)碼: ???A
文章編號(hào): ??1000-3142(2024)01-0077-12
Floral morphology and embryological features of
anther development in Renanthera imschootiana
WANG Yuecheng1, TAO Kaifeng1, ZHU Yong1, ZHANG Jin1, LUO Yan2, LI Lu1*
( 1. Department of Biodiversity Conservation, Southwest Forestry University, Kunming 650224, China; 2. Department of Horticulture
and Gardening, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla 666300, Yunnan, China )
Abstract: ??There are few data on the rare and endangered orchids of Renanthera Lour. Floral morphology and embryological features of anther development in R. imschootiana were investigated, and the taxonomic significance of R. imschootiana was analyzed by comparing with existing data. ?The results were as follows: (1) Flowers were characterized by such taxonomic features as tri-lobed labium with an unconspicous spur, cylindrical column, purple anther cap with yellow stripes, and a pair of deeply cleft pollinia connected by a well-developed stipe and a viscidium. (2) Anther primordium was differentiated into two lateral thecae containing two densely dyed microsporangia, and then, a sterile septum was formed longitudinally and unequally which divided each microsporangium into two jointly sub-microsporangium. The septum became absorbed during microsporogenesis and collapsed when anther ripened, resulting in a pair of deeply cleft pollinia. (3) The developed anther wall comprised 5-6 layers, including an epidermis, 2-3 endothecium, a middle layer, and a tapetum, which conformed to the massive type. Tapetal cell was uni-nucleated and glandular. (4) Microspore mother cells underwent a simultaneous cytokinesis during meiosis resulting in the tetrahedral or isobilateral microspore tetrads. Finally, each microspore retained in tetrads and produced 2-celled pollen via mitosis. Pollen tetrads were compactly arranged, leading to a hard cleft pollinia in each thecae. No pollen abortion was observed during anther development. The results provide new reference for taxonomy and conservation of Renanthera.
Key words: ?Renanthera, morphology, pollinium development, anther wall of massive tape, microsporogenesis, male gametophyte development
火焰蘭屬(Renanthera)隸屬于蘭科樹(shù)蘭亞科(Epidenroideae)萬(wàn)代蘭族(Vandeae),物種數(shù)目相對(duì)較少,僅20種,主要分布于東南亞和熱帶喜馬拉雅地區(qū)(吉占和等,1999)。中國(guó)有3種,地理分布較為狹窄(金效華等,2019)。其中,火焰蘭(R. coccinea)分布略廣,見(jiàn)于中國(guó)的云南南部、廣西和海南,以及緬甸、泰國(guó)、老撾和越南(陳心啟等,1999; 吳征鎰,2003; 金效華等,2019)。云南火焰蘭(R. imschootiana)為中國(guó)云南特有種,僅見(jiàn)于模式標(biāo)本產(chǎn)地越南和中國(guó)的云南南部元江(吉占和等,1999)。近年發(fā)表的中華火焰蘭(R. sinica)為中國(guó)特有種,僅分布于中國(guó)云南文山(劉仲健等,2003)。火焰蘭屬是一類(lèi)大型附生蘭,莖長(zhǎng)、具攀附性,葉片革質(zhì),圓錐花序或總狀花序,具多朵花,花大、花色鮮紅或橘黃或帶斑點(diǎn),形似火焰,具有較高的觀賞價(jià)值(陳心啟等,1999),現(xiàn)已出現(xiàn)一些人工雜交園藝品種(羅劍飄等,2012; 曹雯靜等,2014)。
火焰蘭屬地理分布狹窄、野生居群和標(biāo)本館館藏稀少,不僅被《瀕危野生動(dòng)植物種國(guó)際貿(mào)易公約—附錄I》收錄(http: //www. cites.org.cn),還被列入《中國(guó)生物多樣性紅色名錄—高等植物》的極危種名單(楊明森,2013)、《國(guó)家重點(diǎn)保護(hù)野生植物名錄》(國(guó)家林業(yè)和草原局農(nóng)業(yè)農(nóng)村部,2021)和《云南省極小種群保護(hù)名錄(2021版)》(孫衛(wèi)邦,2021)。目前,關(guān)于火焰蘭屬的地理分布、居群結(jié)構(gòu)、生物學(xué)和繁殖生態(tài)學(xué)等資料相對(duì)匱乏,亟待補(bǔ)充(李娟等,2018)。李娟等(2018)在2014 — 2018年間,經(jīng)野外調(diào)查發(fā)現(xiàn),云南火焰蘭在云南元江有2個(gè)居群點(diǎn),位于海拔1 100~1 200 m范圍的常綠闊葉和落葉混交林中,附生在栓皮櫟(Quercus variabilis)和毛枝青岡(Cyclobalanopsis helferiana)的樹(shù)干上,共有12株,其中僅有1株更新幼苗,為衰退型種群,認(rèn)為開(kāi)花而坐果率低是其瀕危的直接原因。因此,有必要開(kāi)展云南火焰蘭的生殖生物學(xué)和傳粉生態(tài)學(xué)研究。
花形態(tài)和花藥發(fā)育是蘭科傳粉生物學(xué)、繁育系統(tǒng)和胚胎發(fā)育的主要內(nèi)容,包括傳粉綜合征(pollination syndrome)(Pansarin & Maciel, 2017; 黃雙全和郭友好,2000; Zhang et al., 2022)、花藥成熟時(shí)散粉單元(pollen dispersal unit)的分化(Singer et al., 2008)和花藥發(fā)育過(guò)程中小孢子發(fā)生和雄配子體發(fā)育的胚胎學(xué)特征(Swamy, 1949; Rao, 1967; Sood, 1989; Kant & Hossain, 2010; 張錦等,2019;譚慶琴等,2020;李璐等,2020)。這些特征在蘭科的亞科、族、亞族和屬及其種間的物種分類(lèi)和系統(tǒng)演化方面發(fā)揮著重要作用(Dressler, 1986, 1993; Freudenstein et al., 2002; Pridgeon et al., 2005; Freudenstein & Chase, 2015; Mosquera-Mosquera et al., 2019; 李璐,2023)。國(guó)產(chǎn)火焰蘭屬的3種植物在植株形態(tài)和花型花色方面極為相似,野外調(diào)查時(shí)極易混淆(陳心啟等,1999; 吳征鎰,2003; 金效華等,2019)。劉仲健等(2003)在首次描述中華火焰蘭的形態(tài)特征時(shí),重點(diǎn)比較分析了國(guó)產(chǎn)3個(gè)種的花形態(tài)特征,認(rèn)為花序、花色與唇瓣裂片是火焰蘭屬物種鑒定的關(guān)鍵特征。Rusdi和Najwa(2020)觀察了產(chǎn)自馬來(lái)西亞的貝拉火焰蘭(Renanthera bella)在人工栽培條件下的生長(zhǎng)發(fā)育過(guò)程,為火焰蘭屬的人工繁育提供了新資料。目前,關(guān)于珍稀瀕危植物火焰蘭屬的研究資料較少,考慮到花形態(tài)學(xué)和胚胎學(xué)的重要性,本文以云南火焰蘭為研究對(duì)象,通過(guò)解剖鏡和石蠟切片技術(shù)觀察其花形態(tài)和花藥發(fā)育的胚胎學(xué)特征,并結(jié)合現(xiàn)有資料比較分析了其分類(lèi)學(xué)意義,為該屬的物種鑒定和保護(hù)生物學(xué)提供新資料。
1 材料與方法
云南火焰蘭引種栽培于西南林業(yè)大學(xué)科研教學(xué)實(shí)踐基地(云南豐春坊生物科技有限公司蘭花種質(zhì)資源圃),正常生長(zhǎng)開(kāi)花,花期為每年的4—5月。2017—2020年,連續(xù)4年采集不同發(fā)育時(shí)期的花蕾和開(kāi)放花,固定于50%酒精中,常溫保存。取新鮮開(kāi)放花于體式顯微鏡(Leica M165 FC)下解剖觀察,記錄唇瓣、合蕊柱、花藥帽、花粉團(tuán)、粘盤(pán)和粘盤(pán)柄等花結(jié)構(gòu)的形態(tài)、顏色和大小等,并進(jìn)行拍照處理。取固定后的花蕾材料,按形態(tài)大小分為5個(gè)發(fā)育階段,每個(gè)階段取5~10朵,經(jīng)酒精系列脫水、二甲苯透明、浸蠟包埋,常規(guī)石蠟切片(Leica RM 2235),切片厚度為6~8 μm;用鐵礬—蘇木精染色、中性樹(shù)脂膠封片(李正理,2006),烘干后在顯微鏡下觀察、拍照,并使用圖像編輯軟件Adobe Photoshop 2020排版。
2 結(jié)果與分析
2.1 花形態(tài)結(jié)構(gòu)
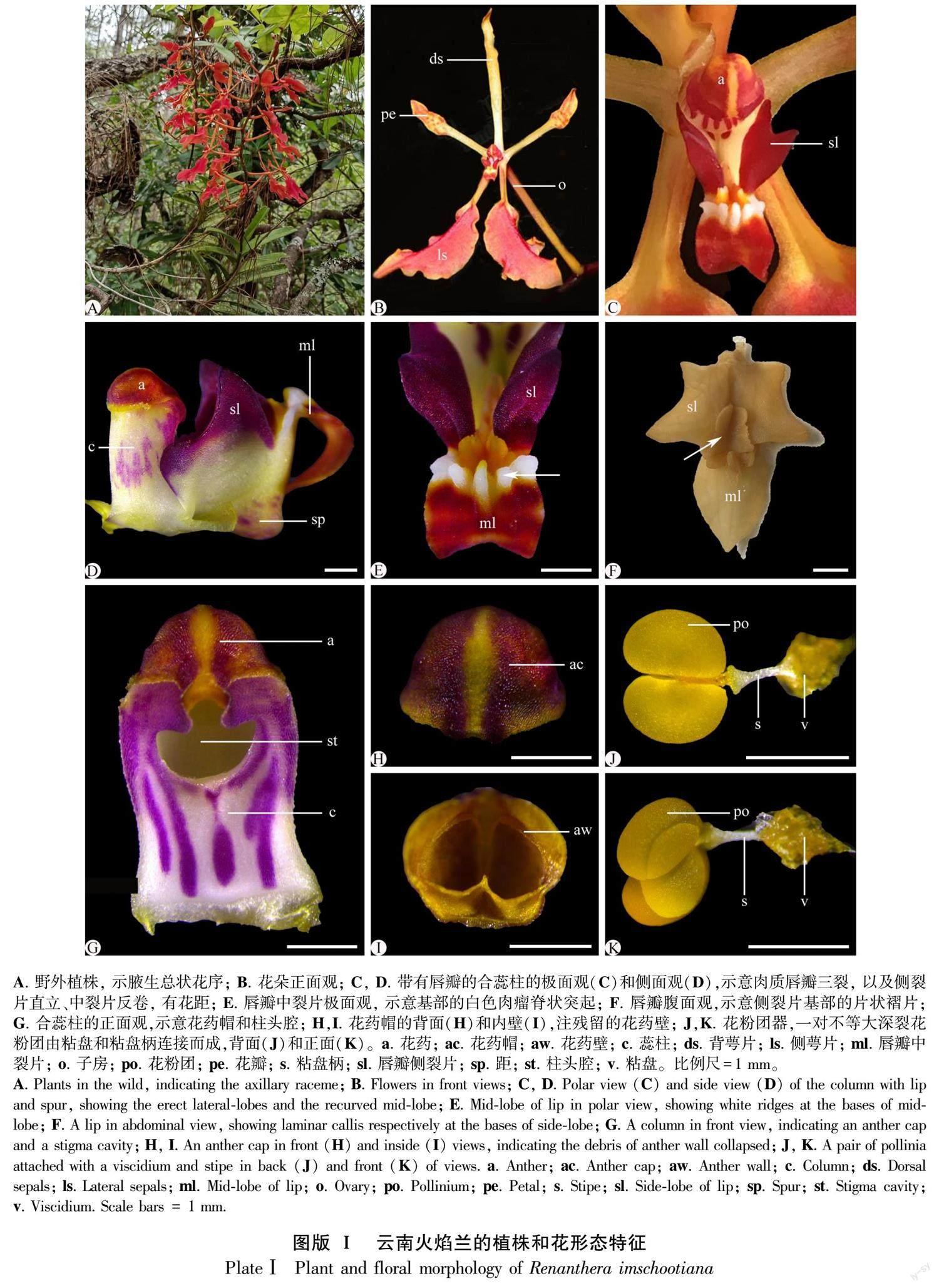
云南火焰蘭為大型附生草本,開(kāi)花時(shí)具有典型的腋生總狀花序,花序軸纖細(xì),多分枝,有5~30朵花(圖版Ⅰ:A)。花兩側(cè)對(duì)稱(chēng),中萼片和側(cè)花瓣倒披針形,均為淺黃色;兩枚側(cè)萼片狹匙形,橘紅色(圖版1:B)。肉質(zhì)唇瓣小而三裂,側(cè)裂片深紫,三角形、先端銳尖,直立、高于合蕊柱,基部具2枚邊緣不規(guī)則的黃色褶片(圖版Ⅰ:C-E);中裂片卵形,反卷,先端深紅色,基部具白色突起瘤狀物(圖版Ⅰ:E,F(xiàn)),花距不明顯,囊狀突起,長(zhǎng)約2 mm。合蕊柱短,長(zhǎng)4~5 mm,圓柱形、白底帶紫紅斑塊,柱頭腔深陷,下半部呈半圓形,上部開(kāi)口處有深凹裂縫,構(gòu)成向內(nèi)合攏的蕊柱翅(圖版Ⅰ:D,G)。花藥帽盔狀、半圓形,外部淺紫色,內(nèi)壁明顯分割為2室,在花藥開(kāi)裂花粉團(tuán)釋放后花藥壁殘留組織(圖版Ⅰ:H,I)。每個(gè)藥室有1枚花粉團(tuán),不等大深裂,遠(yuǎn)軸面較大,近軸面較小,由白色的細(xì)圓柱形粘盤(pán)柄和黃色的粘盤(pán)相連接,構(gòu)成帶有附屬結(jié)構(gòu)的花粉塊或花粉團(tuán)器(a pollinarium)(圖版Ⅰ[KG-0.8mm]: J,K)。
2.2 一對(duì)深裂花粉團(tuán)的發(fā)育模式
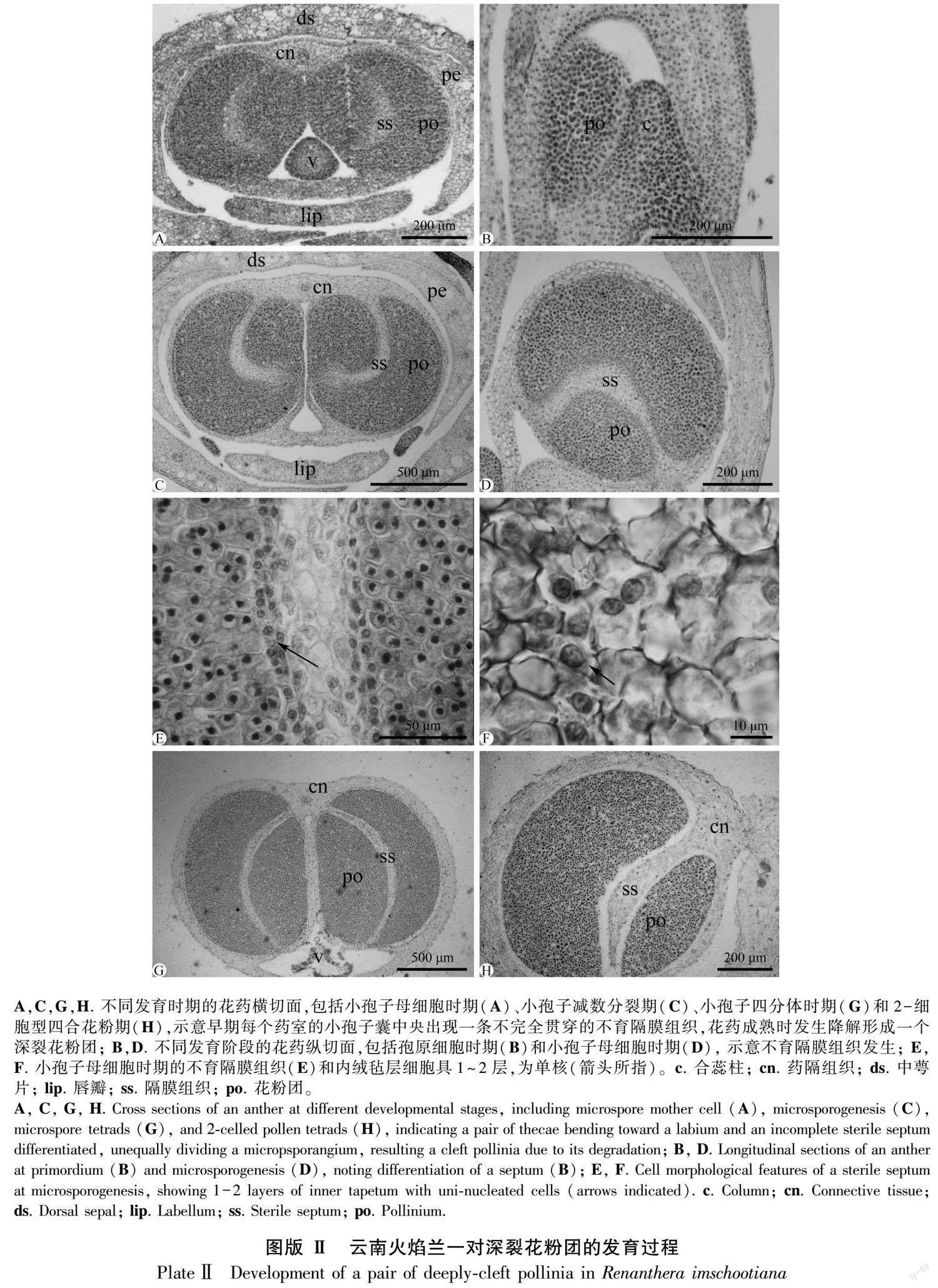
早期的花藥原基分化出面向唇瓣彎曲生長(zhǎng)的一對(duì)側(cè)生并列花藥室,每個(gè)花藥室分化出一個(gè)小孢子囊,由核質(zhì)比大、形態(tài)大小均勻的孢原細(xì)胞組成(圖版Ⅱ:A,B),繼而分化為一團(tuán)染色較深的小孢子囊和一條染色較淺的不育隔膜組織(圖版Ⅱ:A,C,D)。不育隔膜組織由染色較淺的薄壁細(xì)胞組成,明顯區(qū)別于細(xì)胞分裂活動(dòng)旺盛、染色較深的小孢子母細(xì)胞(圖版Ⅱ:E)。它從近藥隔組織端的小孢子囊中部出發(fā),先沿花藥室縱軸方向延伸生長(zhǎng)至小孢子囊中部,再橫折向藥室內(nèi)側(cè)生長(zhǎng),止于近花藥壁的5~6層小孢子母細(xì)胞處,分化為一條橫折彎曲、不完全貫穿小孢子囊的組織(圖版Ⅱ:C),分割出一對(duì)不等大、深裂的次生小孢子囊(圖版Ⅱ:C)。在小孢子母細(xì)胞減數(shù)分裂期間,不育隔膜組織靠近小孢子母細(xì)胞處的細(xì)胞分化為1~2層形態(tài)較小、近圓球形單細(xì)胞核的內(nèi)絨氈層(圖版Ⅱ:E,F(xiàn))。從小孢子四分體到雄配子體發(fā)育階段,每個(gè)花藥室的內(nèi)絨氈層和不育隔膜組織細(xì)胞開(kāi)始降解,形成空隙,把由小孢子囊發(fā)育而來(lái)的花粉囊分裂為一個(gè)不等大深裂的花粉團(tuán)(圖版Ⅱ:G, H)。因此,云南火焰蘭的成熟花藥里,產(chǎn)生了一對(duì)深裂的花粉團(tuán),這與花形態(tài)解剖觀察結(jié)果一致。
2.3 多層型花藥壁發(fā)育
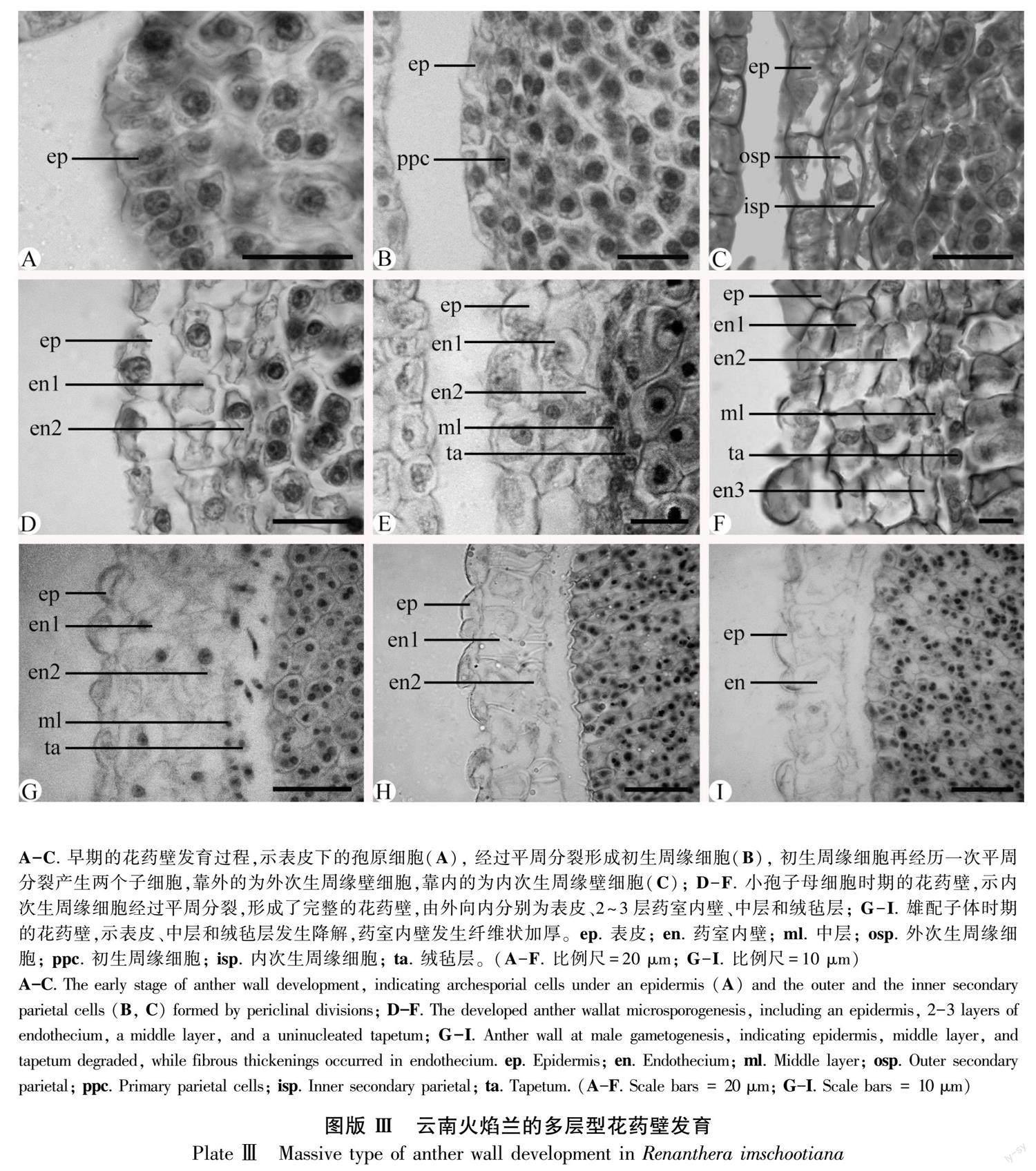
早期的花藥原基由表皮下的一團(tuán)染色較深的孢原細(xì)胞形成(圖版Ⅲ:A)。表皮下的孢原細(xì)胞分化為初生周緣壁細(xì)胞(圖版Ⅲ:B)。初生周緣壁細(xì)胞通過(guò)2~3次平周分裂,相繼發(fā)育為內(nèi)、外次生周緣壁細(xì)胞(圖版Ⅲ:C)。到小孢子母細(xì)胞時(shí)期,發(fā)育為完整的花藥壁,由外至內(nèi)分別為表皮、2~3層藥室內(nèi)壁、中層和絨氈層(圖版Ⅲ:D-F)。因此,花藥壁發(fā)育類(lèi)型為“多層型”(massive tape)(Sood, 1989)。此時(shí),表皮細(xì)胞和藥室內(nèi)壁細(xì)胞增大,呈方廓形;中層細(xì)長(zhǎng);絨氈層近圓球形、具單核、為腺質(zhì)型(圖版Ⅲ:E,F(xiàn))。在小孢子母細(xì)胞減數(shù)分裂期間,中層和絨氈層出現(xiàn)降解,表皮細(xì)胞發(fā)生退化,呈不規(guī)則鋸齒狀(圖版Ⅲ:G-I)。在雄配子體發(fā)育至2-細(xì)胞花粉時(shí)期,表皮逐步降解,有明顯殘跡,中層和絨氈層降解完全,出現(xiàn)明顯空隙,2層藥室內(nèi)壁細(xì)胞出現(xiàn)明顯環(huán)狀加厚(圖版Ⅲ:H,I),為藥室開(kāi)裂散粉提供了基礎(chǔ)。
2.4 小孢子發(fā)生及雄配子體發(fā)育
首先,早期的花藥原基表皮下包圍著一團(tuán)染色較深的孢原細(xì)胞,經(jīng)數(shù)次有絲分裂后分化為排列緊密、核質(zhì)比大的小孢子母細(xì)胞(圖版Ⅳ:A)。小孢子母細(xì)胞的核質(zhì)比增大,明顯呈多邊形,進(jìn)入減數(shù)分裂Ⅰ階段(圖版Ⅳ:B),產(chǎn)生了沒(méi)有子細(xì)胞壁的2個(gè)子核(圖版Ⅳ:C)。隨著發(fā)育的進(jìn)行,2個(gè)子核同時(shí)進(jìn)行下一步分裂,構(gòu)成了保持在同一個(gè)細(xì)胞壁內(nèi)、排列為正四面體或左右對(duì)稱(chēng)的4個(gè)子核(圖版Ⅳ:D)。然后,4個(gè)子核間出現(xiàn)縊痕,同時(shí)各分化出子細(xì)胞壁,構(gòu)成小孢子四分體,排列方式多為正四面體(圖版Ⅳ:E),偶見(jiàn)左右對(duì)稱(chēng)型(圖版Ⅳ:F),未見(jiàn)其他排列類(lèi)型。在雄配子體發(fā)育階段,小孢子四分體沒(méi)有從胼胝質(zhì)中釋放出來(lái),繼續(xù)通過(guò)有絲分裂,形成一個(gè)居中的大營(yíng)養(yǎng)細(xì)胞和一個(gè)靠邊的小生殖細(xì)胞(圖版Ⅳ:G)。最后,生殖細(xì)胞被吸入營(yíng)養(yǎng)細(xì)胞內(nèi),構(gòu)成了2-細(xì)胞型花粉(圖版Ⅳ:H)。4個(gè)成熟花粉保持在同一胼胝質(zhì)體內(nèi),形成了四合花粉,排列緊密且形成了堅(jiān)硬的成熟花粉團(tuán)。值得注意的是,本文選取約50朵花蕾作為研究材料,觀察了100個(gè)花藥室的發(fā)育,均未觀察到花粉敗育現(xiàn)象。
3 討論與結(jié)論
本研究通過(guò)解剖鏡和石蠟切片技術(shù),從花形態(tài)的結(jié)構(gòu)和花藥發(fā)育的細(xì)胞組織切片水平,揭示了云南火焰蘭的花形態(tài)特征和花藥發(fā)育的胚胎學(xué)特征,證實(shí)了一對(duì)深裂花粉團(tuán)的形成與每個(gè)花藥室小孢子囊中央的一條不完全貫穿的不育隔膜組織的分化和降解有關(guān),結(jié)合現(xiàn)有資料,分析了它們的分類(lèi)學(xué)及保護(hù)生物學(xué)意義。
3.1 可用于火焰蘭屬物種鑒定和分類(lèi)的花形態(tài)特征
首先,本研究支持前人觀點(diǎn),認(rèn)為總狀花序的著生位置、花色和唇瓣裂片特征是國(guó)產(chǎn)3種火焰蘭屬植物的物種鑒定依據(jù)(吉占和等,1999; 劉仲健等,2003; Chen et al., 2009; 金效華等,2019)。腋生總狀花序在云南火焰蘭和中華火焰蘭出現(xiàn),對(duì)生總狀花序見(jiàn)于火焰蘭。花色為橘紅色系見(jiàn)于云南火焰蘭和火焰蘭,花色淺黃色系帶紫紅色斑點(diǎn)在中華火焰蘭出現(xiàn)。唇瓣在3種中均為3裂,但中裂片和側(cè)裂片特征在種間差異明顯。中裂片上半部囊狀球形見(jiàn)于中華火焰蘭,其余2種均無(wú)囊狀結(jié)構(gòu)。側(cè)裂片為直立的三角形且基部具一對(duì)平行高褶片在云南火焰蘭出現(xiàn);方形或近圓形見(jiàn)于火焰蘭;卵圓形或披針形在中華火焰蘭出現(xiàn)(劉仲健等,2003)。本研究觀察到云南火焰蘭的唇瓣中裂片和側(cè)裂片基部的各有3枚白色肉質(zhì)隆起的脊?fàn)钗锖婉奁瑺铍蓦阵w,在其余2種中均未有詳細(xì)描述,值得進(jìn)一步研究。
其次,本研究認(rèn)為花粉團(tuán)數(shù)目及形態(tài)、粘盤(pán)與粘盤(pán)柄特征在火焰蘭屬和種間變化明顯,具有重要的分類(lèi)學(xué)意義。火焰蘭屬所在的樹(shù)蘭亞科具有高度愈合的花粉團(tuán)及花粉團(tuán)附屬物,包括花藥帽、粘盤(pán)、粘盤(pán)柄和花粉團(tuán)柄等(Dressler, 1993; Singer et al., 2008; Mosquera-Mosquera et al., 2019)。然而,關(guān)于這些花結(jié)構(gòu)特征的研究在火焰蘭屬資料中很少見(jiàn)(吉占和等,1999; 劉仲健等,2003; Chen et al., 2009; 金效華等,2019)。本研究利用解剖鏡觀察了云南火焰蘭的成熟花藥結(jié)構(gòu),發(fā)現(xiàn)花藥開(kāi)裂時(shí),露出2個(gè)不等大、深裂的花粉團(tuán),由近圓形且厚實(shí)的粘盤(pán)粘結(jié),并且該特征在組織切片上已得到驗(yàn)證。顯然,這與現(xiàn)有資料將火焰蘭屬描述為4個(gè)近等大的花粉團(tuán),基部有彈絲,粘盤(pán)柄稍長(zhǎng)而寬、粘盤(pán)厚、近圓形的特征明顯不同(吉占和等,1999)。同時(shí),中華火焰蘭作為新種發(fā)表時(shí),雖沒(méi)有詳細(xì)描述它的花粉團(tuán)數(shù)目和形態(tài),但線描圖記錄了其具一對(duì)不等大的花粉團(tuán)且粘盤(pán)為馬鞍形(劉仲健等,2003)。火焰蘭的花粉團(tuán)形態(tài)和數(shù)目不詳,粘盤(pán)柄被描述為中部屈膝狀(吉占和等,1999),但其他2種的粘盤(pán)柄均為平直(劉仲健等,2003)。因此,花粉團(tuán)形態(tài)和粘盤(pán)及粘盤(pán)柄等細(xì)節(jié)特征在火焰蘭屬具有重要的分類(lèi)學(xué)意義,需要對(duì)其花形態(tài)特征進(jìn)行準(zhǔn)確的描述。
3.2 花藥發(fā)育有助于確定指甲蘭亞族多樣的花粉團(tuán)類(lèi)型
長(zhǎng)期以來(lái),關(guān)于花粉團(tuán)形態(tài)和數(shù)目在樹(shù)蘭亞科,尤其是指甲蘭亞族不同類(lèi)群的變化及其分類(lèi)學(xué)意義頗有爭(zhēng)議(Dressler, 1993; Freudestein et al., 2002; Chase et al., 2015)。這主要是因一些物種的花粉團(tuán)特征(孔裂,或淺裂,或深裂)的位置較為隱蔽,往往藏在2個(gè)花藥室內(nèi)壁連接處,在解剖鏡下也不容易觀察清楚而造成的“誤判”。例如,巾唇蘭(Pennilabium yunnanensis)在解剖鏡下為一對(duì)全緣花粉團(tuán),但組織切片卻揭示其為一對(duì)細(xì)孔裂花粉團(tuán)(李璐等,2020)。Freudenstein和Rasmussen(1996)通過(guò)石蠟切片技術(shù),觀察了樹(shù)蘭亞科具不同花粉團(tuán)類(lèi)型的10屬代表類(lèi)群的花藥發(fā)育過(guò)程,揭示了2個(gè)、4個(gè)或8個(gè)花粉團(tuán)是早期花藥原基的小孢子囊內(nèi)分化出不同數(shù)目(2,4)且縱向或橫向排列的不育隔膜組織將其分割為次生小孢子囊,在小孢子發(fā)生和雄配子體階段被吸收而降解出現(xiàn)空隙,把成熟花粉囊分割為不同數(shù)目的花粉團(tuán)。類(lèi)似研究在其他樹(shù)蘭亞科成員里已得到了證實(shí)。例如,具有一對(duì)孔裂或淺裂的花粉團(tuán)是在花藥早期分化出一條不完全貫穿小孢子囊的不育隔膜組織(An incomplete sterile septum)在花藥成熟時(shí)因發(fā)生降解形成空隙而造成,見(jiàn)于美冠蘭(Eulophia hormusjii)(Bhanwra et al., 2003)、紋瓣蘭(Cymbidium aloifolium)(Kant et al., 2013)和大花萬(wàn)代蘭(Vanda coerulea)(張錦等,2019)。竹葉蘭(Arundina graminifolia)的8個(gè)花粉團(tuán)是花藥發(fā)育早期,每個(gè)藥室的小孢子囊分化出的一對(duì)十字交叉的不育隔膜組織,在花藥成熟時(shí)發(fā)生降解形成空隙,因?qū)⒒ǚ勰曳指魹槌?枚棒狀花粉團(tuán)而形成的(譚慶琴等,2020)。本研究從形態(tài)結(jié)構(gòu)和組織切片水平揭示了云南火焰蘭的花粉團(tuán)數(shù)目為2個(gè),呈一對(duì)深裂花粉團(tuán),而非如資料所記載的火焰蘭屬具4個(gè)花粉團(tuán)(吉占合等,1999)。因此,有必要通過(guò)組織切片觀察蘭科花藥發(fā)育過(guò)程中不育隔膜組織分化的數(shù)目及其排列位置,以及它們?cè)诔墒旎ㄋ庪A段因發(fā)生降解而導(dǎo)致的花粉團(tuán)形態(tài)和數(shù)目的關(guān)系(李璐等,2020)。這對(duì)于根據(jù)花粉團(tuán)數(shù)目和形態(tài)特征來(lái)探討萬(wàn)代蘭族—指甲蘭亞族的分類(lèi)學(xué)和系統(tǒng)學(xué)具有重要啟示(Freustein et al., 2002)。
3.3 蘭科“多層型”花藥壁發(fā)育模式與花粉團(tuán)特征密切相關(guān)
本研究觀察到云南火焰蘭發(fā)育完整的花藥壁有5~6層,屬于“多層型”發(fā)育類(lèi)型(massive type)(Sood, 1989),具有重要的分類(lèi)系統(tǒng)學(xué)和傳粉生態(tài)學(xué)意義。通常,被子植物的花藥壁層數(shù)均為5層以下,可歸為4種花藥壁發(fā)育類(lèi)型,包括基本型、單子葉型、雙子葉型以及簡(jiǎn)化型(Johri et al., 1992; 胡適宜,2005)。然而,蘭科的花藥壁層數(shù)在亞科和屬間變化較大,不僅有4層的單子葉型(monocotyledonous type),也有6~7層的多層型,值得收集更多資料分析其分類(lèi)學(xué)意義。在擬蘭亞科(Kocyan & Endress, 2001)、杓蘭亞科(Swamy, 1949; Ghimire et al., 2020)和蘭亞科(Sood, 1986; Sood, 1988; Kant & Goel, 2013)的一些物種里,花藥壁層數(shù)多為4層,屬于單子葉型。然而,在樹(shù)蘭亞科,花藥壁層數(shù)既有4層的單子葉型,也有多達(dá)6~7層的多層型。例如,單子葉型花藥壁見(jiàn)于原沼蘭屬 (Malaxis)(Sood, 1992)、石斛屬(Dendrobium)(Gurudeva, 2016)以及云南巾唇蘭(李璐等,2020)。多層型花藥壁首次命名于羊耳蒜屬(Liparis)(Sood, 1989),后來(lái)陸續(xù)見(jiàn)于蘭屬(Cymbidium)(Kant et al., 2013)、苞舌蘭(Spathoglottis plicata)(Sriyot et al., 2015)、美冠蘭屬(Bhanwra et al., 2003)等。其中,大花萬(wàn)代蘭(Vanda coerulea)的花藥壁超過(guò)7層,高達(dá)9層(張錦等,2019),這在現(xiàn)有資料里較為少見(jiàn)。關(guān)于多層型花藥壁,除了蘭科的樹(shù)蘭亞科外,在廣義夾竹桃科(Apocynaceae s. l.)也有報(bào)道(Johri et al., 1992; 艾靜等,2021),例如馬鈴果屬(Voacanga)的花藥壁普遍為8~9層,有的多達(dá)12~16層(Maheshwari, 1971)。因此,本研究認(rèn)為多層型花藥壁的出現(xiàn)與花粉團(tuán)的形成在蘭科和蘿藦科分類(lèi)系統(tǒng)與傳粉生態(tài)適應(yīng)性方面具有一定相關(guān)性(Endress, 2016),值得重視。
3.4 小孢子減數(shù)分裂的胞質(zhì)分裂方式和小孢子四分體排列多樣化的分類(lèi)系統(tǒng)意義
本研究觀察到云南火焰蘭的小孢子母細(xì)胞經(jīng)同時(shí)型胞質(zhì)分裂形成正四面體和左右對(duì)稱(chēng)的小孢子四分體,接著通過(guò)有絲分裂形成2-細(xì)胞型四合花粉,構(gòu)成堅(jiān)固致密的花粉團(tuán),這與蘭科現(xiàn)有研究結(jié)果一致(Johri et al., 1992)。在蘭科的花藥發(fā)育過(guò)程中,同時(shí)型胞質(zhì)分裂方式比較普遍,見(jiàn)于大部分蘭科類(lèi)群,如杓蘭亞科的杓蘭屬(Sood & Rao, 1986),樹(shù)蘭亞科的天麻(Gastrodia alata)(梁漢興,1983)、苞舌蘭(Sriyot et al., 2015)、巾唇蘭(李璐等,2020)和竹葉蘭(譚慶琴等,2020)等。相反,連續(xù)型胞質(zhì)分裂方式較少,出現(xiàn)在蘭亞科的眉蘭屬(Ophrys)(Aybeke, 2012)和綬草屬(Spiranthes)(Kant et al., 2013),樹(shù)蘭亞科的大花萬(wàn)代蘭(張錦等,2019)等。關(guān)于胞質(zhì)分裂方式的系統(tǒng)演化意義,已有研究認(rèn)為同時(shí)型胞質(zhì)分裂較為常見(jiàn),是被子植物的祖征,而連續(xù)型較為少見(jiàn),屬于衍征(Furness et al., 2002)。這在蘭科所在的單子葉植物綱或天門(mén)冬目(Asparagales)均有類(lèi)似的觀點(diǎn)(Rudall et al., 1997; Furness & Rudall, 1999)。因此,本研究支持在蘭科系統(tǒng)演化中,同時(shí)型胞質(zhì)分裂方式較為常見(jiàn),且屬于祖征。值得一提的是,同時(shí)型胞質(zhì)分裂方式也見(jiàn)于杓蘭亞科(Swamy,1949; Sood & Rao, 1986; Ghimire et al., 2020)。
本研究觀察到云南火焰蘭的小孢子四分體排列形式有2種,即正四面體和左右對(duì)稱(chēng),這與大部分蘭科的特征一致。現(xiàn)有資料報(bào)道了蘭科的小孢子四分體排列方式多樣,通常有2~3種,多達(dá)5種,包括正四面體、左右對(duì)稱(chēng)、十字交叉、“T”型和線型(胡適宜,1982; Johri et al., 1992)。小孢子四分體的排列方式基本與花藥成熟時(shí)四合花粉的排列方式保持一致,這在具有花粉小塊的蘭亞科(Kant & Arabaci, 2010; Kant et al., 2013)和具堅(jiān)固花粉團(tuán)的樹(shù)蘭亞科(張錦等,2019; 譚慶琴等,2020; 李璐等,2020)的花藥發(fā)育過(guò)程中得到證實(shí)。結(jié)合現(xiàn)有資料,本研究認(rèn)為,盡管每種蘭花的小孢子四分體排列形式多樣,但正四面體和左右對(duì)稱(chēng)最常見(jiàn),這也是蘭科小孢子發(fā)生的普遍特征之一。
3.5 云南火焰蘭花形態(tài)及花藥發(fā)育的保護(hù)生物學(xué)意義
本研究發(fā)現(xiàn)在花藥發(fā)育過(guò)程中,云南火焰蘭的小孢子發(fā)生和雄配子體發(fā)育均未出現(xiàn)花粉敗育現(xiàn)象。因此,前人提到過(guò)云南火焰蘭野外居群個(gè)體數(shù)量少(李娟等,2018)的原因與花粉發(fā)育無(wú)關(guān),但有必要從胚珠發(fā)育、大孢子發(fā)生、雌配子體形成以及雙受精后的胚胎和種子發(fā)育等方面開(kāi)展生殖生物學(xué)研究,為理解火焰蘭屬的繁殖生物學(xué)和瀕危機(jī)制提供新證據(jù)。
致謝 ?感謝云南豐春坊生物科技有限公司的王曉云女士和徐志鋒先生提供實(shí)驗(yàn)材料;玉溪市林業(yè)和草原局的吳建勇先生提供部分圖片; 西南林業(yè)大學(xué)云南生物多樣性研究院提供實(shí)驗(yàn)平臺(tái)。
參考文獻(xiàn):
AI ?J, LI L, WANG YP, et al., 2021. Sporogenesis and gametogenesis in Chinese endemic plant of Dregea sinensis Hemsl. var. sinensis (Apocynaceae sensu lato) and their taxonomic insights ?[J]. Guihaia, 41(12): ?2014-2023. ?[艾靜, 李璐, 王艷萍, 等, 2021. 中國(guó)特有種苦繩(廣義夾竹桃科)的大小孢子發(fā)生和雌雄配子體發(fā)育及其分類(lèi)學(xué)意義 [J]. 廣西植物, 41(12): 2014-2023.]
AYBEKE M, 2012. Anther wall and pollen development in Ophrys mammosa L. (Orchidaceae) ?[J]. Plant Syst Evol, 298(6): ?1015-1023.
BHANWRA RK, VIJ SP, 2003. The development of anther and male gametophyte in Eulophia hormusjii Duthie (Orchidaceae) ?[J]. Orchid Soc Ind, 17(1/2): ?87-91.
CAO WJ, LI HX, TANG YJ, et al., 2014. Karyotype analysis of four intragenus and intergenus hybridized combinations F1 generation of Renanthera ?[J]. Subtrop Plant Sci, 43(1): ?19-23. ?[曹雯靜, 李虹俠, 唐源江, 等, 2014. 4個(gè)火焰蘭屬內(nèi)及屬間雜交新組合F1代核型分析 [J]. 亞熱帶植物科學(xué), 43(1): 19-23.]
CHASE MW, CAMERON KM, FREUDENSTEIN, et al., 2015. An updated classification of Orchidaceae ?[J]. Bot J Linn Soc, 177(2): ?151-174.
CHEN XQ, JI ZH, LUO YB, 1999. Native orchids of China in colour ?[M]. Beijing: Science Press: 318. ?[陳心啟, 吉占和, 羅毅波, 1999. 中國(guó)野生蘭科植物彩色圖版鑒 [M]. 北京: 科學(xué)出版社: 318.]
CHEN XQ, LIU ZJ, ZHU GH, et al., 2009. Flora of China: Vol. 25 ?[M]. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press: 451-452.
DRESSLER RL, 1986. Features of pollinaria and orchid classification ?[J]. Lindleyana, 1(2): ?125-130.
DRESSLER RL, 1993. Phylogeny and classification of the orchid family ?[M]. Cambridge: Cambridge University Press.
ENDRESS PK, 2016. Development and evolution of extreme synorganization in angiosperm flowers and diversity: a comparison of Apocynaceae and Orchidaceae ?[J]. Ann Bot, 117(5): 749-767.
FREUDENSTEIN JV, RASMUSSEN FN, 1996. Pollinium development and number in the Orchidaceae ?[J]. Am J Bot, 83(7): 813-824.
FREUDENSTEIN JV, HARRIS EM, RASMUSSEN FN, 2002. The evolution of anther morphology in orchids: incumbent anthers, superposed pollinia, and the vandoid complex ?[J]. Am J Bot, 89(11): ?1747-1755.
FREUDENSTEIN JV, CHASE MW, 2015. Phylogenetic relationships in Epidendroideae (Orchidaceae), one of the great flowering plant radiations: progressive specialization and diversification ?[J]. Ann Bot, 115(4): 665-681.
FURNESS CA, RUDALL PJ, 1999. Microsporogenesis in monocotyledons ?[J]. Ann Bot, 84(4): ?475-499.
FURNESS CA, RUDALL PJ, SAMPSON FB, 2002. Evolution of microsporogenesis in angiosperms ?[J]. Int J Plant Sci, 163(2): 235-260.
GHIMIRE B, SON S, KIM JH, et al., 2020. Gametophyte and embryonic ontogeny: understanding the reproductive calendar of Cypripedium japonicum Thunb. (Cypripedoideae, Orchidaceae), a lady s slipper orchid endemic to East Asia ?[J]. BMC Plant Biol, 20(1): ?426.
GURUDEVA MR, 2016. Development of male and female gametophytes in Dendrobium ovatum (L.) Kraenzl. (Orchidaceae) ?[J]. J Orchid Soc Ind, 30: 75-87.
HU SY, 1982. Embryology of angiosperms ?[M]. Beijing: Higher Education Press. ?[胡適宜, 1982. 被子植物胚胎學(xué) [M]. 北京: 高等教育出版社.]
HU SY, 2005. Reproductive biology of angiosperms ?[M]. Beijing: Higher Education Press: 29-92. ?[胡適宜, 2005. 被子植物生殖生物學(xué) [M]. 北京: 高等教育出版社: 25-92.]
HUANG SQ, GUO YH, 2000. Research progress of pollination biology ?[J]. Chin Sci Bull, 45(3): ?225-237. ?[黃雙全, 郭友好, 2000. 傳粉生物學(xué)的研究進(jìn)展 [J]. 科學(xué)通報(bào), 45(3): 225-237.]
JI ZH, CHEN XQ, LUO YB, et al., 1999. Flora Republicae Popularis Sinicae: Vol. 19 ?[M]. Beijing: Science Press: 291-294. [吉占和, 陳心啟, 羅毅波, 等, 1999. 中國(guó)植物志: 第19卷 [M]. 北京: 科學(xué)出版社: 291-294.]
JIN XH, LI JW, YE DP, 2019. Atlas of native orchids in China ?[M]. Zhengzhou: Henan Science and Technology Press, 1036-1039. ?[金效華, 李劍武, 葉德平, 2019. 中國(guó)野生蘭科植物原色圖版鑒 [M]. 鄭州: 河南科學(xué)技術(shù)出版社, 1036-1039.]
JOHRI BM, AMBEGAOKAR KB, SRIWASTAVA PS, 1992. Comparative embryology of angiosperms ?[M]. Springer Berlin Heidelberg.
KANT R, ARABACI G, 2010. Development of anther in relation to sectile pollinium in ?Zeuxine strateumatica (Lindl.) Schltr ?[J]. J Biol Life Sci, 1(1): ?5-12.
KANT R, HOSSAIN MM, 2010. Development of pollinium in Malaxis muscifera (Lindl.) Kuntze ?[J]. Bangladesh J Botany, 39(2): ?193-198.
KANT R, GOEL S, 2013. Development of anther in Goodyera repens (L.) R. Br.: A massulate orchid ?[J]. Vegetos, 26(1): ?9-18.
KANT R, HOSSAIN MM, ATTRI LK, 2013. Pollinium development in Spiranthes sinensis (Pers.) Ames. and Cymbidium pendulum SW: A comparative study ?[J]. Bangladesh J Bot, 42(2): ?307-314.
KOCYAN A, ENDRESS PK, 2001. Floral structure and development of Apostasia and Neuwiedia (Apostasioideae) and their relationships to other Orchidaceae ?[J]. Int J Plant Sci, 162(4): ?847-867.
LI J, SHI M, L Y, et al., 2018. Discovery of an extremely endangered species Renanthera imschootiana and its population situation ?[J]. For Res, 31(3): ?9-14. ?[李娟, 石明, 呂亞媚, 等, 2018. 極危植物云南火焰蘭的發(fā)現(xiàn)及種群現(xiàn)狀 [J]. 林業(yè)科學(xué)研究, 31(3): 9-14.]
LI L, 2023. Research progress on diversity of androecium development of Orchidaceae [J]. Guihaia, 43(8): 1537-1552. [李璐,2023. 蘭科雄蕊發(fā)育多樣性研究進(jìn)展 [J]. 廣西植物, 43(8): 1537-1552.]
LI L, ZHANG J, WANG YP, et al., 2020. Pollinia development of in a new species of Pennilabium yunnanensis (Orchidaceae) with systematic implications ?[J]. Bull Bot Res, 40(2): 161-171. ?[李璐, 張錦, 王艷萍, 等, 2020. 蘭科新種巾唇蘭的花粉團(tuán)發(fā)育及分類(lèi)學(xué)意義 [J]. 植物研究, 40(2): 161-171.]
LI ZL, 2006. Plant tissue production science ?[M]. Beijing: Peking University Press. ?[李正理, 2006. 植物組織制片學(xué) [M]. 北京: 北京大學(xué)出版社.]
LIAN HX, 1981. Sporogenesis and the formation of male and female gametophyte of Gastrodiaelata [J]. J Integr Plant Biol, 3: 186-191. ?[梁漢興, 1981. 天麻大小孢子的發(fā)生和雌雄配子體的形成 ?[J]. 植物學(xué)報(bào), 3: 186-191.]
LIU ZJ, CHEN XQ, ZHANG JY, 2003. Renanthera sinica, A new species of Orchidaceae from Yunnan ?[J]. J Wuhan Bot Res, 21(1): ?37-39. ?[劉仲健, 陳心啟, 張建勇, 2003. 云南蘭科一新種——中華火焰蘭 [J]. 武漢植物學(xué)研究, 21(1): 37-39.]
LUO JP, WU KL, WENG SF, 2012. Research progress on Renanthera ?[J]. Guangdong Agric Sci, 39(10): ?69-72. ?[羅劍飄, 吳坤林, 翁殊斐, 2012. 火焰蘭屬植物研究進(jìn)展 [J]. 廣東農(nóng)業(yè)科學(xué), 39(10): 69-72.]
MAHESHWARI DH, 1971. Embryology of Apocynaceae I. Plumiereae ?[J]. J Ind Bot Soc, (50): ?74-85.
Ministry of Agriculture and Rural Affairs of State Forestry and grassland administration, 2021. List of National Key Protected Wild Plants in China ?[R]. Beijing: Ministry of Agriculture and Rural Affairs of State Forestry and Grassland Administration. ?[國(guó)家林業(yè)和草原局農(nóng)業(yè)農(nóng)村部, 2021. 國(guó)家重點(diǎn)保護(hù)野生植物名錄 [R]. 北京: 國(guó)家林業(yè)和草原局農(nóng)業(yè)農(nóng)村部.]
MOSQUERA-MOSQUERA HR, VALENCIABARRERA RM, ACEDO C, et al., 2019. Variation and evolutionary transformation of some characters of the pollinarium and pistil in Epidendroideae(Orchidaceae) ?[J]. Plant Syst Evol, 305(5): ?353-374.
PANSARIN ER, MACIEL AA, 2017. Evolution of pollination systems involving edible trichomes in orchids ?[J]. Aob Plants, 9(4): ?plx033.
PRIDGEON AM, CRIBB PJ, CHASE MW, et al., 2005. Genera Orchidacearum: Vol. 4. Epidendroideae(Part one) ?[M]. New York: Oxford University Press.
RAO AN, 1967. Flower and seed development in Arundina graminifolia ?[J]. Phytomorphology, (17): ?291-300.
RUSDI NA, NAJWA A, 2020. Scanning electron microscopy analysis of early floral development in Renanthera bella J. J. Wood, an endemic orchid from Sabah ?[J]. Pertanika J Trop Agric Sci, 43(3): ?377-389 .
SINGER RB, BARBARA G, CROSS H, et al., 2008. The use of orchid pollinia or pollinaria for taxonomic identification ?[J]. Selbyana, 29(1): ?6-19.
SOOD SK, 1986. Gametogenesis, integuments initiation and embryogeny in three species of Habenaria ( Orchidaceae, Orchideae) ?[J]. Proc Ind Acad Sci (Plant Sci), 96(6): ?487-494.
SOOD SK, RAO PRM, 1986. Gametophytes, embryogeny and pericarp of Microstylis wallichii Lindl. ?[J]. Bot Mag Tokyo, 99: 351-359.
SOOD SK, 1988. Development of gametophytes, embryogeny and pericarp in Goodyera repens (Orchidaceae, Neottieae) ?[J]. Proc Indian Acad Sci (Plant Sci), 98(2): ?149-156.
SOOD SK, 1989. Embryology and systematic position of Liparis(Orchidaceae) ?[J]. Plant Sys Evol, 166: 1-9.
SOOD SK, 1992. Embryology of Malaxis saprophyta, with comments on the systematic position of Malaxis (Orchidaceae) ?[J]. Plant Syst Evol, 179(1/2): ?95-105.
SRIYOT N, THAMMATHAWORN A, THEERAKULPISUT P, 2015. Embryology of Spathoglottis plicata Blume: A reinvestigation and additional data [J]. Trop Natural Hist, 15(2): ?97-115.
SUN WB, 2021. List of Yunnan protected plant species with extremely small populations (2021) ?[M]. Kunming: Yunnan Science and Technology Press. ?[孫衛(wèi)邦, 2021. 《云南省極小種群野生植物保護(hù)名錄(2021版)》 [M]. 昆明: 云南科技出版社.]
SWAMY B, 1949. Embryological studies in the Orchidaceae. II. Embryogeny ?[J]. Am Midl Nat, 41(1): ?184-201.
TAN QQ, LI L, ZHANG J, et al., 2020. Pollinia development in Arundina graminifolia (Orchidaceae) with taxonomic implications ?[J]. Guihaia, 40(1): ?83-94. ?[譚慶琴, 李璐, 張錦, 等, 2020. 傳統(tǒng)傣藥竹葉蘭的花粉團(tuán)發(fā)育及分類(lèi)學(xué)意義 [J]. 廣西植物, 40(1): 83-94.]
WU ZY, 2003. Flora of Yunnan: Vol. 14 ?[M]. Beijing: Science Press: 737. ?[吳征鎰, 2003. 云南植物志, 第14卷 [M]. 北京: 科學(xué)出版社: 737.]
YANG MS, 2013. China red list of higher plants—evaluation s report ?[M]. Beijing: Chinas Environmental Yearbook: 646. ?[楊明森, 2013. 中國(guó)生物多樣性紅色名錄——高等植物卷 [M]. 北京: 中國(guó)環(huán)境年鑒社: 646.]
ZHANG HP, TAO ZB, TRUNSCHKE J, et al., 2022. Reproductive isolation among three nocturnal moth-pollinated sympatric Habenaria species (Orchidaceae) ?[J]. Fr Plant Sci, (13): ?908852.
ZHANG J, ZHANG H, LI L, et al., 2019. Pollinia development in rare orchid of Vanda coerulea with its taxonomic implications ?[J]. Acta Bot Boreal-Occident Sin, 39(7): 1143-1153. ?[張錦, 張輝, 李璐, 等, 2019. 珍稀瀕危植物大花萬(wàn)代蘭的花粉團(tuán)發(fā)育及其分類(lèi)學(xué)意義 [J]. 西北植物學(xué)報(bào), 39(7): 1143-1153.]
( 責(zé)任編輯 蔣巧媛 )
收稿日期: ??2023-04-05
基金項(xiàng)目: ??國(guó)家自然科學(xué)基金(32060049); 中國(guó)科學(xué)院西雙版納熱帶植物園園林園藝中心研究基金(E2ZK291B05)。
第一作者: ?王樂(lè)騁(1998-),碩士研究生,研究方向?yàn)橘Y源利用與植物保護(hù),(E-mail)wangyuechengwyb@163.com。
* 通信作者: ??李璐,博士,副研究員,研究方向?yàn)橹参锵到y(tǒng)與進(jìn)化、植物形態(tài)解剖學(xué),(E-mail)lilu@swfu.edu.com。
