基于轉錄組學分析PI3K/AKT通路對冷卻灘羊肉貯藏期間肌肉代謝穩態的影響
2024-03-10 13:12:30羅瑞明胡曉磊畢永昭陳雪妍王金霞胡麗筠
食品科學 2024年3期
張 倩,楊 波,羅瑞明,胡曉磊,畢永昭,陳雪妍,王金霞,李 榮,胡麗筠
(寧夏大學食品與葡萄酒學院,寧夏 銀川 750021)
磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)是磷酸化細胞內磷脂酰肌醇的酶,根據它們的結構可分為3 類:I類PI3K、II類PI3K和III類PI3K[1]。其中,I類PI3K(PI3KI)調節內分泌反應,如細胞內胰島素和生長因子,并激活蛋白激酶B(protein kinase B,AKT),增強細胞葡萄糖攝取和刺激肌肉細胞合成糖原[2]。另外,AKT也能刺激蛋白質合成并抑制肌肉中蛋白質的降解[3]。由于PI3KI作為營養代謝的關鍵調控因子,并參與動物和人類的肌肉生理過程,因此它與肌肉穩態和生長之間的關系已有較多的研究。PI3KI在肌肉萎縮中也具有重要作用,特別是與人類疾病相關的高脂血癥[4]。最近的研究表明,其他類的PI3K也會影響葡萄糖和蛋白質代謝,并與人體的某些肌肉疾病有關[5-6]。PI3K/AKT信號通路利用多種調節因子調控細胞的生長、增殖以及代謝等生物過程維持細胞和生物體的穩態,如胰島素樣生長因子(insulin like growth factor,IGF)[7]。PI3K/AKT信號通過激活哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)調節葡萄糖穩態、脂質代謝、蛋白質合成等過程,這也受能量狀態和營養物的調節。伴隨其在刺激蛋白質合成和肌肉生長中的重要合成代謝作用,PI3K/AKT途徑也可能在抑制動物的蛋白質降解中發揮作用[8]。
灘羊是寧夏重點特色養殖的經濟動物,以其肉質鮮嫩、無膻味、含脂率低、營養含量豐富的獨特品質而深受大眾喜愛[9]。截止到2021年,寧夏“鹽池灘羊”品牌價值已高達88.17億 元[10]。肌肉纖維內蛋白質合成和降解之間的平衡影響肌肉質量和纖維大小,這兩個過程對營養狀況和激素平衡等因素都有影響。在脊椎動物中,當蛋白質降解速率大于蛋白質合成速率時,肌肉就會發生萎縮,導致質量下降[11-12]。反之,會發生肥大,導致肌肉質量增加。由于PI3K/AKT信號通路在刺激蛋白質合成和肌肉生長中的重要合成代謝作用,并調控肌肉生長發育、代謝調控及穩態維持等重要生物學過程,在抗肌萎縮蛋白缺陷型肌肉等疾病中均能檢測到PI3K/AKT信號通路相關基因的異常表達或蛋白的異常磷酸化[13]。近些年來,人們對PI3K/AKT信號通路的調控和功能有了深入的了解。然而,PI3K/AKT途徑仍然很復雜,特別在冷卻灘羊肉中,對PI3K/AKT信號通路的功能和代謝穩態動態調節的分子機制仍不清楚。因此,研究冷卻貯藏期間灘羊肉中PI3K/AKT信號通路的分子調控機制具有重要的意義。
隨著組學技術的興起,轉錄組學從RNA層次研究基因表達的情況,探索特定時期下生物體組織或器官的全部基因組轉錄情況及規律[14]。其中包含微陣列技術、基因表達序列分析、大規模平行測序和高通量測序等轉錄組測序技術[15]。而高通量測序以其檢測通量高、數字信號精確等優勢被廣泛應用[16-17]。近年來,中國鹽池灘羊基因組組裝已經完成,并通過大規模全基因組重測序研發出了鑒別技術,為灘羊肉品質改良和產地溯源提供了堅實的技術保障[18-19]。本研究旨在分析經PI3K抑制劑(LY294002)注射處理后冷卻灘羊肉貯藏期間轉錄組的信息,揭示PI3K/AKT信號通路在灘羊肉代謝調控中的作用,有助于研究PI3K/AKT信號通路在冷卻灘羊中的復雜功能,為冷卻灘羊肉貯藏期間肉品質量控制技術研發提供一定的理論參考。
1 材料與方法
1.1 材料與試劑
灘羊右后腿肉由寧夏鹽池縣大夏牧場食品有限公司提供。選取9 只體質量相近的6 月齡閹割公灘羊,屠宰前統一管理。依據GB 12694—2016《食品安全國家標準 畜禽屠宰加工衛生規范》要求屠宰,屠宰前無電刺激,屠宰后立即放入-20 ℃冰箱1 h,使其中心溫度降至0 ℃,密封托盤包裝后將其轉移至0~4 ℃冰箱貯藏。
HY-10108 LY294002溶液 美國MedChemExpress公司;TRIzol試劑盒 美國ThermoFisher Scientific公司;GeneReadTM 試劑盒德國Qiagen 公司;NEB fragmentation buffer 美國NEB公司;dNTPs 美國Thermo公司;DNA聚合酶I、RNase H 美國TaKaRa生物公司;AMPure XP beads 美國Beckman Coultier公司。
1.2 儀器與設備
NanoPhotometer納米光度計 德國Implen公司;2100生物分析儀 美國Agilent公司;Qubit 2.0熒光計上海Life Technologies公司;Illumina高通量測序儀中國Novogene公司;SW-CJ-1G超凈工作臺 上海博迅實業公司;ScanSpeed MiniVac Alpha冷凍離心機 美國Scan Speed公司;Thermo Scientific Forma 994 -80 ℃超低溫冰箱 美國Thermo公司;GeneAmp PCR system 9700普通聚合酶鏈式反應(polymerase chain reaction,PCR)儀美國Life公司;BA-200 Digital數碼三目攝像顯微鏡麥克奧迪實業集團有限公司。
1.3 方法
1.3.1 樣品采集
選用寧夏鹽池縣大夏牧區的灘羊作為實驗材料,采集在0~4 ℃條件貯藏的冷卻灘羊右后腿肉(從腰椎與薦椎連接處斬下的右后腿部位肌肉)。將取下的灘羊右后腿剔除可見脂肪及結締組織后迅速取約50 g 樣本作為0 d 樣品。其余樣品切成大小均一的塊狀,每塊約50 g,隨機分為對照組(命名為C)和LY294002組,共18 組。對照組不做任何處理,LY294002組以料液比10∶1注射10 μmol/L LY294002溶液。置0~4 ℃、相對濕度85%條件下,分別采集貯藏0、4、8 d后樣品各6 g封裝于滅菌冷凍管中,用液氮速凍后置于-80 ℃冰箱中用于轉錄組測序。對于不便立即測定的指標,在采樣點取下樣品后迅速置于液氮中冷凍并轉移至-0 ℃冰箱保存待用。每組樣本分別進行3 個生物學重復。
1.3.2 RNA提取、cDNA文庫構建和測序
LY294002注射組和對照組的樣品按照制造商的說明使用TRIzol試劑盒進行RNA提取[20]。使用NanoPhotometer納米光度計和2100生物分析儀檢測RNA樣品的數量和質量。樣品檢測合格后,通過GeneReadTM試劑盒去除rRNA。隨后加入NEB fragmentation buffer將mRNA打斷成短片段,以mRNA為模板,用六堿基隨機引物合成一鏈cDNA,然后加入緩沖液、dNTPs和DNA聚合酶I和RNase H合成二鏈cDNA,再用AMPure XP beads純化雙鏈cDNA。純化的雙鏈cDNA先進行末端修復、加A尾并連接測序接頭,再用AMPure XP beads進行片段大小選擇。最后進行PCR擴增,并用AMPure XP beads純化PCR產物,得到最終的文庫[21]。文庫構建完成后,先使用Qubit2.0熒光計進行初步定量,稀釋文庫至1.5 ng/μL,隨后使用2100生物分析儀對文庫的insert size進行檢測,insert size符合預期后,使用實時PCR(real-time PCR)方法對文庫的有效濃度進行準確定量(文庫有效濃度>2 nmol/L),以保證文庫質量。文庫制備物在Illumina HiSeq平臺上測序。
1.3.3 質量控制、轉錄組組裝和基因功能注釋
fastq格式的Raw reads首先通過內部perl腳本進行處理。在該步驟中,通過從Raw reads(原始序列)中去除接頭序列、包含poly-N的讀數和低質量讀數,獲得Clean reads(過濾序列)。同時,計算Q20、Q30、GC含量和過濾序列重復水平。所有下游分析都基于Clean reads。使用拼接軟件Trinity(version: r20140413p1)基于left.fq和right.fq完成轉錄組組裝,其中min_kmer_cov默認設置為2,所有其他參數默認設置。基因功能基于以下數據庫進行注釋:非冗余蛋白序列數據庫(Non-redundant Protein Sequence Database,Nr)、核酸序列數據庫(Nucleotide Sequence Database,Nt)、蛋白質序列數據庫(Swiss-Prot)、蛋白質家族數據庫(Protein Families,Pfam)、基因本體論(Gene Ontology,GO)和京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)。
1.3.4 差異表達基因(differential genes,DEGs)分析和功能富集
使用DESeq(2012)軟件包對兩個條件/組進行差異表達分析。使用Benjamini和Hochberg控制錯誤發現率的方法調整所得的P值。用基于負二項分布的DESeq進行差異分析,Padj<0.05和|log2FC|>1(FC表示差異倍數(fold chang))的基因被指定為差異表達[22]。通過微生信數據可視化網站(http://www.bioinformatics.com.cn/)實施DEGs的GO富集分析和KEGG途徑分析。0 d對比4 d組記為0/4 d組,4 d對比8 d組記為4/8 d組。
1.3.5 real-time PCR驗證
real-time PCR用于驗證RNA-Seq結果。隨機選取FC>2或<0.5且P<0.01的6 個DEGs進行real-time PCR,GAPDH基因用作內部參考基因。使用Primer Premier 5.0設計用于real-time PCR的引物,列于表1中。擴增程序根據以前的方法:95 ℃,30 s;95 ℃,10 s;60 ℃,30 s,40 個循環[23]。通過2-ΔΔCt計算目標mRNA的相對表達率。使用獨立樣本t檢驗比較對照組和LY294002處理組。當P<0.05時,差異被認為具有統計學意義。

表1 real-time PCR引物序列Table 1 Primer sequences used for quantitative real-time PCR
1.3.6 肌纖維直徑和間距測定
每組的肌肉組織用4%多聚甲醛固定液固定樣本,隨后進行脫水、修剪、包埋、切片、染色、鏡檢[24];采用數碼三目攝像顯微鏡觀察切片,測量肌纖維直徑和肌纖維間距,每個樣本共測量5 組數據,取平均值分別記為該樣本的肌纖維直徑和間距[25]。
1.4 數據處理
2 結果與分析
2.1 轉錄組測序和de novo組裝
對來自對照組的9 個cDNA文庫和來自LY294002組的9 個cDNA文庫進行測序[26]。共獲得115.50 Gb的序列,包括254941108 個Raw reads和248253912 個Clean reads。其中,96525684 個和151728228 個過濾序列分別來自對照組和LY294002組。對照組和LY294002組的堿基測序錯誤率分別為0.03%和0.02%。詳細信息如表2所示。18 個文庫過濾序列的從頭組裝產生62215 個單基因,總長度為183401678 bp,其中N50為1562 bp。從頭組裝詳細信息如表3所示。
2.2 基因功能注釋
對Nr、Nt、Swiss-Prot、Pfam、GO、KEGG數據庫進一步搜索,共組裝62215 個單基因。其中,獲得功能注釋信息的單基因分別為25459(26.48%)、42157(58.14%)、18908(19.15%)、24130(24.87%)、25067 個(31.15%)和11487 個(14.52%)。總共4863 個(6.42%)單基因可以與所有數據庫匹配,48129 個(58.41%)單基因至少與一個數據庫匹配(表4)。
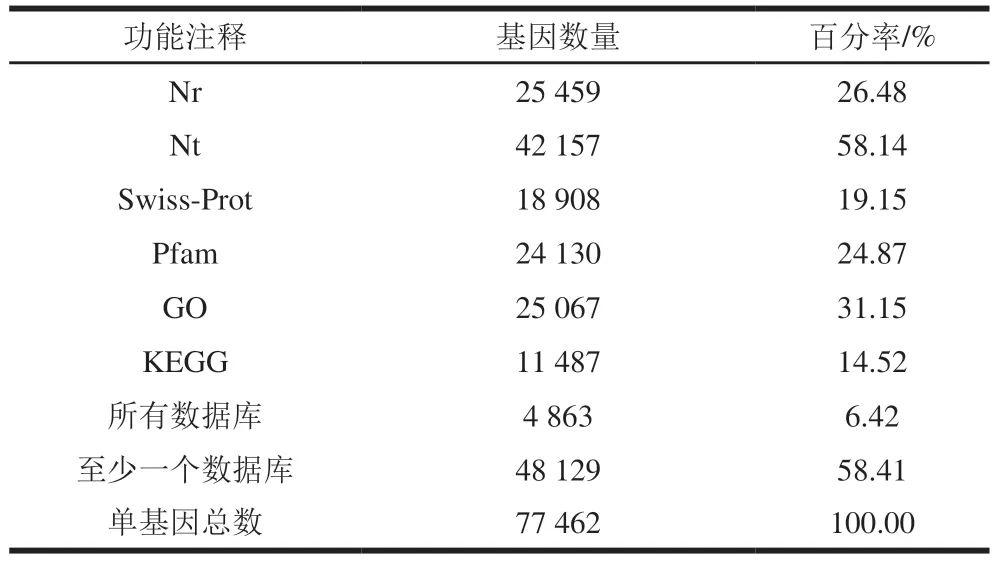
表4 灘羊肉轉錄組功能注釋結果Table 4 Results of function annotation of Tan mutton transcriptome
根據GO分析,25067 個單基因被分為3 個功能類別:生物過程、細胞成分和分子功能類別。0/4 d組這3 個功能類別中最主要的分別是單一生物代謝過程、細胞外隙和結構分子活動(圖1A),4/8 d組這3 個功能類別中最主要的分別是線粒體轉運、MHC II類蛋白復合體和氧化還原酶活性(圖1B)。表明0~4 d期間,位于細胞外隙的基因參與單一生物代謝過程等,這些基因編碼的蛋白質具備結構分子活性等功能。4~8 d期間,位于MHC II類蛋白復合體的基因參與調控線粒體轉運等過程,這些基因編碼的蛋白質具備氧化還原酶活性等功能。此外,KEGG分析顯示,10049 個單基因被映射到27 個通路上,這些通路可分為新陳代謝、遺傳信息處理、環境信息處理、細胞過程和生物體系統五大類。其中,0/4 d組富集DEGs較多的是信號轉導、能量代謝、碳水化合物代謝(圖2A);4/8 d組富集DEGs較多的是信號轉導、翻譯(圖2B)。在所有途徑中,信號轉導是富集到DEGs最多的途徑。通過進一步對通路篩選后,發現PI3K/AKT信號通路、氧化磷酸化通路、叉頭框蛋白O(forkhead box O,FOXO)信號通路以及mTOR信號通路在兩組DEGs中顯著富集。

圖1 單基因的GO分類Fig.1 GO classification of unigenes
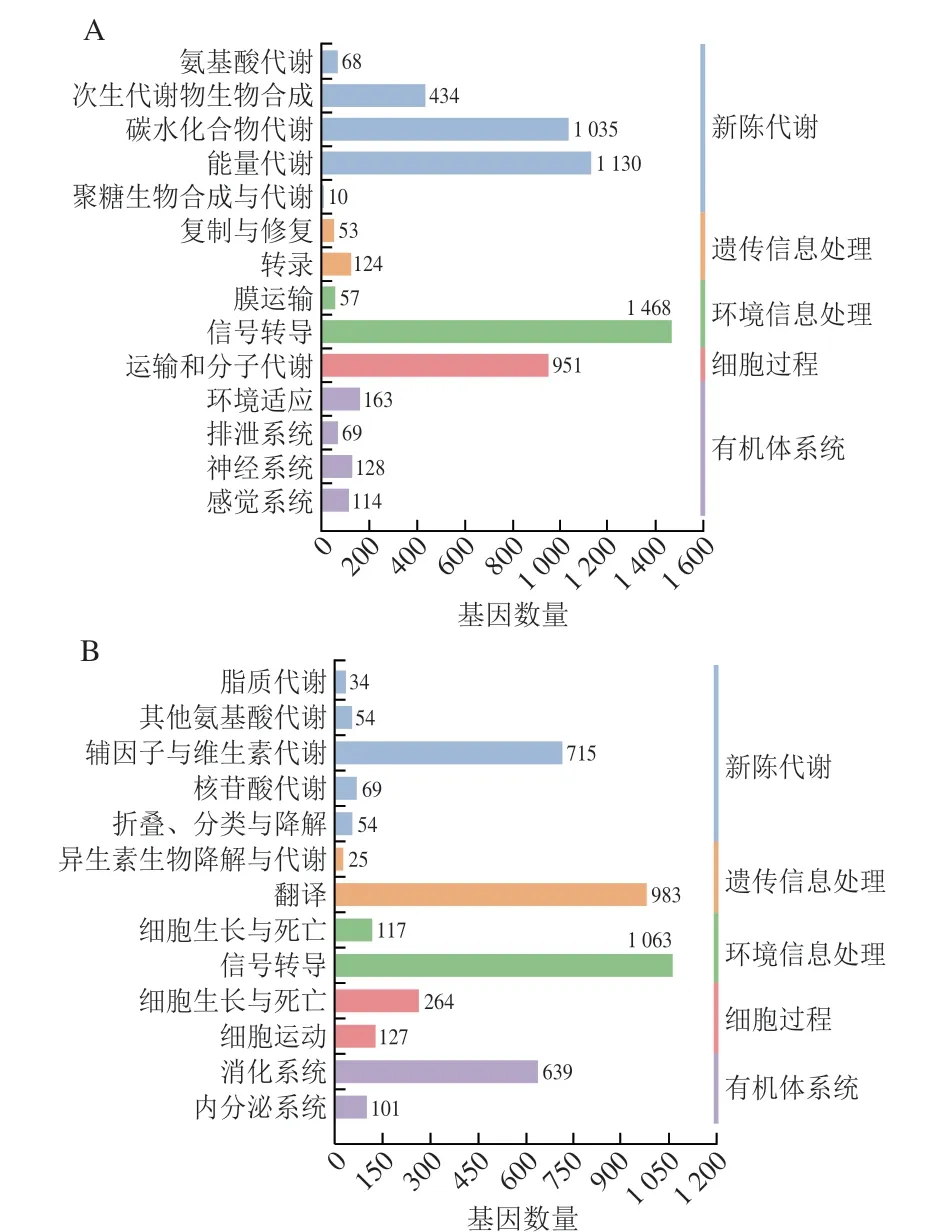
圖2 KEGG分類Fig.2 KEGG classification
2.3 差異表達分析和富集分析
為了比較0、4、8 d對照組和LY294002組之間的差異表達,使用DESeq(2012)軟件包識別DEGs,以Padj<0.05、log2FC>1或log2FC<-1為篩選條件,共發現583 個DEGs。其中,0 d對照組和LY294002組中發現474 個DEGs,包括320 個上調基因和154 個下調基因(圖3A),BAD基因極顯著上調可能是由于AKT活性受到抑制而促進B淋巴細胞瘤-2基因相關啟動子(Bcl-2asociated death promoter,BAD)與Bcl-2基因的結合,從而減弱Bcl-2基因抑制凋亡的作用。另外,Caspase-9蛋白水解酶被激活,促使Caspase-9降解肌肉結構蛋白并影響肌肉嫩度;4 d對照組和LY294002組中發現96 個DEGs,包括44 個上調基因和52 個下調基因(圖3B),AS160表達顯著下調會使下游葡萄糖轉運體(glucose transporter,GLUT)4蛋白降解,誘發骨骼肌攝取葡萄糖能力降低,進而導致葡萄糖代謝紊亂。CHI3L1基因表達下調可能是PI3K/AKT通路受到抑制后,激活了下游與碳水化合物分解代謝和蛋白質磷酸化酶等有關;8 d對照組和LY294002組中發現13 個DEGs,包括2 個上調基因和11 個下調基因(圖3C),上調基因與肌肉組織結構有關,包括肌球蛋白重鏈7B和肌鈣蛋白C1基因,下調的基因MYH7B、MYOZ2、RRAD與糖酵解受到抑制和鈣通道調節有關。

圖3 對照組和LY294002組DEGs分布趨勢火山圖Fig.3 Volcano maps of the distribution trends of DEGs between the control and LY294002 groups
為深入了解LY294002注射組可能差異調節的生物學過程,選用0/4 d LY294002組進行GO注釋分析。發現上調基因主要富集在30 個GO條目中,信號轉導、胞外區和受體結合分別是生物過程、細胞成分和分子功能類別中富集最多的亞類(圖4A)。下調基因也主要富集在30 個GO項,氨基聚糖代謝、胞外區和受體結合分別是生物過程、細胞成分和分子功能中富集最多的亞類(圖4B)。
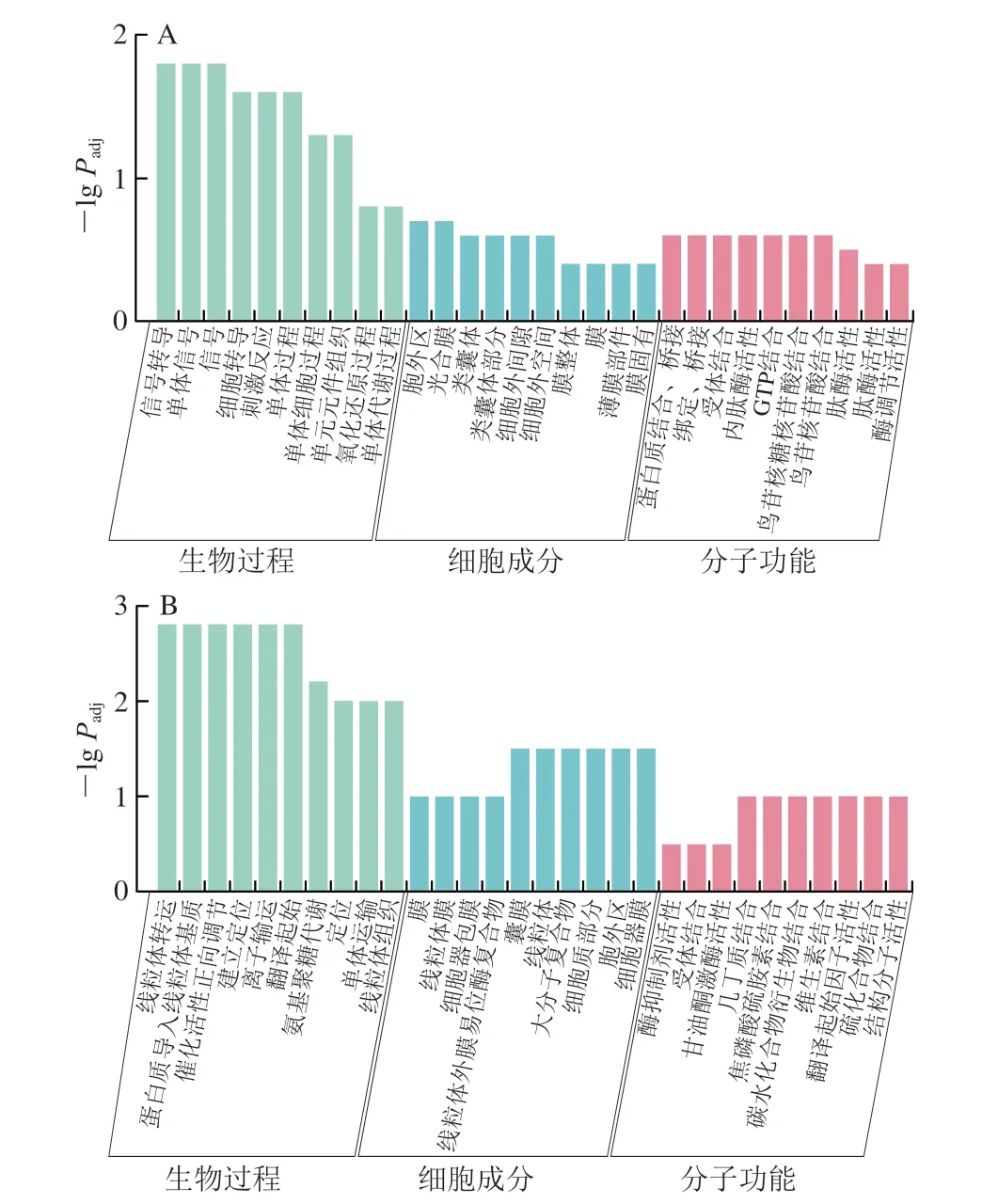
圖4 0/4 d LY294002組DEGs的GO注釋結果Fig.4 GO annotation results of DEGs in LY294002 group on days 0 versus 4
為進一步探索DEGs可能參與哪些信號通路,使用微生信在線軟件(http://www.bioinformatics.com.cn/)進行了KEGG通路富集分析,FDR≤0.05被視為富集通路,結果見圖5。

圖5 前20 DEGs的KEGG分類Fig.5 KEGG classification of top 20 DEGs
圖5A顯示了關于0/4 d LY294002組上調基因的最重要的前20 條途徑。在前20 條途徑中,精氨酸和脯氨酸代謝、ECM受體交互作用、PPAR信號通路、黏附斑激酶信號通路和甘油脂類代謝受到LY294002處理的影響。下調的基因在細胞衰老、糖酵解/糖異生、絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信號通路、氨基酸的生物合成和自噬途徑中顯著富集(圖5B)。KEGG分析結果表明,PI3K/AKT途徑在肌肉的氨基酸、葡萄糖和脂類代謝以及細胞命運中起著重要作用。
2.4 real-time PCR驗證
為證實RNA-Seq的結果,選擇涉及cAMP信號通路、轉運和分解代謝、精氨酸和脯氨酸代謝、細胞凋亡、甘油脂類代謝和MAPK信號通路的6 個代表性DEGs(表5),并應用real-time PCR測定這些基因的相對表達水平。結果顯示,這些基因的表達與來自RNA-Seq結果的FPKM值一致(圖6),這表明轉錄組測序數據可靠。
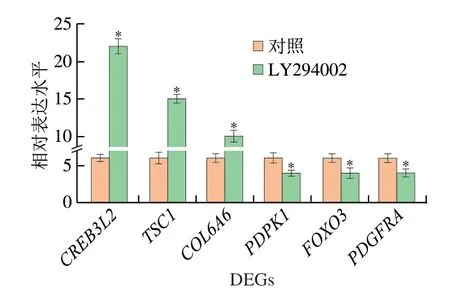
圖6 6 個DEGs的相對表達水平Fig.6 Relative expression levels of six DEGs

表5 6 個代表性DEGsTable 5 Six representative DEGs
2.5 組織切片分析
為了研究LY294002抑制劑對肌纖維的影響,石蠟切片觀察貯藏0 d時對照組和LY294002注射組的肌纖維。結果顯示,與對照組相比,LY294002注射組的肌纖維直徑顯著減小,肌纖維之間的間距顯著增加(表6)。此外,與對照組相比,LY294002注射組的細胞間隙變大,細胞質內容物明顯減少(圖7),和肌肉萎縮的特征相似。結合轉錄組測序的結果,表明LY294002抑制PI3K/AKT信號通路會導致肌肉細胞代謝失衡,進而影響肌肉細胞的生長,最終導致肌肉發生萎縮。

圖7 貯藏0 d對照組和LY294002組的灘羊肉組織切片Fig.7 Tissue sections of Tan mutton in the control and LY294002 groups

表6 0 d對照組和LY294002組肌纖維對比Table 6 Comparison of muscle fibers between the control and LY294002 groups on day 0
3 討論
PI3K/AKT信號通路作為細胞內的信號轉導通路,對細胞生長、存活和代謝控制等生物學功能產生影響[27]。為揭示PI3K/AKT信號通路對冷卻灘羊肉貯藏過程中的作用,采用高通量測序技術研究LY294002處理后冷卻貯藏期間灘羊肉轉錄組的變化。通過轉錄組分析,總共組裝了62215 個單基因,N50為1562 bp,其中62.57%可以被注釋。為進一步研究PI3K抑制劑(LY294002)對冷卻灘羊肉的影響,采用GO和KEGG富集分析對583 個基因進行了分類。GO分析表明DEGs主要富集于信號轉導過程和胞外區。KEGG富集分析表明,PI3K-AKT信號通路、氧化磷酸化通路、FOXO信號通路及mTOR信號通路為不同貯藏期間DEGs共同注釋到的代謝通路,這些途徑是肌肉細胞功能的關鍵部分。其中,胰島素受體通過參與PI3K-AKT途徑作用,發動細胞內信號轉導作用的多效級聯反應,刺激葡萄糖的吸收、增殖和蛋白質合成,并通過招募大量PI3K相關蛋白與肌動蛋白細胞骨架連接。因此,對灘羊肉品質特性如保水性及嫩度具有十分重要的調節作用。氧化磷酸化是肌肉運動時產生ATP的有效途徑,Jiang Shengwang等[28]研究表明,氧化磷酸化與糖酵解、脂肪酸代謝、細胞凋亡和肌肉收縮有關,對宰后肉品質形成具有重要的影響。FOXO信號通路在肌肉中的部分作用是調節轉錄,通過響應胰島素或IGF-1激活AKT誘導FoxO轉錄因子磷酸化,從而抑制其轉錄活性。此外,FoxO控制著肌肉中萎縮相關基因,包括Fbxo32和Trim63以及自噬基因,FoxO的缺失可以防止肌肉萎縮,貯藏期間肌肉能量代謝的差異可能與該通路變化有關。mTOR信號通路受到抑制時,肌肉代謝發生變化,包括氧化代謝受損,線粒體調節改變以及與AKT過度激活相關的糖原積累。Risson等[29]研究也表明,骨骼肌中的mTOR缺乏會導致代謝變化,從而導致糖原積累。因此,宰后肌肉代謝的轉換和肉品質變化均與貯藏期間肌肉轉錄水平的變化密切相關。
肌纖維是由蛋白質合成和蛋白質降解維持動態平衡的調節過程[30]。PI3K/AKT途徑不僅在蛋白質合成和肌肉生長中起重要作用,而且在抑制蛋白質降解中也發揮著作用[31]。Dong Jun等[32]研究發現PI3K信號通路能顯著促進肌肉的生長,通過其下游的β-連環蛋白調節成骨細胞并加快骨折愈合。PI3K/AKT也受分泌型糖蛋白(Wnt配體)調控,參與Wnt配體誘導的成骨細胞功能。該研究闡明了Wnt/PI3K/AKT/β-連環蛋白在成骨細胞中的功能聯系,突出了PI3K/AKT和Wnt/β-連環蛋白之間復雜的相互作用,它們在骨折愈合中起著至關重要的作用。王欣悅等[33]研究綿羊胚胎骨骼肌蛋白質組學數據發現,具有調控綿羊胚胎骨骼肌纖維生長發育作用的PI3K/AKT信號通路在骨骼肌纖維直徑增大的第105天顯著富集,AKT2是調控該信號通路的重要候選蛋白,并對骨骼肌纖維發育及分化產生影響。PI3K和mTOR的抑制劑對瘦素誘導的蛋白質分解顯著的抑制作用,這說明PI3K-AKTmTOR信號通路活化是瘦素調節蛋白質分解所必需的[8]。PI3K/AKT信號可以使FOXO轉錄因子磷酸化抑制蛋白質分解,FOXO轉錄因子通過調節參與肌肉萎縮基因的表達降低肌肉質量。同時發現當PI3K信號被LY294002抑制時,FOXO的表達顯著增加,這表明肌纖維中的蛋白質降解水平提高。組織學分析的結果顯示,肌纖維直徑減少和肌纖維之間的間距顯著增加,這與肌肉萎縮的特征相似[34]。Ang II能夠通過抑制PI3K/AKT/FOXO1信號通路的磷酸化,抑制蛋白的合成,誘導骨骼肌細胞萎縮[34]。Liu Lingyu等[35]研究發現福莫諾汀(formononetin,FMN)顯著抑制了大鼠肌肉生長抑制素α的表達。另外,FMN顯著增加了PI3K、AKT和FOXO3a的磷酸化以及CKD大鼠和C2C12肌管肌肉中肌源性增殖和分化標志物,促進了肌源性分化因子D和肌生成蛋白的表達。這些結果表明,PI3K/AKT/FOXO途徑調節蛋白質降解和合成之間的平衡,從而介導肌肉萎縮和維持骨骼肌質量。從刺激到細胞過程調節的信號轉導,包括參與葡萄糖穩態的過程,PI3K/AKT信號通路通過促進葡萄糖轉運、糖原合成介導肌肉代謝[12]。在本研究中,DEGs的KEGG途徑富集分析表明糖酵解/糖異生、碳水化合物代謝過程、甘油脂類代謝和脂類途徑的反應均受到影響。這些結果表明,PI3K/AKT通路的正向功能對于冷卻灘羊肉代謝控制是必不可少的,其功能障礙可能導致糖和脂質穩態受損。
PI3K/AKT信號傳導通過在生物體生長和關鍵細胞過程(如細胞周期進程、細胞生長和存活)期間介導生長因子信號,在細胞生理學中發揮重要作用[36]。MAPK也是關鍵信號通路,其激活對于肌肉細胞增殖是不可或缺的[37]。由于PI3K/AKT途徑的功能與Ras/MAPK途徑相似,所以發現這兩種途徑之間存在一定程度的相互作用[38]。在對這兩種轉導途徑相互作用的初步研究中,發現PI3K抑制劑可以降低MAPK的活性。Mano等[39]研究表明小鼠中的p38 MAPK信號通路被激活,導致泛素-蛋白酶體系統和自噬的激活。p38激活劑通過p38 MAPK信號通路的激活導致肌肉結構發生改變,PI3K活性對Ras/MAPK通路活性有非常重要的誘導作用。鑒于相互作用的復雜性,肌肉中PI3K/AKT途徑和Ras/MAPK途徑之間的真實關系尚未完全揭示。轉錄組數據顯示,一些上調基因富集在MAPK信號通路中,表明PI3K/AKT通路可能對MAPK信號通路有抑制作用。而湯崇輝等[40]發現作為PI3K/AKT抑制劑的渥曼青霉素對大鼠腦細胞中MAPK的活化有抑制作用。這些結果表明PI3K/AKT途徑和MAPK途徑之間的相互作用在不同的細胞類型和細胞分化的不同階段可能是不同的。
4 結論
本研究利用PI3K抑制劑LY294002對貯藏期間冷卻灘羊肉處理后進行高通量測序,轉錄組信息鑒定了583 個DEGs。0~4 d,DEGs富集較多的途徑是信號轉導、能量代謝及碳水化合物代謝;4~8 d富集DEGs較多的是信號轉導和翻譯過程。不同貯藏時間DEGs對肌肉代謝存在不同程度的調控作用,轉錄水平的變化主要通過氨基酸的生物合成、糖酵解/糖異生能力、甘油脂類代謝、MAPK信號通路的激活等過程參與調控肌肉代謝的轉換,為進一步闡明冷卻灘羊肉肌肉中PI3K/AKT代謝穩態的分子機制提供了方向。從應用的角度來看,建議控制PI3K活性在適當水平以維持冷卻灘羊肉的代謝穩態。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中國生殖健康(2019年3期)2019-02-01 06:12:26
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
中國衛生(2014年12期)2014-11-12 13:12:52
