熱處理對蕎麥麩皮蛋白結構與理化特性的影響
2024-03-10 11:25:14王立博任艷娟閆銘歡劉文超王昊然吳偉菁羅登林
食品科學 2024年4期
關鍵詞:結構
陳 靜,王立博,,任艷娟,閆銘歡,張 杰,張 彬,劉文超,王昊然,吳偉菁,羅登林,
(1.河南科技大學食品與生物工程學院,河南 洛陽 471023;2.河南省食品原料工程技術研究中心,河南 洛陽 471023;3.北京農學院食品科學與工程學院,北京 102206;4.廈門醫學院,福建 廈門 361023)
蕎麥,也稱凈腸草、烏麥、三角麥,屬于蓼科、蕎麥屬、雙子葉植物,常被歸類為“偽谷物”[1]。蕎麥因其生長周期短、適應惡劣環境能力強,成為許多地區糧食種植的重要作物之一[2]。目前已發現的蕎麥品種共有28 種,其中苦蕎與甜蕎已被大量種植與利用,是一種藥食同源食品[3]。蕎麥富含淀粉、蛋白質、黃酮等生物活性成分,攝入蕎麥能有效預防或輔助治療糖尿病、心血管硬化、高血壓、肥胖等慢性疾病[4]。近年來,隨著對蕎麥谷物資源的研究與開發,其作為功能性食材的可行性已被證實。市場中常見的蕎麥制品有面條、面包、餅干、饅頭、茶等[5-6],多是采用蒸、煮、炒等簡單的熱加工方式處理。而在熱加工過程中,蕎麥各功能組分結構、理化性質和功能特性等的變化規律,以及在加工中產生的影響亦是備受關注的熱點。
蕎麥麩皮富含優質蛋白質、多酚和膳食纖維,可有效預防或輔助治療多種慢性疾病[7]。而蕎麥麩皮作為蕎麥粉質加工中的主要副產物,常被用作動物飼料或肥料,未得到高值化利用。此外,蕎麥麩皮中蛋白質的質量分數高達21.6%~25.3%(干質量),是一種優質的植物蛋白來源[8]。蕎麥麩皮蛋白中還富含一般谷物蛋白缺乏的賴氨酸和精氨酸,其中,賴氨酸含量比大米和小麥多2 倍以上,可以有效預防賴氨酸缺乏癥,調節人體營養均衡[9]。但由于蕎麥的無麩質特性,導致蕎麥麩皮蛋白在食品加工及應用方面受到限制,致使蕎麥麩皮蛋白的營養功能并未得到充分的開發與利用。因此,本實驗以苦蕎和甜蕎麩皮為原料,通過等電點沉淀法制備出苦蕎麩皮蛋白(tatary buckwheat bran protein,TBBP)與甜蕎麩皮蛋白(common buckwheat bran protein,CBBP),并以市售大豆分離蛋白(soy protein isolate,SPI)為參考,經熱處理制備出3 種蛋白質的熱凝膠(TBBP-G、CBBP-G與SPI-G),通過觀察蕎麥麩皮蛋白與蛋白質熱凝膠分子質量、二級結構、內源熒光光譜、游離巰基與二硫鍵、微觀結構,熱特性及流變學特性的變化規律,分析熱處理對蕎麥麩皮蛋白結構及理化性質的影響,以期為推進蕎麥麩皮蛋白在食品領域中的加工與應用奠定基礎。
1 材料與方法
1.1 材料與試劑
苦蕎麩皮(黑豐1號)山西雁門清高食業有限責任公司;甜蕎麩皮(榆林紅花)定邊縣塞雪糧油工貿有限責任公司;大豆分離蛋白(分散型)河南鯤華生物技術有限公司。
考馬斯亮藍R250 上海藍季科技發展有限公司;十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)凝膠制備試劑盒 北京索萊寶科技有限公司;溴化鉀(光譜純)天津市科密歐化學試劑有限公司;三氯乙酸天津盛通泰化工有限公司;磷酸二氫鈉、磷酸氫二鈉、鹽酸胍、氫氧化鈉、鹽酸、石油醚、無水乙醇(均為分析純)天津市德恩化學試劑有限公司。
1.2 儀器與設備
UV-2100型紫外-可見分光光度計 尤尼柯(上海)儀器有限公司;Mini-PROTEAN Tetra電泳儀、ChemiDoc XRS+凝膠成像系統 美國伯樂公司;TDZ5-WS離心機 湘儀實驗室儀器開發有限公司;FE20 pH計、DSC1差示掃描量熱儀 梅特勒托利多科技(中國)有限公司;LGJ-10D冷凍干燥機 北京四環起航科技有限公司;DHR-2型旋轉流變儀 沃特世科技(上海)有限公司;VERTEX 70傅里葉變換紅外光譜(Fourier transform infrared spectroscopy,FTIR)儀 德國Bruker公司;TM3030 Plus臺式掃描電子顯微鏡 日立科學儀器(北京)有限公司;Cary eclipse熒光分光光度計美國安捷倫公司。
1.3 方法
1.3.1 蕎麥麩皮蛋白的制備
以蕎麥麩皮粉為原料,按料液比1∶10(g/mL)加入體積分數75%乙醇溶液,用保鮮膜封口,于250 r/min常溫攪拌40 min,去除類黃酮、色素等物質。經4500 r/min離心10 min,收集沉淀,上述步驟重復5 次。按料液比1∶10(g/mL)在沉淀中加入石油醚,保鮮膜封口并置于通風櫥浸泡脫脂12 h,經體積分數75%乙醇溶液洗滌3 次,放入50 ℃烘箱干燥4 h備用[10]。
將處理后的蕎麥麩皮粉與蒸餾水按料液比1∶10(g/mL)混合均勻,滴加1 mol/L NaOH溶液調pH值至10,40 ℃、200 r/min水浴攪拌60 min,經4500 r/min離心20 min,收集上清液,滴加1 mol/L HCl溶液調pH值至4.2,經4000 r/min離心15 min后收集沉淀,經蒸餾水沖洗2~3 次,蒸餾水復溶后,用1 mol/L NaOH溶液調pH值至7,真空冷凍干燥后研磨(過200 目篩),即得蕎麥麩皮蛋白樣品粉末[10]。經凱氏定氮法測得苦蕎及甜蕎麩皮蛋白的純度分別為(96.11±0.54)%(以干質量計,下同)和(95.83±0.28)%。
1.3.2 蛋白質熱凝膠的制備
稱取15 g蛋白質樣品溶于85 mL蒸餾水中,制備質量分數為15%的蛋白分散液,用保鮮膜封口,常溫250 r/min攪拌30 min使其混合均勻,沸水浴加熱30 min,冷水浴冷卻,真空冷凍干燥后,即得蛋白質熱凝膠樣品,經研磨(過200 目篩)后,即為蛋白質熱凝膠粉末[11]。
1.3.3 SDS-PAGE
將蛋白質及其熱凝膠樣品粉末按料液比1∶100(g/mL)加入蒸餾水混合均勻,與上樣緩沖液按體積比1∶3混合,煮沸5 min冷卻后5000 r/min離心3 min,取上清液9 μL進行SDS-PAGE(分離膠體積分數12%,濃縮膠體積分數5%)。電泳采用恒流模式,分離膠電流為35 mA,濃縮膠電流為25 mA。電泳結束后,用考馬斯亮藍R250染色劑染色30 min,沖洗干凈后,常溫脫色至條帶清晰[12]。
1.3.4 二級結構的測定
稱取蛋白質及其熱凝膠樣品粉末2 mg與200 mg KBr(光譜純)均勻混合。研磨均勻后壓制成片進行測定,掃描范圍為4000~400 cm-1,所有樣品連續掃描32 次。樣品光譜數據的平均值經Thermo Scientific OMNIC軟件分析獲得,二級結構使用PeakFit v4.12軟件分析得到。
1.3.5 內源熒光光譜的測定
將蛋白質及其熱凝膠樣品粉末用磷酸鹽緩沖液(pH 7)稀釋至10 mg/mL,充分混合后以8000 r/min離心5 min獲得上清液。樣品上清液在295 nm波長處激發,掃描發射波長范圍300~400 nm,激發和發射狹縫寬均為5 nm,每個樣品平行掃描3 次[13]。
1.3.6 游離巰基與二硫鍵含量的測定
稱取80 mg蛋白質及其熱凝膠樣品粉末于25 mL試管中,與1 mL Tris-甘氨酸緩沖液混合均勻,再加入4.7 g鹽酸胍,并用緩沖液稀釋至10 mL[14]。
游離巰基含量測定:取1 mL混合溶液于離心管中,加入4 mL脲-鹽酸胍溶液和0.05 mL Ellman’s試劑振蕩混合,并測定溶液在412 nm波長處的吸光度,按式(1)計算游離巰基含量:
式中:A412nm為樣品在412 nm波長處的吸光度;D為稀釋因子,游離巰基稀釋因子為5.02,總巰基稀釋因子為10;ρ為樣品的質量濃度/(mg/mL)。
二硫鍵含量測定:取1 mL混合溶液于50 mL離心管中,加入0.05 mLβ-巰基乙醇與4 mL脲-鹽酸胍溶液,25 ℃避光保存1 h。在離心管中加入10 mL質量分數12%三氯乙酸溶液,混合均勻后繼續25 ℃避光保存1 h,5000 r/min離心10 min后收集沉淀,將沉淀與5 mL質量分數12%三氯乙酸溶液混合均勻,重復離心3 次。取沉淀物溶于10 mL 8 mol/L脲中,并加入0.04 mL Ellman’s試劑振蕩混合,測定412 nm波長處溶液的吸光度,將吸光度代入式(1)計算總巰基含量,并根據式(2)計算二硫鍵含量:
1.3.7 掃描電子顯微鏡觀察
將經研磨過篩的蛋白質樣品粉末以及冷凍干燥的蛋白質熱凝膠固體顆粒樣品置于附有導電硅膠帶的樣品臺上,用洗耳球吹去多余粉末,進行噴金處理,20 kV條件下觀察樣品形態。
1.3.8 熱特性的測定
稱取5 mg蛋白質樣品粉末,置于鋁坩堝中,用壓樣機密封,并以空白鋁坩堝為對照。氮氣流速40 mL/min,樣品掃描溫度范圍0~200 ℃,升溫速率為10 ℃/min[15]。
1.3.9 流變學特性的測定
采用DHR-2型旋轉流變儀,平板直徑40 mm,間隙1 mm,應變量0.1%,固定頻率1 Hz。稱取蛋白質樣品粉末與蒸餾水混合,制備體積分數10%的蛋白質溶液。取適量蛋白質溶液于流變儀平臺上,并在夾具邊緣涂一層硅油,防止升溫過程中水分蒸發,進行溫度掃描,25 ℃保持5 min,以10 ℃/min升溫至90 ℃,在90 ℃保持10 min,以10 ℃/min降溫至25 ℃,并在25 ℃保持5 min,測定蛋白的儲能模量(G’)與損耗模量(G”)。在25 ℃進行頻率掃描,角頻率范圍為0.1~100 rad/s。表觀黏度測定的剪切速率范圍為1~100 s-1[16]。
1.4 數據處理與分析
采用SPSS 26.0軟件對數據進行顯著性分析,使用Origin 2021軟件對實驗結果進行繪圖,每組樣品進行3 次平行實驗。
2 結果與分析
2.1 SDS-PAGE結果分析
由圖1可知,熱處理前后蕎麥麩皮蛋白的分子質量主要分布在中低分子質量區域。未經處理的TBBP與CBBP分子質量分布相似,集中于10~55 kDa,而分子質量為25 kDa與33 kDa的弱條帶可歸于13S球蛋白的堿性和酸性多肽鏈[17]。經熱處理后,TBBP-G在25~40 kDa之間的條帶灰度變弱,55 kDa附近的條帶消失;CBBP-G在33~40 kDa間的部分條帶灰度減弱或消失,40~55 kDa附近的條帶灰度明顯變弱,而在分子質量為25 kDa附近的條帶灰度略有加強,表明熱處理使得蕎麥麩皮蛋白發生降解。這可能歸因于熱處理使蕎麥麩皮蛋白中的部分亞基被降解為小分子質量的多肽或寡肽,使條帶主要分布在低分子質量區域[18]。而SPI經熱處理后,低分子質量條帶減弱,分子質量主要聚集在40 kDa與90 kDa,且SPI-G的分子質量略有升高。通過與SPI及SPI-G的分子質量分布進行對比,可以發現蕎麥麩皮蛋白與SPI間分子質量的分布范圍均有不同,熱處理會使蕎麥麩皮蛋白與SPI原有的大分子發生部分降解。
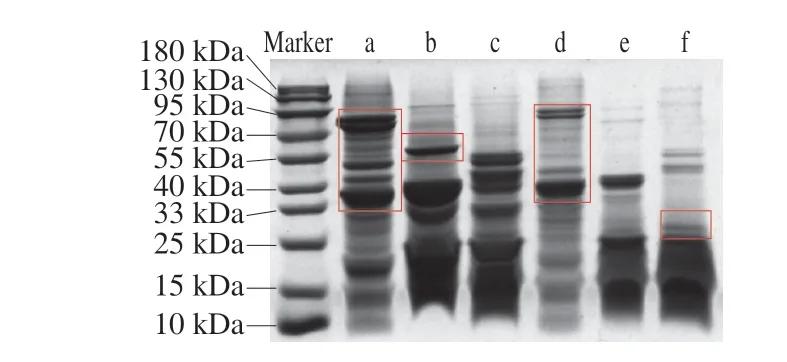
圖1 SPI、TBBP與CBBP及其熱凝膠的SDS-PAGE圖Fig.1 SDS-PAGE patterns of SPI,TBBP and CBBP and their thermally induced gels
2.2 二級結構結果分析
由圖2A可知,蕎麥麩皮蛋白在熱處理后并無新的吸收峰出現,只涉及峰強度與峰位的變化,說明熱處理沒有改變分子鏈的基團,而是引起分子內氫鍵的重組。SPI、TBBP與CBBP在3300 cm-1左右均有一個強吸收峰,表明蛋白質分子多聚體中可能存在大量分子內或分子間氫鍵[19]。熱處理使3 種蛋白在3300 cm-1左右的吸收峰發生藍移,分別從3296、3321 cm-1和3302 cm-1遷移至3286、3317 cm-1和3292 cm-1,這可能是由于加熱影響到蛋白質的空間結構,使蛋白質分子內氫鍵被破壞,其中TBBP-G的吸收峰偏移最小,說明熱處理對TBBP內氫鍵的影響小于SPI與CBBP。SPI、TBBP與CBBP在2931 cm-1處均有一個吸收峰,熱處理后藍移至2933 cm-1,這可能是因為熱處理誘導蛋白質分子內的C—H鍵伸縮振動增強,使蛋白質分子結構展開或斷裂,吸收峰發生遷移[20]。此外,熱處理后SPI-G、TBBP-G與CBBP-G在酰胺I帶(主要為C=O伸縮振動,1600~1700 cm-1)的吸收峰均發生偏移,并且TBBP-G與CBBP-G的偏移程度相對較大,這表明熱處理會使SPI、TBBP與CBBP分子中的α-螺旋結構向無規卷曲結構轉變[21]。
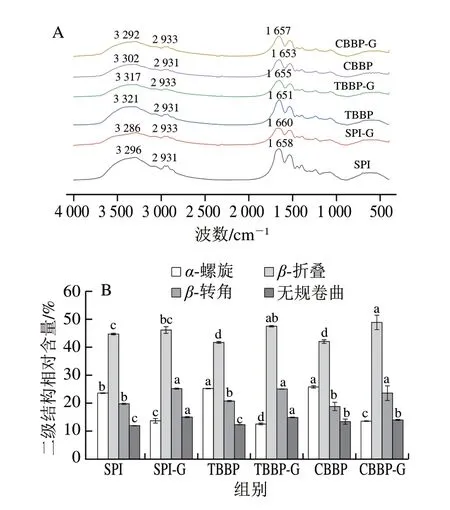
圖2 SPI、TBBP與CBBP及其熱凝膠的FTIR譜圖(A)與二級結構(B)Fig.2 FTIR spectra (A) and secondary structures (B) of SPI,TBBP and CBBP and their thermally induced gels
酰胺I帶包括α-螺旋、β-折疊、β-轉角和無規卷曲結構,常用于解析蛋白質的二級結構[22]。由圖2B可知,未經過熱處理的SPI、TBBP與CBBP中二級結構的主要組成為相對穩定的α-螺旋與β-折疊結構,兩者結構總占比可衡量蛋白質的穩定性[23],TBBP與CBBP中α-螺旋和β-折疊結構所占總比例相近(分別為66.95%和67.84%),均低于SPI(68.30%),表明制備的蕎麥麩皮蛋白二級結構的穩定性低于SPI。經熱處理后,SPI-G、TBBP-G與CBBP-G中α-螺旋結構的相對含量均降低,而β-折疊、β-轉角和無規卷曲結構占比均有不同程度的增加。這說明經過熱處理后,α-螺旋結構中氫鍵斷裂,使其發生解螺旋,R基團發生聚合現象,破壞了α-螺旋結構的穩定性,使SPI、TBBP與CBBP的二級結構由有序的α-螺旋結構轉變為相對無序的β-折疊、β-轉角和無規卷曲結構,從而影響蛋白質二級結構的穩定性[24]。同時,熱處理使SPI、TBBP與CBBP的α-螺旋結構發生解螺旋,形成不規則且疏松的片層狀凝膠聚集體結構(如圖4B、D、F),更有利于蛋白質與水分子相互作用,將水分子保留在凝膠結構的孔洞中,從而提高蛋白質熱凝膠的保水性[25]。
2.3 內源熒光光譜結果分析
熱處理可以不同程度地激發或抑制蛋白質中色氨酸、酪氨酸殘基暴露,產生熒光,因此通過內源熒光光譜圖可以觀察到熱處理對蛋白質三級結構的影響[26]。如圖3所示,經熱處理后,SPI、TBBP與CBBP的最大發射波長及熒光強度有不同程度的變化。其中TBBP與CBBP經熱處理后最大發射波長增大,即熱處理后發生紅移,并且TBBP-G與CBBP-G的熒光強度明顯降低,這是由于熱處理使得結構較為疏松的蕎麥麩皮蛋白發生變性,內部的色氨酸與酪氨酸殘基暴露在水溶液中,致使最大發射波長發生紅移,熱穩定性降低[27]。而SPI-G熒光強度略有增加,最大發射波長并未出現偏移,這說明熱處理下SPI的三級結構相對穩定。
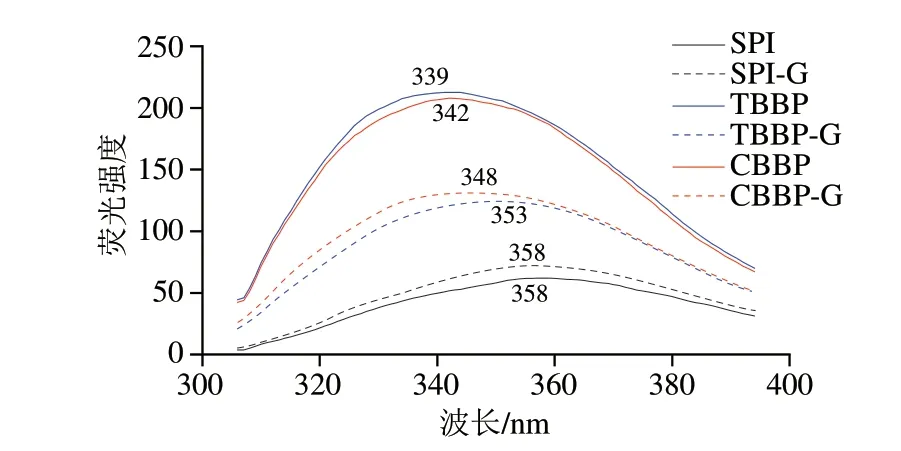
圖3 SPI、TBBP與CBBP及其熱凝膠的內源熒光光譜Fig.3 Intrinsic fluorescence spectra of SPI,TBBP,CBBP and their thermally induced gels
2.4 游離巰基與二硫鍵結果分析
如表1所示,TBBP與CBBP中游離巰基含量顯著高于SPI(P<0.05),這與3 種蛋白質的結構不同有關。蛋白質中游離巰基與總巰基的比值可反映解折疊的程度[28],TBBP與CBBP游離巰基與總巰基的比值分別為0.33與0.30,大于SPI(0.12),表明蕎麥麩皮蛋白解折疊程度更大,疏水性殘基相應暴露更多,使其表面疏水性更強。經加熱處理后,3 種蛋白質熱凝膠中游離巰基的含量均顯著增加(P<0.05),其中TBBP-G與CBBP-G分別增加至10.32 μmol/g和6.67 μmol/g,說明熱處理使部分包裹在蛋白質內部的巰基基團暴露出來[29]。此外,經過熱處理后,TBBP-G與CBBP-G中二硫鍵含量也隨之增加,特別是CBBP-G從5.70 μmol/g顯著增加至11.43 μmol/g(P<0.05),這歸因于高溫條件下蛋白質解折疊速率增加,暴露出更多的游離巰基,分子間相互作用增強,使游離巰基不斷向二硫鍵轉化[30]。而熱處理后形成的SPI-G中二硫鍵含量則顯著降低(P<0.05),可能是SPI熱穩定性較高,其分子結構未發生顯著改變,只有部分肽鏈展開,二硫鍵斷裂,導致SPI-G中游離巰基含量升高,二硫鍵含量降低[31]。
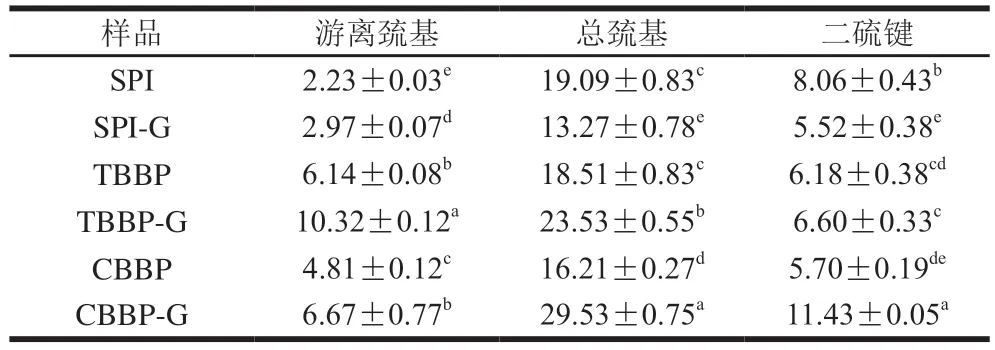
表1 SPI、TBBP與CBBP及其熱凝膠中巰基與二硫鍵含量Table 1 Contents of sulfhydryl and disulfide bonds in SPI,TBBP,CBBP and their thermally induced gels μmol/g
2.5 微觀結構分析
由圖4可知,SPI呈現大小不規則的球狀結構,并且部分較小的SPI球狀結構發生塌陷,這可能是在加工過程中受到機械損傷,使蛋白質粉末顆粒被破壞。而TBBP與CBBP結構相似,蛋白質粉末顆粒大小不均一,呈塊狀不規則聚集體,粉末顆粒內部結合緊密,未發現明顯孔洞,且表面粗糙有突起物。經熱處理后,SPI-G的球狀結構發生破裂,形成片層狀的網絡結構,且凝膠壁表面光滑,無明顯的小團聚體。TBBP-G形成表面更加粗糙的凝膠聚集體,不規則片狀聚集體略有疏松,可觀察到細小孔洞及裂縫[32]。而CBBP-G的凝膠結構疏松,可明顯觀察到部分凝膠壁薄片與脊狀結構,凝膠壁出現明顯裂紋且表面粗糙,有少量碎屑。上述結果表明,高溫熱處理破壞了SPI、TBBP與CBBP分子顆粒的分布與狀態,并且與SPI-G呈現的表面光滑且致密的片層狀網絡結構不同,蕎麥麩皮蛋白凝膠結構更加疏松,可以明顯觀察到凝膠壁間的孔洞,這可能是因為高溫熱處理對蕎麥麩皮蛋白結構的破壞更明顯,使原本包裹在蛋白質分子內部的極性側鏈發生解離,促進蛋白質與水分子的結合,從而使蕎麥麩皮蛋白熱凝膠的微觀結構呈現出孔洞明顯的凝膠壁[33]。

圖4 SPI、TBBP與CBBP及其熱凝膠的微觀結構Fig.4 Microstructure of SPI,TBBP,CBBP and their thermally induced gels
2.6 熱特性結果分析
表2 為SPI、TBBP 與CBBP熱變性的起始溫度(To)、峰值溫度(Td)、終止溫度(Tf)和焓值(ΔH)。Td表示蛋白質從天然狀態轉變為變性狀態時的溫度,可反映出蛋白質的熱穩定性,表2中TBBP與CBBP的Td分別為78.38 ℃與77.57 ℃,顯著低于SPI(80.93 ℃),表明SPI有更強的熱穩定性,可能與其加熱后形成更緊密的微觀結構有關。To和Tf的結果規律與Td一致,均為SPI>TBBP>CBBP,進一步表明高溫條件下SPI的結構更穩定。熱變性焓值是使蛋白質的分子結構從有序向無序、從折疊向展開轉變的過程中所需要的熱量,其值與蛋白質結構的有序程度和聚集性有關[34]。如表2所示,3 種蛋白質的ΔH大小依次為SPI>TBBP>CBBP,但無顯著差異(P>0.05),表明TBBP和CBBP完全變性所需要的能量小于SPI,這可能是由于SPI中二硫鍵含量更高,打破二硫鍵使蛋白質發生解折疊形成凝膠所需的熱量更多[35]。

表2 SPI、TBBP與CBBP的熱變性溫度與焓值Table 2 Thermal denaturation temperatures and enthalpy of SPI,TBBP and CBBP
2.7 流變學特性結果分析
如圖5A所示,當溫度恒定為25 ℃時,3 種蛋白的G’與G”值始終為SPI>CBBP>TBBP,且CBBP與TBBP遠小于SPI,這與SPI有較高的黏性有關。當溫度從25 ℃升高至90 ℃的過程中,SPI的G’與G”值隨時間延長不斷降低,而CBBP與TBBP的G’與G”值則先降低后升高,這是由于起始階段,隨溫度緩慢升高,蛋白質溶液黏性逐漸下降,流體特性偏向于液態,從而使蕎麥麩皮蛋白溶液G’與G”值降低。當溫度升高至55 ℃左右,CBBP與TBBP分子開始相互聚集,結構改變形成凝膠,流體特性由液態偏向于固態,G’和G”值進而顯著增加。在溫度升高至90 ℃并處于保溫階段時,3 種蛋白質溶液的G’與G”值隨時間延長均趨于穩定狀態,這是因為溫度過高導致蛋白質發生變性,并且在氫鍵或疏水相互作用下形成相對穩定的凝膠結構。在降溫階段,3 種蛋白溶液的G’與G”值均繼續升高,并且在最高值保持穩定,這是因為在降溫過程中,蛋白質分子的氫鍵與分子間相互作用力進一步增強,從而展現出更高的G’和G”值[36]。
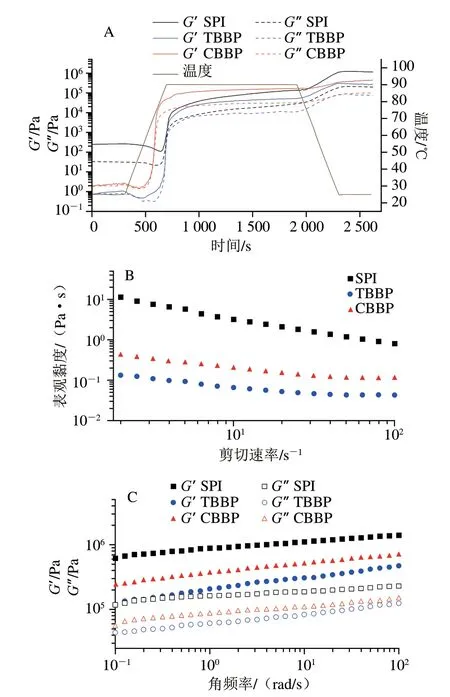
圖5 SPI、TBBP與CBBP的流變學特性變化曲線Fig.5 Rheological curves of SPI,TBBP and CBBP
如圖5B所示,隨著剪切速率的增大,3 種蛋白質溶液的表觀黏度均明顯降低,呈現出非牛頓流體特性。而在相同剪切速率下,CBBP與TBBP的表觀黏度顯著低于SPI,這歸因于SPI的分子質量較高,分子鏈較長,更容易卷曲纏連,分子間移動受阻,不易流動,使得在相同剪切速率下有較高的表觀黏度[37]。當剪切速率大于10 s-1時,CBBP與TBBP的表觀黏度隨剪切速率的增加而緩慢降低并逐漸趨于穩定,這是因為隨剪切速率增大,蛋白質分子間卷曲連接的結構會被剪切為無規則排列的線團或小顆粒,當蛋白質分子呈軸向排列時,表觀黏度減小的趨勢逐漸趨于平緩[38]。
圖5C為SPI、TBBP與CBBP的頻率掃描曲線圖。從整體看,SPI、TBBP與CBBP的G’和G”值均隨角頻率增加而緩慢增加,呈頻率依賴性,且G’始終大于G”,這說明SPI、TBBP與CBBP中分子間產生較強的交聯作用,使結合力增強,從而獲得更高的模量,形成弱凝膠。在同一角頻率下,3 種蛋白溶液中G’與G”值均為SPI>CBBP>TBBP,這與蛋白質的聚集程度有關,由SPI的微觀結構可知(圖4),SPI分子多呈不規則球狀聚集體,部分蛋白質顆粒由于發生變性結構展開并將小顆粒蛋白質包裹,使分子間相互纏結聚集,形成較強的網絡結構,從而使SPI表現出較強的凝膠特性[39]。
3 結論
本研究通過對蕎麥麩皮蛋白分子質量、二級結構、內源熒光光譜、游離巰基與二硫鍵等結果進行分析,探究熱處理對其結構與理化特性的影響。SDS-PAGE結果顯示,熱處理使TBBP與CBBP的部分亞基被降解為小分子質量的多肽或寡肽,使條帶主要分布在低分子質量區域。二級結構結果表明,熱處理后TBBP與CBBP的二級結構由有序的α-螺旋結構轉變為相對無序的β-折疊、β-轉角和無規卷曲結構。由內源熒光光譜結果可知,熱處理后TBBP與CBBP的最大發射波長發生紅移,熒光強度降低,并且游離巰基與二硫鍵含量顯著增加。TBBP-G與CBBP-G的微觀結構觀察結果表明,熱處理后蕎麥麩皮蛋白表面更加粗糙,不規則片狀聚集體略有疏松,孔洞與裂縫增多。與SPI相比,TBBP與CBBP的Td與ΔH更低。且在相同剪切速率與角頻率下,CBBP與TBBP的表觀黏度與G’、G”值均顯著小于SPI。綜上所述,熱處理降低了蕎麥麩皮蛋白的結構穩定性,促使理化性質發生不同程度的變化。本研究為深入了解蕎麥麩皮蛋白的熱加工特性,闡明其在熱加工過程中的功能組分結構、理化特性等變化規律奠定理論基礎,為擴大其在熱加工制造領域的開發與利用提供參考。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
