基于梯度降溫的草魚暫養(yǎng)及有水保活運(yùn)輸技術(shù)
2024-03-10 11:25:16何靜怡岑劍偉郝淑賢陳勝軍趙永強(qiáng)王悅齊楊少玲
食品科學(xué) 2024年4期
何靜怡,魏 涯,岑劍偉,3, ,郝淑賢,陳勝軍,黃 卉,趙永強(qiáng),王悅齊,楊少玲,林 織
(1.上海海洋大學(xué)食品學(xué)院,上海 201306;2.中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所,農(nóng)業(yè)農(nóng)村部水產(chǎn)品加工重點(diǎn)實(shí)驗(yàn)室,廣東 廣州 510300;3.中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所熱帶水產(chǎn)研究開發(fā)中心,海南 三亞 572019;4.廣東順欣海洋漁業(yè)集團(tuán)有限公司,廣東 陽江 529800)
我國(guó)消費(fèi)者對(duì)“生猛海鮮”的意識(shí)根深蒂固,鮮活水產(chǎn)品的售價(jià)比冷凍產(chǎn)品高,經(jīng)濟(jì)類活魚售價(jià)是冷凍或冷藏魚的2~5 倍,鮮活水產(chǎn)品的長(zhǎng)途保活運(yùn)輸是實(shí)現(xiàn)產(chǎn)品升值的有效途徑[1-2],因此保活運(yùn)輸是鮮活水產(chǎn)品市場(chǎng)流通的核心關(guān)鍵技術(shù)。目前對(duì)運(yùn)輸應(yīng)激因素的考慮更加全面,多種生理生化指標(biāo)的測(cè)量及組學(xué)技術(shù)的應(yīng)用使得魚體的應(yīng)激響應(yīng)機(jī)制逐漸清晰[3],有水運(yùn)輸中水質(zhì)及魚類應(yīng)激行為的實(shí)時(shí)監(jiān)測(cè)技術(shù)已實(shí)現(xiàn)遠(yuǎn)程化、智能化[4-5],活運(yùn)技術(shù)的進(jìn)步改善了我國(guó)水產(chǎn)品產(chǎn)銷及分布不均衡等問題。草魚(Ctenopharyngodon idellas)作為我國(guó)四大家魚之一,是重要的經(jīng)濟(jì)魚類,因自身具有廣溫性、易訓(xùn)食、長(zhǎng)速快的特點(diǎn)使其養(yǎng)殖量常年穩(wěn)居首位[6],2021年我國(guó)草魚總產(chǎn)量為557.51萬 t,居全國(guó)淡水養(yǎng)殖水產(chǎn)品總量第一[7]。溫度是影響活魚運(yùn)輸?shù)闹匾蛩兀线m的低溫環(huán)境可有效減慢代謝,維持運(yùn)輸過程中的良好水質(zhì)和減少能量消耗,Santos等[8]研究水溫對(duì)江尾海鰻幼魚的麻醉效果,發(fā)現(xiàn)誘導(dǎo)和復(fù)蘇時(shí)間隨溫度升高而降低,且不受體質(zhì)量影響;Chen Chengzhuang等[9]研究高溫脅迫下不同飼養(yǎng)策略對(duì)鯉魚各項(xiàng)血清生化指標(biāo)的影響,發(fā)現(xiàn)間歇喂養(yǎng)方式可減小其高溫應(yīng)激的體內(nèi)氧化反應(yīng);這些研究均揭示了溫度對(duì)魚類的重要性。此外,冷馴是活魚運(yùn)輸中必不可少的步驟,作為暫養(yǎng)的關(guān)鍵步驟之一,降溫速率將直接影響魚體健康進(jìn)而影響企業(yè)經(jīng)濟(jì)效益。目前關(guān)于草魚保活運(yùn)輸中溫度及降溫速率的研究尚少,本實(shí)驗(yàn)探索降溫速率對(duì)草魚存活時(shí)長(zhǎng)、生化指標(biāo)等的影響,以期為制定草魚暫養(yǎng)及有水運(yùn)輸規(guī)范、實(shí)現(xiàn)草魚長(zhǎng)途有水保活運(yùn)輸提供理論和數(shù)據(jù)支持。
1 材料與方法
1.1 材料與試劑
草魚購(gòu)自廣東省廣州市蓮湖水產(chǎn)養(yǎng)殖場(chǎng),體質(zhì)量為(1200±100)g,體長(zhǎng)(43±2)cm,無體表傷,眼球清澈,鱗片完整。實(shí)驗(yàn)用魚置于提前曝氣的水中暫養(yǎng)2 h以緩解運(yùn)輸及操作應(yīng)激,期間保證供氧充足。
谷丙轉(zhuǎn)氨酶(alanine aminotransferase,ALT)、谷草轉(zhuǎn)氨酶(aspartate aminotransferase,AST)、堿性磷酸酶(alkaline phosphatase,AKP)、超氧化物歧化酶(superoxide dismutase,SOD)、過氧化氫酶(catalase,CAT)、谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)、丙二醛(malondialdehyde,MDA)、葡萄糖檢測(cè)試劑盒 南京建成生物工程研究所有限公司;魚皮質(zhì)醇(cortisol,COR)酶聯(lián)免疫吸附測(cè)定試劑盒 北京萊伯沃德科技有限公司;氨氮標(biāo)準(zhǔn)物質(zhì)、納氏試劑北京金智研生物科技有限公司。
1.2 儀器與設(shè)備
風(fēng)冷式冷水機(jī) 蘇州肯道節(jié)能設(shè)備有限公司;Multi 3630 IDS便攜式多參數(shù)水質(zhì)分析儀 德國(guó)WTW公司;Synergy全功能酶標(biāo)儀 美國(guó)BioTek公司;UV2550紫外-可見分光光度計(jì) 日本島津公司;HWS-24恒溫水浴鍋上海一恒科學(xué)儀器有限公司;高速臺(tái)式冷凍離心機(jī)湖南湘儀實(shí)驗(yàn)室儀器開發(fā)有限公司;SPX型智能生化培養(yǎng)箱 寧波江南儀器廠。
1.3 方法
1.3.1 草魚休眠臨界溫度
利用實(shí)驗(yàn)室自制梯度降溫及暫養(yǎng)裝置(圖1),按照袁圓玥等[10]的降溫方式進(jìn)行梯度降溫實(shí)驗(yàn):當(dāng)溫度(T)>25 ℃時(shí),以3 ℃/h降溫;當(dāng)10 ℃<T≤25 ℃,以2 ℃/h降溫;當(dāng)T≤10 ℃時(shí),以1 ℃/h降溫。將緩解運(yùn)輸應(yīng)激后的9 尾草魚放入預(yù)先曝氣24 h的清水中,水中溶氧大于8.5 mg/L、pH 7.5±0.2、暫養(yǎng)密度為1∶20(kg/L)。記錄草魚在不同溫度下的呼吸頻率及活動(dòng)現(xiàn)象,鰓蓋一張一合記為一次呼吸,統(tǒng)計(jì)1 min的呼吸次數(shù)[11],并記錄不同溫度下魚體對(duì)光照(室內(nèi)自然光)、敲擊箱體產(chǎn)生的振動(dòng)聲、物體輕觸體表等反應(yīng)的靈敏度。
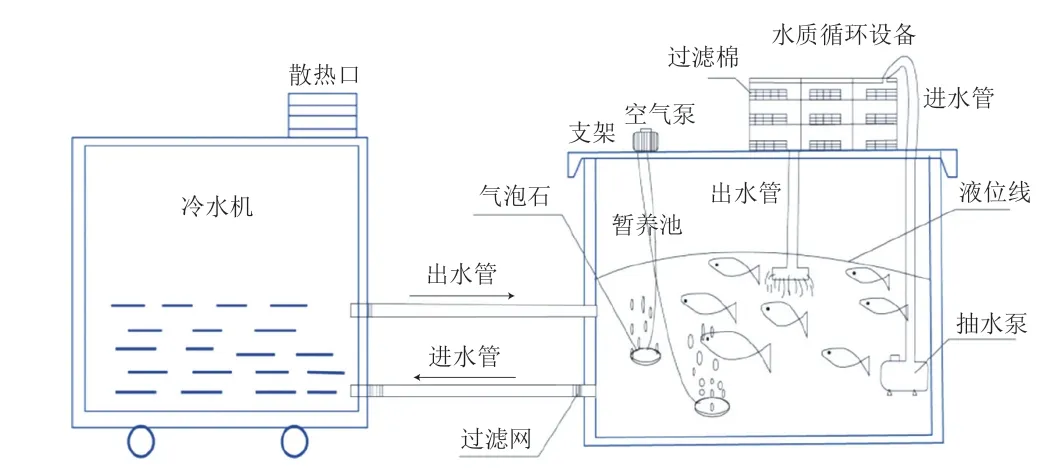
圖1 實(shí)驗(yàn)室梯度降溫及暫養(yǎng)裝置Fig.1 Schematic diagrams of gradient cooling and temporary care devices used in the laboratory
1.3.2 冷馴降溫處理
以梯度降溫方式(10 ℃<T≤25 ℃,以2 ℃/h降溫)[10]將暫養(yǎng)箱水溫從室溫(23~25 ℃)分別降至24、20、18、16、14、12 ℃,每個(gè)溫度記錄6 尾魚,每4 h觀察草魚存活狀態(tài),每12 h采集水樣,參考HJ 535—2009《水質(zhì) 氨氮的測(cè)定 納氏試劑分光光度法》[12]的方法測(cè)定氨氮濃度。
由實(shí)驗(yàn)結(jié)果可知,16 ℃為草魚的半休眠溫度,分別以1、3、5、7 ℃/h將暫養(yǎng)水箱溫度從室溫(23~25 ℃)降溫至16 ℃后,快速將草魚轉(zhuǎn)移至pH 7.5±0.2、溶氧大于8.5 mg/L、魚-水比為1∶20(kg/L)的保活水箱中,冷水機(jī)控溫,保證水溫為(16.0±0.1)℃,保活期間室內(nèi)無光照,空氣泵持續(xù)打氧。設(shè)置3 組平行,每組15 尾魚,分別在保活0、12、24、36、48 h時(shí)各取3 尾魚。以室溫條件下(不做降溫處理)保活的草魚為對(duì)照組,魚-水比、其他水環(huán)境參數(shù)及光照同實(shí)驗(yàn)組。
1.3.3 血清制備
分別在保活0、12、24、36、48 h時(shí)各取3 尾魚采血。參考袁仲瑾等[11]的方法,用一次性無菌針管從草魚尾部靜脈取血,全血不加抗凝劑,37 ℃恒溫生化箱中靜置0.5 h,待血液凝固分層后,以4 ℃、4000 r/min離心10 min,取血清于-80 ℃保存?zhèn)溆茫謩e取對(duì)照組保活0、12、24、36、48 h的草魚血液,血清制備方法同實(shí)驗(yàn)組。
1.3.4 血清生化指標(biāo)測(cè)定
用檢測(cè)試劑盒測(cè)定ALT活力、AST活力、AKP活力、COR質(zhì)量濃度、血糖(glucose,GLU)濃度。
1.3.5 抗氧化應(yīng)激指標(biāo)測(cè)定
用檢測(cè)試劑盒測(cè)定SOD活力、GSH-Px活力、CAT活力、MDA含量。
1.4 數(shù)據(jù)處理與分析
2 結(jié)果與分析
2.1 草魚休眠臨界溫度確定
魚類等變溫動(dòng)物都有一個(gè)能區(qū)分生死的溫度,稱為臨界溫度[13],在有水保活過程中,合適的低溫環(huán)境可降低魚體呼吸頻率和代謝水平,在無水保活過程中,可將環(huán)境溫度降低至魚類的生態(tài)冰溫區(qū)使其進(jìn)入休眠狀態(tài)[14]。由表1可知,草魚的應(yīng)激靈敏度和呼吸頻率隨溫度降低逐漸下降。當(dāng)溫度高于28 ℃時(shí),呼吸急促,出現(xiàn)浮頭或撞壁等應(yīng)激行為;在20~26 ℃時(shí),魚體應(yīng)激敏感,鰓蓋張合規(guī)律,魚體正常游動(dòng);溫度降至18 ℃時(shí),呼吸頻率明顯降低,對(duì)敲擊箱體的振動(dòng)聲和光照反應(yīng)不明顯,對(duì)觸碰刺激仍保持敏感;降至16 ℃時(shí),應(yīng)激反應(yīng)遲鈍,全部靜止在水底,鰓蓋張合幅度小;降溫至8 ℃時(shí),鰓蓋半閉合,將魚體放回室溫水中5~8 min內(nèi)復(fù)蘇;降至6 ℃時(shí),魚體出現(xiàn)裂鰓,根據(jù)蔡曉芳[13]和袁仲瑾[11]等關(guān)于臨界溫度的判斷方法,以出現(xiàn)呼吸極不規(guī)律、觀察不到鰓蓋張合甚至出現(xiàn)裂鰓時(shí)的溫度作為臨界溫度,故認(rèn)為6 ℃為草魚的運(yùn)輸臨界溫度,16 ℃的魚體應(yīng)激不敏感,靜止在水底,是適合草魚運(yùn)輸?shù)陌胄菝邷囟取?/p>
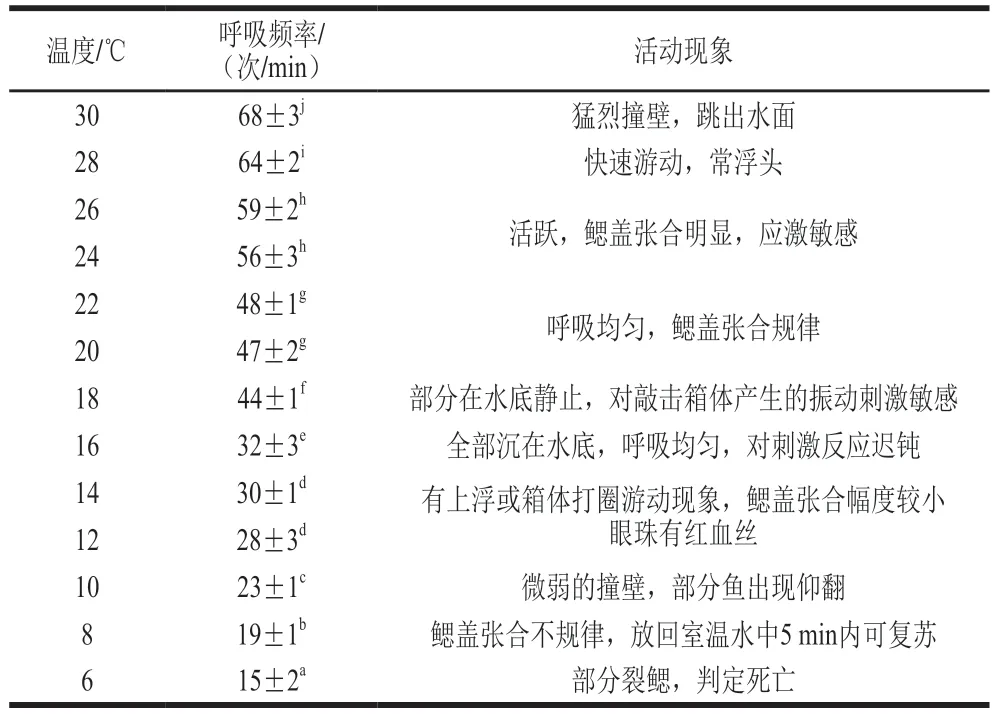
表1 不同溫度下草魚的呼吸頻率和活動(dòng)現(xiàn)象Table 1 Respiratory rates and activity phenomena of grass carp at different temperatures
2.2 不同溫度條件下的水質(zhì)總氨氮質(zhì)量濃度變化及草魚生存函數(shù)曲線
水質(zhì)對(duì)魚類存活具有重要意義,溫度、氨氮濃度等是魚類存活的重要影響因素[15],Zhao Congcong等[16]研究不同濃度的總氨氮質(zhì)量濃度對(duì)幼年草魚存活80 d后各器官特異性的影響,發(fā)現(xiàn)低質(zhì)量濃度總氨氮有利于促進(jìn)和維持草魚苗生長(zhǎng),當(dāng)總氨氮質(zhì)量濃度達(dá)到9.0 mg/L時(shí),肌肉細(xì)胞開始表現(xiàn)出毒性,不同器官的抗氧化系統(tǒng)反應(yīng)不同,其中鰓更易受到低濃度氨氮的影響。當(dāng)水溫大于28 ℃和小于12 ℃時(shí)會(huì)引起魚體不良生理反應(yīng),這與周鑫[17]研究的溫度脅迫結(jié)果一致,當(dāng)溫度超過26 ℃時(shí)草魚各組織中熱應(yīng)激蛋白70(heat stress protein 70,hsp70)和hsp90的表達(dá)量均顯著上升,說明溫度升高導(dǎo)致魚體內(nèi)蛋白質(zhì)變性,而溫度高于28 ℃時(shí),草魚的耗氧率隨溫度升高而下降。由圖2可知,水質(zhì)氨氮質(zhì)量濃度隨保活時(shí)間延長(zhǎng)和溫度升高持續(xù)增加,保活相同時(shí)間下,24 ℃水溫中氨氮質(zhì)量濃度最高,該溫度下保活48 h后的水質(zhì)總氨氮質(zhì)量濃度為23.70 mg/L,而該保活時(shí)間下12、14、16 ℃的總氨氮質(zhì)量濃度顯著降低,分別為8.23、8.27、8.73 mg/L,這與黃蓋鰈、梭鱸的排氨率隨溫度降低而降低的研究結(jié)果[18-19]一致,表明低溫可有效降低草魚的新陳代謝,減緩水體中氨氮的富集速度。硬骨魚的排泄產(chǎn)物主要由氨和尿素組成,其中氨氮占排泄物總量的80%[20],魚作為變溫動(dòng)物,溫度可直接影響其新陳代謝速率和蛋白質(zhì)利用率。有研究表明,魚類的氨氮排泄率與溫度呈正相關(guān)[21-22]。圖3為不同溫度下草魚的生存函數(shù)曲線,16 ℃下草魚的存活率最高,存活時(shí)間最長(zhǎng),66.7%的草魚存活時(shí)長(zhǎng)超過70 h,而12、14、24 ℃僅有33.3%存活時(shí)長(zhǎng)超過50 h,結(jié)合存活時(shí)長(zhǎng)、總氨氮質(zhì)量濃度及草魚的活動(dòng)情況,認(rèn)為16 ℃是草魚進(jìn)行暫養(yǎng)及有水保活運(yùn)輸?shù)耐扑]溫度。
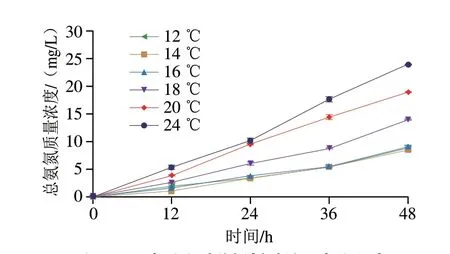
圖2 溫度對(duì)水質(zhì)總氨氮質(zhì)量濃度的影響Fig.2 Effect of temperature on the concentration of total ammonia nitrogen in water
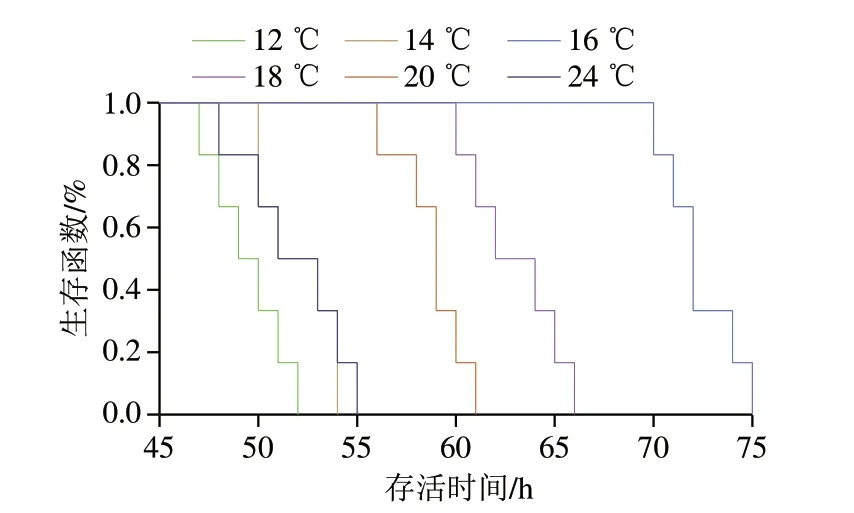
圖3 溫度對(duì)草魚存活時(shí)間的影響Fig.3 Effect of temperature on the survival time of grass carp
2.3 降溫速率對(duì)草魚血清生化指標(biāo)的影響
由圖4A~C可知,1、5 ℃/h降溫處理組在保活48 h后ALT、AST、AKP水平均高于3 ℃/h組和對(duì)照組,魚體承受的應(yīng)激損傷隨時(shí)間延長(zhǎng)逐漸加重;以3 ℃/h速率降溫時(shí),魚體的AST及AKP活力在不同保活時(shí)間點(diǎn)達(dá)最大值后下降,表明在此條件下,魚體能較快完成自身調(diào)節(jié),在低溫環(huán)境下重新達(dá)到穩(wěn)態(tài)平衡。3 ℃/h組AST活力在保活12 h時(shí)達(dá)最大值(143.75 U/L),保活48 h下降為102.01 U/L,低于相同保活時(shí)長(zhǎng)下其他降溫處理組;各組的ALT活力整體上隨保活時(shí)間延長(zhǎng)逐漸升高,1、5 ℃/h降溫處理組保活48 h后ALT活力分別為3 ℃/h組的1.5、1.8 倍;3 ℃/h組AKP活力隨保活時(shí)間延長(zhǎng)先增加后降低,1、3、5 ℃/h組最大值分別為48.74、45.53、60.55 U/L,是對(duì)照組的2.1、2.4 倍和3.0 倍,5 ℃/h組的AKP活力最高,影響最大。由圖4D、E可知,3 個(gè)降溫處理組保活12、24、36 h的COR及GLU水平均顯著高于對(duì)照組;3 ℃/h降溫處理組保活48 h后的GLU含量顯著低于1、5 ℃/h組。1 ℃/h降溫處理組保活0 h時(shí),COR和GLU水平均顯著高于其他組別;由于在1 ℃/h降溫速率下,達(dá)到半休眠溫度所需的冷馴時(shí)間長(zhǎng),魚體處于水溫變化的時(shí)間長(zhǎng),GLU濃度隨時(shí)間延長(zhǎng)先增加后降低,與COR質(zhì)量濃度變化趨勢(shì)一致;1、3、5 ℃/h降溫處理組GLU濃度分別在保活12、24 h和36 h達(dá)最大值,為6.10、5.52、7.18 mmol/L,保活48 h后GLU濃度均有所下降,但仍高于對(duì)照組GLU水平。對(duì)比對(duì)照組與降溫處理組的5 項(xiàng)血清生化指標(biāo)可得,降溫操作使魚體產(chǎn)生應(yīng)激,其中5 ℃/h降溫速率快,水溫短時(shí)間內(nèi)快速降低造成魚體強(qiáng)烈應(yīng)激,COR質(zhì)量濃度迅速升至較高水平應(yīng)對(duì)刺激,保證機(jī)體的正常生命活動(dòng),相比于長(zhǎng)時(shí)間的慢速降溫,短時(shí)間的快速降溫使魚體的應(yīng)激更強(qiáng)烈,而1 ℃/h組降溫至16 ℃所需時(shí)間長(zhǎng),進(jìn)入半休眠狀態(tài)晚于3、5 ℃/h組,水溫長(zhǎng)時(shí)間變化導(dǎo)致魚體產(chǎn)生持續(xù)應(yīng)激反應(yīng),因此以1、5 ℃/h的速率降至16 ℃后進(jìn)行保活使魚體的應(yīng)激比3 ℃/h更強(qiáng)烈,損傷更大。
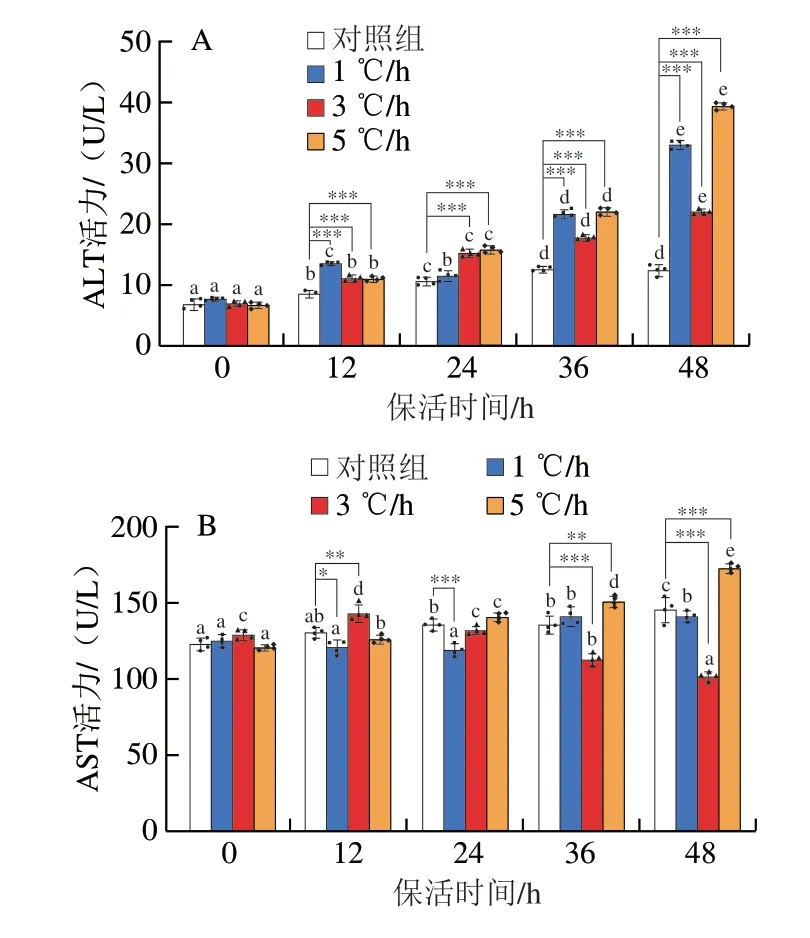
圖4 降溫速率對(duì)草魚ALT(A)、AST(B)、AKP(C)、COR(D)、GLU(E)的影響Fig.4 Effect of cooling rate on ALT (A),AST (B),AKP (C),COR (D),and GLU (E) in grass carp
2.4 降溫速率對(duì)草魚抗氧化能力的影響
SOD是魚體的主要抗氧化物酶,作為機(jī)體抵御氧化應(yīng)激的首道防線,其消除氧化脅迫的效果顯著,SOD活力能間接反映機(jī)體清除自由基的能力[23],由圖5A可知,相比于對(duì)照組,降溫處理組的SOD活性均顯著增加,隨保活時(shí)間延長(zhǎng)先增加后降低,1、3、5 ℃/h降溫處理組均在保活36 h達(dá)最大值,分別為260.49、276.23、315.01 U/mg,是對(duì)照組保活相同時(shí)長(zhǎng)的1.9、2.1、2.4 倍。CAT是清除自由基重要的抗氧化酶,3、5 ℃/h降溫處理組的CAT活力保活24 h達(dá)最大值(7.06、6.92 U/mg),隨后逐漸下降;1 ℃/h降溫處理組的CAT活力隨保活時(shí)間延長(zhǎng)逐漸升高,保活48 h后與對(duì)照組相比無顯著差異,均低于3、5 ℃/h組(圖5B)。GSH-Px是抗氧化系統(tǒng)中的酶類抗氧化劑,能清除因自由基氧化所產(chǎn)生的脂質(zhì)過氧化物,降解H2O2,保護(hù)細(xì)胞膜結(jié)構(gòu)及功能不受過氧化物的干擾及損害[20]。由圖5C可知,3 ℃/h組的GSH-Px活力隨保活時(shí)間延長(zhǎng)先增加后降低;保活48 h的1 ℃/h組GSH-Px活力高于3、5 ℃/h降溫處理組和對(duì)照組,且3 ℃/h降溫處理組的GSH-Px活力恢復(fù)至初始狀態(tài);保活48 h后1、5 ℃/h處理組GSH-Px活力是3 ℃/h組的1.2、1.1 倍。據(jù)劉奇奇[24]研究操作及低溫脅迫對(duì)四指馬鲅的抗氧化系統(tǒng)結(jié)果可知,魚體在應(yīng)對(duì)低溫應(yīng)激時(shí),肝臟和肌肉等不同組織中的GSHPx含量不同,肝臟中含量更高,GSH-Px活性可間接反映肝臟損傷狀態(tài),酶活性的升高說明活性氧自由基含量增加,表明1、5 ℃/h條件對(duì)魚體的氧化應(yīng)激更嚴(yán)重。根據(jù)Kim等[25]研究魚類氧化應(yīng)激的結(jié)果可知,較高的SOD、CAT、GSH-Px活性分別與體內(nèi)過高的O2-·、H2O2和活性氧(reactive oxygen species,ROS)水平有關(guān)。SOD和GSH-Px活性在快、慢速降溫處理組中均較高,表明在降溫過程中,產(chǎn)生較多的O2-·和ROS,上述兩種酶在清除和抵御ROS導(dǎo)致的氧化應(yīng)激反應(yīng)中發(fā)揮重要作用,這與羅偉等[26]的研究結(jié)果相似,在溫度應(yīng)激下魚血清中GSH-Px活性較高。由圖5D可知,1 ℃/h降溫處理組的MDA含量隨保活時(shí)間延長(zhǎng)先升高后降低再升高,在24 h時(shí)降至最低,3、5 ℃/h組均在保活36 h降至最小值5.27、6.02 nmol/mg;1、3、5 ℃/h降溫處理組保活48 h的MDA含量分別是對(duì)照組保活48 h的1.9、1.4 倍和1.7 倍,可見1、5 ℃/h降溫處理組魚體的氧化應(yīng)激強(qiáng)烈,ROS含量增多,從而加深氧化應(yīng)激反應(yīng)程度。降溫處理組的SOD、CAT及GSH-Px活力基本上隨保活時(shí)間延長(zhǎng)先升高后降低,表明降溫導(dǎo)致魚體內(nèi)自由基失衡,誘導(dǎo)體內(nèi)發(fā)生氧化反應(yīng),3 ℃/h降溫處理組保活48 h后,酶活性恢復(fù)至初始狀態(tài),而過快或過慢降溫對(duì)魚體組織均會(huì)造成不同程度損傷,不能及時(shí)且準(zhǔn)確地調(diào)節(jié)抗氧化酶的含量達(dá)到穩(wěn)態(tài)環(huán)境,1、5 ℃/h降溫處理組對(duì)魚體的線粒體數(shù)量和功能的影響遠(yuǎn)大于3 ℃/h組,進(jìn)而影響ROS的產(chǎn)生。
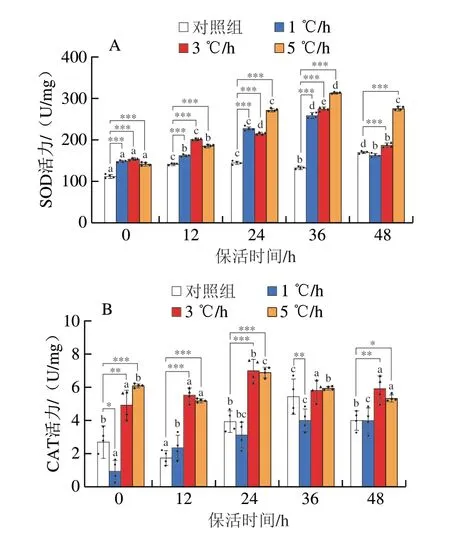
圖5 降溫速率對(duì)草魚SOD活性(A)、CAT活性(B)、GSH-Px活性(C)、MDA含量(D)的影響Fig.5 Effect of cooling rate on SOD (A),CAT (B),GSH-Px (C),and MDA (D) levels in grass carp
3 討論
3.1 草魚的臨界溫度確定
本實(shí)驗(yàn)中,溫度變化引起草魚應(yīng)激反應(yīng),溫度升高草魚代謝加快,應(yīng)激敏感,對(duì)水體的污染加重,溫度過低產(chǎn)生低溫脅迫,眼球出現(xiàn)紅血絲等不良生理反應(yīng)。在草魚的有水保活運(yùn)輸中,水溫不應(yīng)高于28 ℃或低于6 ℃。通過觀察12~24 ℃范圍內(nèi)草魚的存活情況及水質(zhì)總氨氮質(zhì)量濃度發(fā)現(xiàn),隨溫度升高,氨氮質(zhì)量濃度顯著增加,存活時(shí)間延長(zhǎng)先增加后減少。這和朱祥宇等[27]研究不同溫度條件下草魚苗的排氨率結(jié)果相似,在24 ℃水中排氨率達(dá)到最大值(18.99 mg/h),15 ℃達(dá)到最小值(7.53 mg/h),草魚的耗氧率、排糞率均隨溫度升高而增加,呼吸代謝增強(qiáng),水質(zhì)污染速率加快,心肌興奮性加強(qiáng),過高的溫度將導(dǎo)致呼吸活動(dòng)和心臟功能紊亂。因此,當(dāng)溫度高于28 ℃或低于6 ℃時(shí)不利于草魚的保活運(yùn)輸,在16 ℃時(shí)草魚進(jìn)入半休眠狀態(tài),應(yīng)激反應(yīng)遲鈍,存活時(shí)間最長(zhǎng),總氨氮含量最低,是進(jìn)行運(yùn)輸?shù)睦硐霚囟取?/p>
3.2 不同降溫速率對(duì)草魚應(yīng)激反應(yīng)的影響
魚類在進(jìn)行長(zhǎng)途運(yùn)輸過程中,常受到來自外界的壓力,其生理應(yīng)激從激活交感神經(jīng)反應(yīng)開始,刺激垂體分泌促腎上腺皮質(zhì)激素,再由腎間組織釋放以COR為主的皮質(zhì)類固醇,參與應(yīng)激反應(yīng)的被動(dòng)調(diào)節(jié)[28],COR刺激下游生理過程(新成代謝和免疫反應(yīng))應(yīng)對(duì)環(huán)境變化,進(jìn)一步影響動(dòng)物的行為、生長(zhǎng)和生存,高COR水平與食物攝入量、食物轉(zhuǎn)化效率、能量消耗和生長(zhǎng)性能呈負(fù)相關(guān)[29]。葡萄糖含量的升高主要是由兒茶酚胺和COR分別激活肝臟糖異生和肌肉組織糖原分解引起,葡萄糖通過影響胰島素的分泌進(jìn)而影響蛋白質(zhì)分解和脂肪氧化[29-31],魚體內(nèi)的血糖由于肝臟糖原代謝和糖異生反應(yīng),短期內(nèi)出現(xiàn)“高血糖”癥狀[10],隨保活時(shí)間延長(zhǎng),該癥狀得以緩解。不同降溫速率下COR和GLU含量隨時(shí)間延長(zhǎng)先增加后降低,均高于對(duì)照組,表明降溫引起魚體應(yīng)激,保活24 h內(nèi)COR質(zhì)量濃度和GLU濃度升高,48 h后兩指標(biāo)均有所下降,甚至與保活初期無顯著差異,表明48 h后魚體可通過調(diào)節(jié)體內(nèi)各系統(tǒng)適應(yīng)新環(huán)境,亦表明COR及GLU濃度僅適合作為短期應(yīng)激魚體的評(píng)估指標(biāo),超過48 h后該指標(biāo)的準(zhǔn)確度降低。與1、5 ℃/h降溫處理組相比,3 ℃/h降溫處理組的COR質(zhì)量濃度和GLU濃度最低,保活48 h后基本恢復(fù)初始水平,表明以3 ℃/h的降溫速率進(jìn)行冷馴時(shí)魚體的應(yīng)激反應(yīng)程度小。
3.3 不同降溫速率對(duì)草魚肝臟的影響
肝臟是魚體內(nèi)最大的腺體器官,具有代謝、營(yíng)養(yǎng)吸收、免疫等多種功能[32],ALT和AST是肝臟連接糖、脂質(zhì)、蛋白質(zhì)代謝的重要酶,作為評(píng)價(jià)肝臟病變程度最可靠的指標(biāo),當(dāng)魚體未受到強(qiáng)烈應(yīng)激時(shí)2 種酶活性較低[33],其中,ALT主要存在于肝臟中,AST除存在于肝臟外,還存在于心肌細(xì)胞中,因此AST活力亦能反映心肌細(xì)胞受損傷程度[34]。AKP是重要的磷酸單酯水解酶和非特異性磷酸酶,能催化磷酸單酯水解反應(yīng),同時(shí)在消解病原體、細(xì)胞吞噬、蛋白質(zhì)磷酸基團(tuán)轉(zhuǎn)移、鈣磷代謝等過程中發(fā)揮重要作用,可衡量魚體非特異性免疫能力水平,在非必需氨基酸的合成和蛋白質(zhì)分解代謝中發(fā)揮重要的中間作用[35-37]。保活48 h后1、5 ℃/h降溫處理組的AST和ALT活力均高于3 ℃/h組,表明1、5 ℃/h的降溫速率對(duì)心肌和肝臟的損傷更顯著,這與華茂圳等[38]研究降溫速率對(duì)斑石鯛無水保活的結(jié)果一致,其認(rèn)為3 ℃/h冷馴后無水保活6 h對(duì)斑石鯛產(chǎn)生的應(yīng)激反應(yīng)最小,本實(shí)驗(yàn)中以3 ℃/h對(duì)草魚進(jìn)行冷馴,降溫梯度小、時(shí)間短,魚體可快速進(jìn)入半休眠狀態(tài)。在應(yīng)激過程中,ROS過量產(chǎn)生會(huì)增加溶酶體膜的通透性和溶酶體蛋白水解酶向胞漿的釋放,其中AKP是溶酶體的主要酶之一[36],其活性可反映溶酶體狀態(tài),進(jìn)而反映魚體的非特異性免疫能力。3 ℃/h組保活36 h的AKP活力下降,表明魚體隨保活時(shí)間延長(zhǎng)在調(diào)節(jié)自身免疫,達(dá)到新的平衡狀態(tài)。1 ℃/h組的AKP活力持續(xù)增加,表明長(zhǎng)時(shí)間的冷馴過程也對(duì)魚體免疫能力造成損害;保活48 h后5 ℃/h組的AKP活力高于3 ℃/h,快速降溫會(huì)損傷溶酶體膜,對(duì)魚體的免疫系統(tǒng)造成傷害。
3.4 不同降溫速率對(duì)草魚氧化應(yīng)激的影響
ROS的產(chǎn)生和抗氧化防御系統(tǒng)之間缺乏平衡(即氧化應(yīng)激)會(huì)導(dǎo)致DNA羥化、蛋白質(zhì)變性、脂質(zhì)過氧化、細(xì)胞凋亡[39],因此與氧化應(yīng)激相關(guān)的細(xì)胞內(nèi)ROS水平升高會(huì)對(duì)DNA、蛋白質(zhì)和脂質(zhì)造成破壞性影響。其中SOD、CAT、GSH-Px是魚體內(nèi)重要的抗氧化酶系[40],其活性可反映魚體內(nèi)的氧化應(yīng)激程度,SOD主要催化·分解為O2和H2O2,H2O2被CAT和GSH-Px等部分消除,GSH-Px是機(jī)體內(nèi)廣泛存在的一種催化H2O2分解的酶,能特異性地催化還原型谷胱甘肽與H2O2的還原反應(yīng),起到保護(hù)細(xì)胞膜結(jié)構(gòu)和功能完整的作用[40-42]。同時(shí),GSH-Px能夠催化GSH變?yōu)檠趸凸入赘孰模褂卸镜倪^氧化物還原變?yōu)闊o毒的羥基化合物[43]。體內(nèi)ROS的不斷積累誘發(fā)膜脂過氧化,MDA是膜脂過氧化的重要產(chǎn)物之一,其含量反映了細(xì)胞膜過氧化的程度,是判斷脂質(zhì)過氧化程度和組織損傷的指標(biāo)[11]。
緩慢降溫能避免因溫度快速變化導(dǎo)致魚體應(yīng)激,但將延長(zhǎng)魚體進(jìn)入半休眠狀態(tài)的時(shí)間,長(zhǎng)時(shí)間處于水溫變化的環(huán)境中可能導(dǎo)致魚體持續(xù)應(yīng)激,而快速降溫可縮短冷馴時(shí)間,但降溫過快對(duì)魚體組織損傷巨大。本實(shí)驗(yàn)中,7 ℃/h降溫速率下草魚的存活時(shí)長(zhǎng)小于40 h,不利于長(zhǎng)途活運(yùn);5 ℃/h降溫處理組的SOD活性高于其他組別,表明該降溫速率對(duì)魚體的應(yīng)激顯著,產(chǎn)生大量ROS誘導(dǎo)氧化應(yīng)激,縮短存活時(shí)長(zhǎng)。1、5 ℃/h處理組降溫結(jié)束后的MDA含量均顯著高于3 ℃/h,隨保活時(shí)間延長(zhǎng),快速降溫會(huì)損傷魚體肝臟等器官,減少酶的釋放,降低清除自由基的能力,加深脂質(zhì)過氧化程度,升高M(jìn)DA含量。以3 ℃/h降至16 ℃保活48 h后血清生化指標(biāo)及抗氧化酶活力與該降溫速率下保活0 h的差異最小,在該降溫速率下魚體能通過調(diào)節(jié)自身代謝,使得魚體的抗氧化及非特異性免疫能力在低溫下達(dá)到新的平衡狀態(tài),3 ℃/h降溫處理的草魚抗逆性最強(qiáng),這與張玉晗等[44]在無水保活過程中低溫休眠處理花鱸發(fā)現(xiàn)3 ℃/h處理后保活8 h對(duì)花鱸的肝臟損傷最低結(jié)論相似。
4 結(jié)論
本實(shí)驗(yàn)基于梯度降溫研究了草魚的臨界及半休眠溫度,記錄不同溫度條件下草魚的存活時(shí)長(zhǎng)、監(jiān)測(cè)不同溫度下水質(zhì)氨氮含量間接反映溫度對(duì)草魚生理代謝的影響。結(jié)果發(fā)現(xiàn)草魚暫養(yǎng)及有水活運(yùn)的最適溫度為16 ℃,其臨界低溫為6~8 ℃;在進(jìn)行長(zhǎng)途運(yùn)輸時(shí),水溫應(yīng)當(dāng)小于24 ℃。分別以1、3、5、7 ℃/h的降溫速率將暫養(yǎng)箱水溫從室溫(23~25 ℃)降至草魚半休眠溫度(16 ℃)后開展保活,在不同保活時(shí)長(zhǎng)測(cè)量血清各項(xiàng)指標(biāo),得到在暫養(yǎng)冷馴過程中,適合草魚的降溫速率為3 ℃/h,該降溫速率對(duì)草魚體內(nèi)各項(xiàng)生理影響最小,其抗逆性最強(qiáng),適合長(zhǎng)途運(yùn)輸。本研究獲得了草魚適宜的暫養(yǎng)及運(yùn)輸推薦溫度,后續(xù)模擬長(zhǎng)途運(yùn)輸實(shí)驗(yàn)亟待開展,驗(yàn)證經(jīng)過3 ℃/h的降溫速率冷馴后的草魚在不同運(yùn)輸條件的生理變化及對(duì)其品質(zhì)的影響,加快草魚低溫誘導(dǎo)休眠技術(shù)在產(chǎn)業(yè)上的推廣應(yīng)用。
