益景湯對早期糖尿病視網膜病變大鼠視網膜miRNA表達的影響
2024-03-12 06:09:46王麗邱心悅孟春劉光輝
中國中醫眼科雜志 2024年3期
王麗,邱心悅,孟春,劉光輝
糖尿病視網膜病變(diabetic retinopathy,DR)是糖尿病(diabetes mellitus,DM)常見的微血管并發癥,我國作為全球DM 患者人數最多的國家,DR 患病率逐年增加,致盲率也在逐年升高,給社會造成了巨大的經濟負擔[1]。DR 根據病程分為非增殖期和增殖期,增殖性DR 是患者出現視力損害和失明的主要原因。DR 一旦進入增殖期則不可逆轉,因此,對DR 進行積極有效的早期干預是防治DR 的重要策略。中醫藥在DR 的早期防治中扮演了重要的角色[2]。本團隊的前期臨床研究[3-5]發現,益景湯治療早期DR 具有較好的療效,能夠在一定程度上改善患者視網膜微循環、減輕黃斑水腫、調控血管內皮生長因子(vascular endothelial growth factor,VEGF)的水平,基礎研究[6-8]發現,益景湯能夠抑制DM 大鼠視網膜周細胞的凋亡,緩解視網膜毛細血管床的減少,改善視網膜血供,減輕視網膜滲漏。近年來,多項研究[9-12]表明,微小RNA(micro RNA,miRNA)在DR 發生發展及治療方面發揮重要作用。為進一步探討益景湯干預早期DR 的分子機制,本研究擬通過小RNA 測序篩選出可能參與益景湯干預早期DR 發生發展的miRNA,為中醫藥防治早期DR提供新的方法和實驗依據。
1 材料與方法
1.1 實驗動物
30 只SPF 級健康雄性SD 大鼠,2 月齡,體質量(200±20)g,購買自福州諾頓斯生物科技有限公司,許可證號為SYXK(閩)2020-0002,飼養于福建中醫藥大學實驗動物中心。實驗前適應性喂養7 d,自由進食,室溫20~22 ℃,晝夜循環照明。本研究實驗動物及實驗所用條件符合國家科學技術委員會的《實驗動物管理條例》相關規定。
1.2 藥品、試劑與儀器
鏈脲佐菌素(美國Sigma 公司,V900890-100MG),TruSeq 小RNA 樣本制備試劑盒(美國Illumina公司,RS-200-0012),mirVana?miRNA 分離試劑盒(美國Ambion 公司,AM1561),SuperScript Ⅱ逆轉錄酶、Qubit RNA 定量檢測試劑盒(美國Invitrogen 公司,18064-014、Q10211),RNA 分析試劑盒、高靈敏度DNA 芯片(美國Agilent 公司,5067-1511、5067-4626),Qubit DNA 高靈敏度熒光定量試劑盒(美國Invitrogen 公司,Q32854),通用型SYBR快速定量試劑盒、DNA 文庫定量試劑盒(美國KAPA Biosystem 公司,KK4602、KK4808)。2100 生物分析儀(美國Agilent 公司,J06-02),Qubit 2.0 熒光計(美國Invitrogen公司,Q32871),紫外透色儀(美國Clare Chemical Research 公司,D195M),聚合酶鏈式反應(polymerase chain reaction,PCR)熱循環儀(美國MJ公司,B01-01)。
益景湯由生黃芪、山藥、茯苓、生地黃、玄參各15 g,赤芍10 g,蒼術、當歸各9 g,川芎6 g,桃仁、紅花各5 g 組成,中藥材由福建中醫藥大學附屬人民醫院提供。每次取益景湯1 劑,用10 倍體積純凈水浸泡30 min 后煮1.5 h,靜置冷卻過濾,加入圓底燒瓶,使用旋轉蒸發儀濃縮,制成1 g/mL 益景湯藥液,滅菌玻璃瓶灌裝,密封避光4 ℃冰箱保存備用。
1.3 DM模型的建立
20 只動物禁食12 h 后,將鏈脲佐菌素使用0.1 mmol/L、pH 為4.5 的無菌檸檬酸緩沖液溶解,配置成1%溶液后,以70 mg/kg 劑量進行一次性腹腔注射。72 h 后大鼠尾靜脈取血檢測血糖水平,血糖濃度≥16.7 mol/L即視為DM模型制備成功。
1.4 分組與給藥
將造模成功的20 只大鼠采用隨機數字表法分為模型組(model group,MG)和益景湯組(YijingDecoction group,YJG),另將常規喂養的大鼠設為對照組(control group,CG),每組10 只。根據人與大鼠的臨床等效劑量,YJG 組大鼠每日灌胃益景湯12.50 g/kg,并于取材前1 h給藥1次;CG組、MG組大鼠灌胃等體積0.9%氯化鈉注射液,處死前1 h 灌胃1次,3組均干預16周。
1.5 取材
3 組大鼠干預16 周后,記錄體重,麻醉處死,立即摘除雙側眼球,在冰水中剝離視網膜后,置于-80 ℃冰箱中保存,以備小RNA及PCR檢測使用。
1.6 總RNA提取與小RNA通用制備
視網膜組織研磨并分離核酸蛋白后提取RNA,采用紫外分光光度計測定RNA 純度,通過瓊脂糖凝膠電泳檢測RNA 的質量,生物分析儀檢測RNA 完整值(RNA integrity number,RIN)。確定總RNA 合格后,對其進行定量,利用6%聚丙烯酰胺凝膠電泳分離并回收純化22~30 nt范圍內的小RNA。
1.7 小RNA文庫構建和測序
構建小RNA 文庫,具體過程是分別在小RNA加上3'接頭和5'接頭,利用與3'接頭反向互補的引物反轉錄擴增出cDNA,最后進行15 個循環的PCR擴增獲得小RNA 文庫,采用6%聚丙烯酰胺凝膠電泳分離并回收純化。然后采用高靈敏度DNA 片段生物分析儀對文庫進行質控,以保證PCR 切膠產物片段大小符合后續測序要求。最后采用高通量測序技術對該文庫進行測序。
1.8 miRNA分析及靶基因預測
將測序獲得的序列與miRBase v22 數據庫中大鼠已知的miRNA 進行對比,采用分析軟件Bowtie 1.1.1、miRDeep2、R 語言對已知miRNA 及預測miRNA 進行表達量標準化及相關性分析,采用R 包(EdgeR、Limma)對各組樣本進行miRNA 差異表達分析。然后利用Ensemble 數據庫、miRanda 靶基因預測軟件對差異miRNA 進行靶基因預測,并對靶基因進行基因本體論(gene ontology,GO)、Pathway 分析,以判定差異miRNA 主要影響的生物學功能和通路。
1.9 PCR法檢測差異miRNA的表達
根據小RNA 測序初篩結果,進一步選擇差異倍數較大的miRNA 進行PCR驗證,所有引物序列如表所示(表1)。取適量RNA 在逆轉錄體系中合成cDNA,以此為模板加入靶基因上下游引物,進行PCR 擴增。使用Sequence Detector System 軟件進行分析擴增反應所得的數據,以U6 為內參基因,釆用Ct法確定各miRNA的相對表達量。

表1 PCR實驗相關引物信息
1.10 統計學方法
應用HiSeq 2000 測序統計軟件對數據進行統計學分析,篩選樣本間差異表達的miRNA,采用顯著性P值和差異倍數(fold-change,FC)作為差異表達的判斷標準,當P<0.05,且FC≥Log2FC≥1,判斷為上調miRNA;當P<0.05,且FC≤Log0.5FC≤-1,判斷為下調miRNA。FC 值差異倍數越大,說明miRNA 在2 個分組樣本中的表達差異越大。PCR 驗證實驗中符合正態分布且方差齊的計量資料以均數±標準差(±s)表示,采用方差分析(ANOVA)進行多組變量間的相互比較,兩兩比較采用LSD-t檢驗。當P<0.05時,認為差異有統計學意義。
2 結果
2.1 樣本質控
視網膜總RNA 在260 nm/280 nm 處吸光度在1.9~2.1 A 之間;瓊脂糖凝膠電泳檢測28 S 和18 S 處條帶明亮、清晰、銳利,28 S 和18 S 處的RNA 比值≥1.5∶1;Qubit核酸分析試劑盒定量總RNA>2 μg。
2.2 3組miRNA片段長度分析
樣本測序得到的原始圖像經過轉化得到數據,去除測序質量較低、有5'接頭污染、沒有3'接頭序列、包括poly A 和小于18 nt 的小片段等,得到干凈序列統計小RNA 片段的長度分布圖(圖1)。3 組大部分的小RNA長度集中在21~24 nt,峰值出現在22 nt,符合miRNA的長度特征。
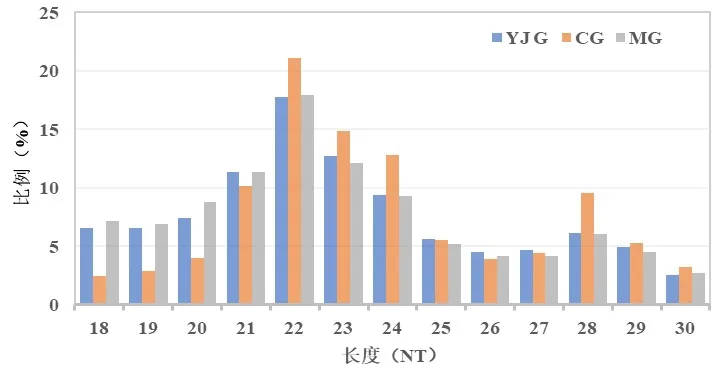
圖1 3組小RNA片段長度分布
2.3 3組已知miRNA差異表達分析
篩選出644 個已知miRNA,其中差異miRNA 共59 個。干預16 周后,與CG 組比較,MG 組35 個miRNA 上調、6 個miRNA 下調,YJG 組20 個miRNA上調、4 個miRNA 下調;與MG 組比較,YJG 組6 個miRNA 上調、13 個miRNA 下調;其中,1 個上調miRNA(miR-673-3p),11 個下調miRNA(miR-1-3p、miR-130a-5p、miR-133a-3p、miR-133b-3p、miR-1b、miR-206-3p、miR-214-3p、miR-23b-5p、miR-31a-3p、miR-451-5p、miR-466b-3p)為益景湯干預DM 大鼠后,發生相反變化趨勢的差異性miRNA,即3 組共同差異性表達miRNA(圖2、表2-表4),差異均有統計學意義(均P<0.05)。

表2 YJG組和CG組差異表達miRNA
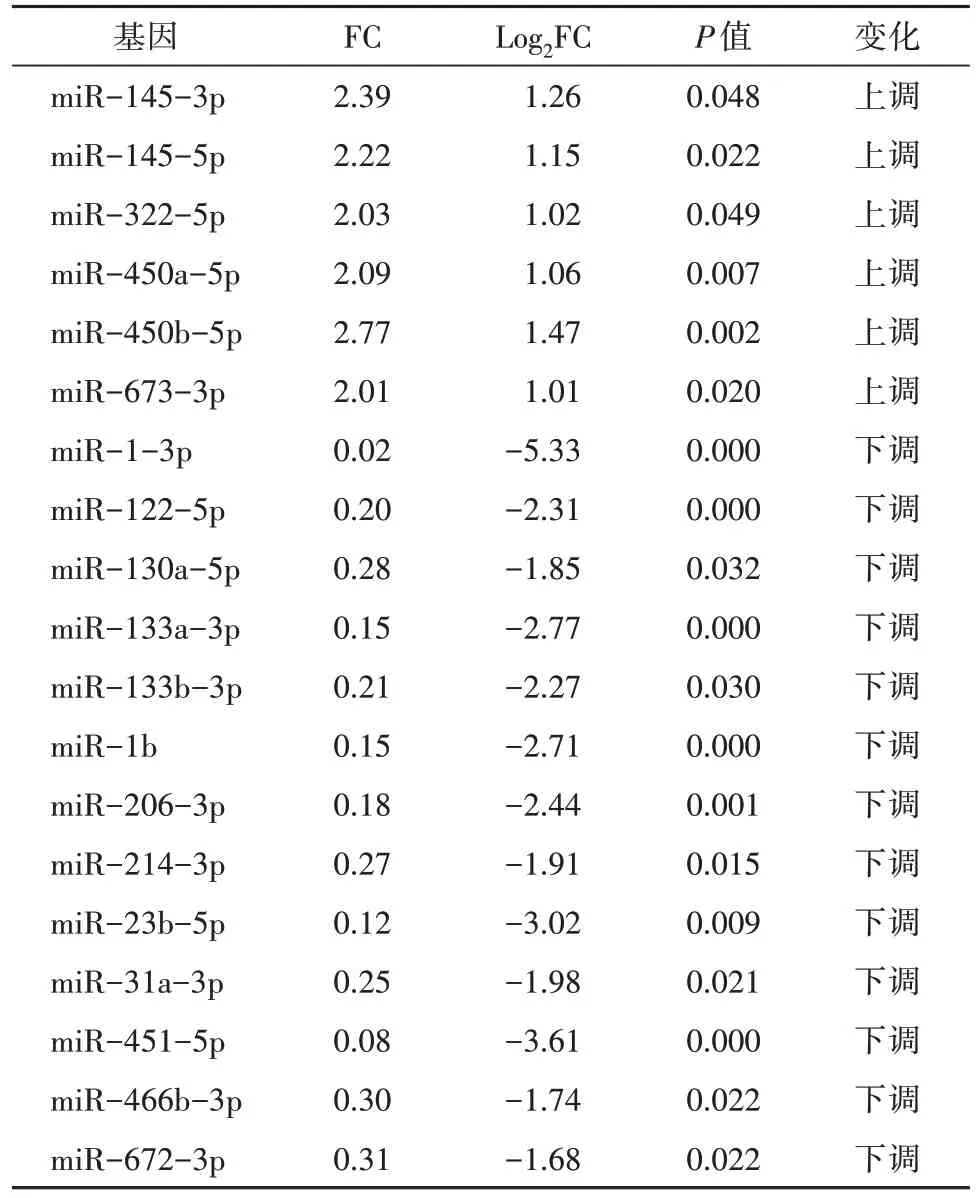
表3 YJG組和MG組差異表達miRNA
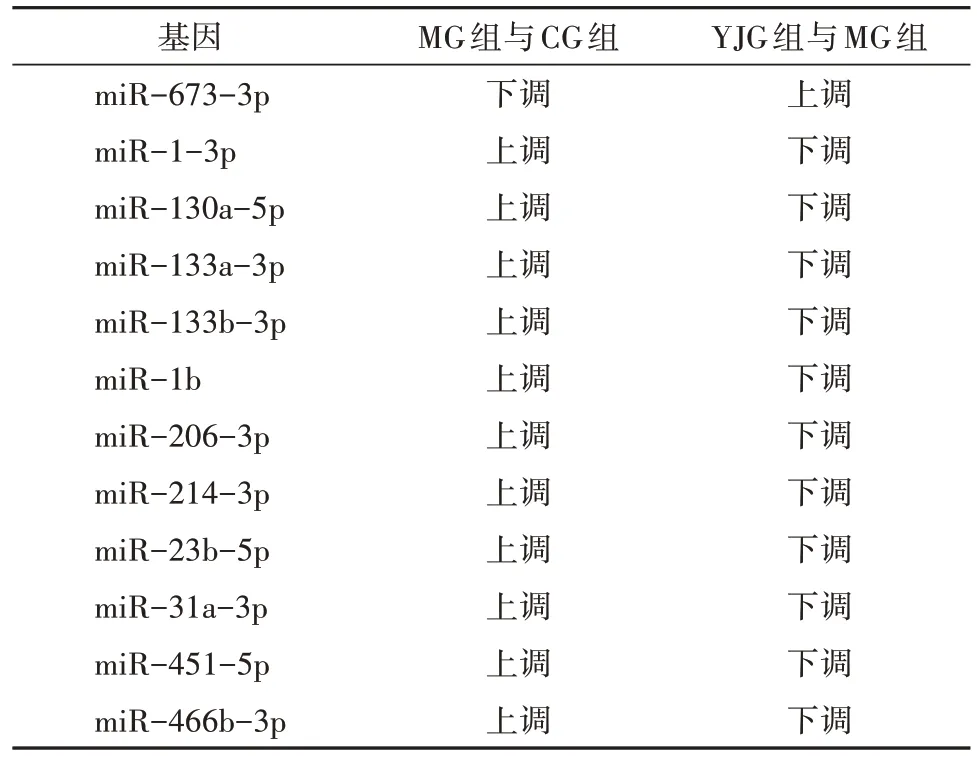
表4 3組共同差異性表達miRNA
2.4 3組新發現miRNA差異表達分析
3 組篩選出242 個新發現的miRNA,其中差異miRNA 共105 個(圖3)。與CG 組比較,MG 組50 個miRNA 上調,15 個miRNA 下調;與CG 組比較,YJG組55 個miRNA 上調,20 個miRNA 下調;與MG 組比較,YJG 組16個miRNA 上調,6個miRNA 下調,差異均有統計學意義(均P<0.05)。
圖3 3組新發現miRNA差異表達散點圖
2.5 差異表達miRNA的靶基因
采用miRanda 預測軟件比對Ensemble 數據庫,結合位點自由能閾值≥140,結合位點分值閾值≤-20 kcal/mol,取預測結果前10 名作為靶基因預測結果(表5)。

表5 益景湯干預DM大鼠前后差異表達miRNA相關靶基因
2.6 靶基因的GO分析結果
差異基因主要涉及生物學過程方面的多種蛋白的轉錄調節,其次是分子功能方面核苷酸三磷酸酶、GTP 酶等活性調節,還與多種細胞成分有關(圖4)。
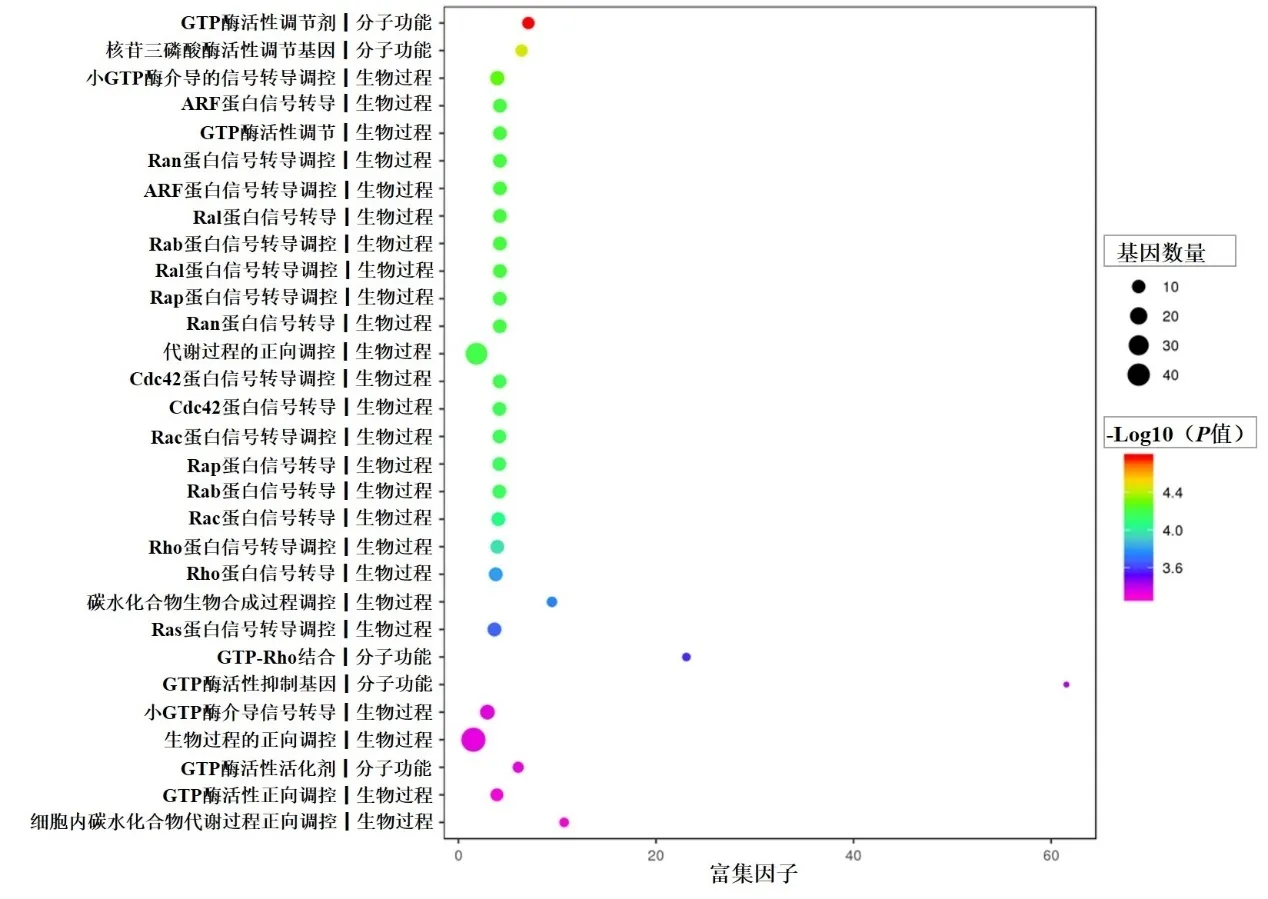
圖4 靶基因的GO分析
2.7 靶基因的Pathway分析結果
利用KEEG 數據庫對靶基因進行Pathway分析,發現靶基因關聯的通路主要是腫瘤相關的信號通路和糖脂代謝相關通路(圖5)。

圖5 靶基因的Pathway分析
2.8 PCR驗證
3 組間大鼠視網膜12 個差異性miRNA 的表達比較(表6、表7),差異均有統計學意義(FmiR-673-3p=22.938,P=0.002;FmiR-1-3p=42.288,P=0.000;FmiR-130a-5p=9.088,P=0.015;FmiR-133a-3p=15.912,P=0.004;FmiR-133b-3p=14.296,P=0.005;FmiR-1b=62.253,P=0.000;FmiR-206-3p=8.999,P=0.016;FmiR-214-3p=10.757,P=0.010;FmiR-23b-5p=112.387,P=0.000;FmiR-31a-3p=16.475,P=0.004;FmiR-451-5p=5.817,P=0.039;FmiR-466b-3p=122.070,P=0.000)。
表6 益景湯對DM大鼠視網膜部分miRNA表達的影響(±s,n=3)
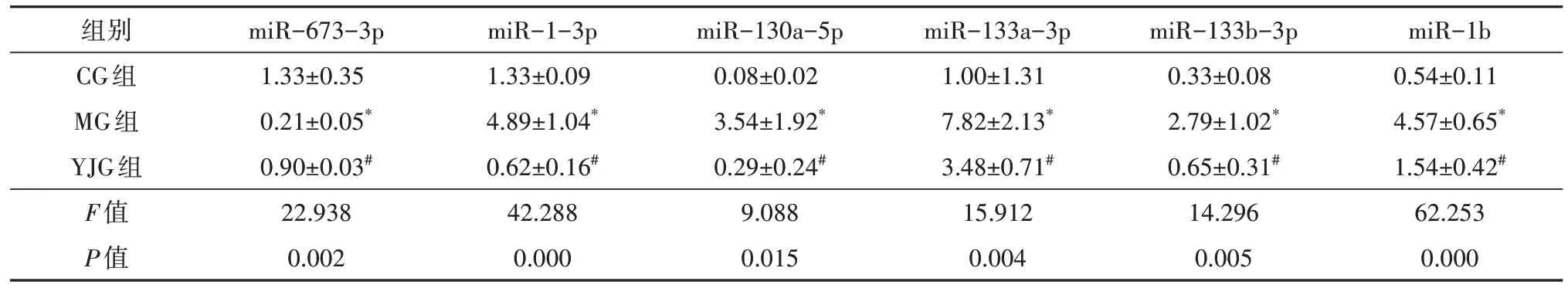
表6 益景湯對DM大鼠視網膜部分miRNA表達的影響(±s,n=3)
注:* 與CG組比較,P<0.05;# 與MG組比較,P<0.05;DM 糖尿病;CG 對照組;MG 模型組;YJG 益景湯組。
組別CG組MG組YJG組F值P值miR-673-3p 1.33±0.35 0.21±0.05*0.90±0.03#22.938 0.002 miR-1-3p 1.33±0.09 4.89±1.04*0.62±0.16#42.288 0.000 miR-130a-5p 0.08±0.02 3.54±1.92*0.29±0.24#9.088 0.015 miR-133a-3p 1.00±1.31 7.82±2.13*3.48±0.71#15.912 0.004 miR-133b-3p 0.33±0.08 2.79±1.02*0.65±0.31#14.296 0.005 miR-1b 0.54±0.11 4.57±0.65*1.54±0.42#62.253 0.000
表7 益景湯對DM大鼠視網膜部分miRNA表達的影響(±s,n=3)
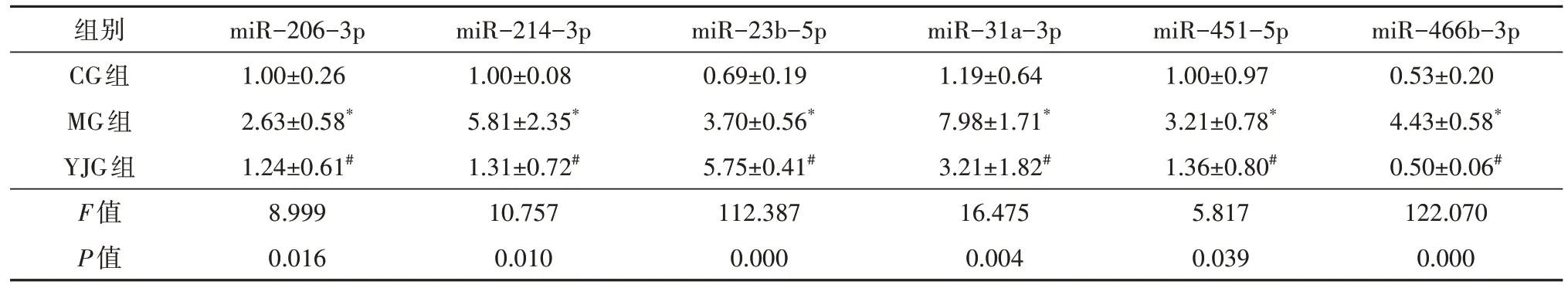
表7 益景湯對DM大鼠視網膜部分miRNA表達的影響(±s,n=3)
注:* 與CG組比較,P<0.05;# 與MG組比較,P<0.05;DM 糖尿病;CG 對照組;MG 模型組;YJG 益景湯組。
組別CG組MG組YJG組F值P值miR-206-3p 1.00±0.26 2.63±0.58*1.24±0.61#8.999 0.016 miR-214-3p 1.00±0.08 5.81±2.35*1.31±0.72#10.757 0.010 miR-23b-5p 0.69±0.19 3.70±0.56*5.75±0.41#112.387 0.000 miR-31a-3p 1.19±0.64 7.98±1.71*3.21±1.82#16.475 0.004 miR-451-5p 1.00±0.97 3.21±0.78*1.36±0.80#5.817 0.039 miR-466b-3p 0.53±0.20 4.43±0.58*0.50±0.06#122.070 0.000
兩兩比較,MG 組大鼠視網膜miR-673-3p 表達低于CG 組(t=6.708,P=0.001),余11 個miRNA 表達均高于CG 組(tmiR-1-3p=7.162,P=0.000;tmiR-130a-5p=3.804,P=0.009;tmiR-133a-3p=5.573,P=0.001;tmiR-133b-3p=4.910,P=0.003;tmiR-1b=10.818,P=0.000;tmiR-206-3p=3.929,P=0.008;tmiR-214-3p=4.143,P=0.006;tmiR-23b-5p=8.867,P=0.000;tmiR-31a-3p=5.590,P=0.001;tmiR-451-5p=3.175,P=0.019;tmiR-466b-3p=13.476,P=0.000),差異均有統計學意義。YJG 組大鼠視網膜miR-673-3p、miR-23b-5p 表達高于MG 組(tmiR-673-3p=4.164,P=0.006;tmiR-23b-5p=6.036,P=0.001),余10 個miRNA 表達均低于MG 組(tmiR-1-3p=8.607,P=0.000;tmiR-130a-5p=3.569,P=0.012;tmiR-133a-3p=3.545,P=0.012;tmiR-133b-3p=4.288,P=0.005;tmiR-1b=7.285,P=0.001;tmiR-206-3p=3.350,P=0.015;tmiR-214-3p=3.878,P=0.008;tmiR-31a-3p=3.924,P=0.008;tmiR-451-5p=2.667,P=0.037;tmiR-466b-3p=13.586,P=0.000),差異均有統計學意義。除miR-23b-5p 外,其余11 個miRNA 與上述RNA 檢測結果一致。
3 討論
miRNA 在蛋白質調節上具有重要作用,通過與靶基因的mRNA 3'非編碼區的相互作用,抑制mRNA 翻譯或誘導其降解,從轉錄水平調節靶基因的表達,從而參與細胞的發育、增殖、分化、死亡以及糖脂代謝等重要進程[13]。研究[14-17]表明,miRNA通過各種不同的機制,參與DR 的發生、發展,在DR的早期診斷和后續治療中具有潛在價值。
DR早期病理改變早在大鼠DM造模后3個月即可出現[18-20],此階段視網膜組織中被發現80 種miRNA 表達顯著升高,6種表達顯著下降;視網膜內皮細胞中有11 種miRNA 表達升高,104 種表達降低[21]。其中miR-21、miR-146、miR-155 和miR-132被揭示與DM 大鼠視網膜內皮細胞中核因子κB(nuclear factor κB,NF-κB)炎癥通路相關;miR-17-5p、miR-18a、miR-20a,miR-21、miR-31 和miR-155則被發現和VEGF相關。本研究發現,DM 大鼠成模16 周后,視網膜組織中35 種miRNA 上調,6 種miRNA 下調,其中miR-31、miR-199a、miR-200a、miR-200b、miR-205、miR-223 上調以及miR-20b 下調趨勢與上述研究一致。
在增殖性DR 中,miR-200a、miR-200b 表達降低,與細胞的增殖、凋亡和調控VEGF 表達水平相關[17,20-21]。本研究發現,在早期DR 大鼠視網膜組織中miR-200a、miR-200b 表達水平升高,提示DR 早期和中晚期的miRNA 表達情況有所不同,可能與視網膜缺血、缺氧程度及病理改變程度相關。
miR-1 和miR-133 在DM 患者血清水平明顯升高,胰島素樣生長因子1(insulin-like growth factor 1,IGF-1)是miR-1 和miR-133 的有效靶點之一,2 種miRNA 負調控其表達,而IGF-1 水平與胰島素抵抗及糖尿病進展呈負相關[22]。miR-133b 可下調Rho相關蛋白激酶信號通路來抑制視網膜色素上皮細胞增殖,并促進其凋亡[23];miR-23b 在高糖培養的人視網膜血管內皮細胞模型中顯著升高,抑制沉默信息調節因子1 轉錄,使NF-κB 活化增加,促進細胞凋亡,干擾miR-23b 表達后,NF-κB 乙酰化顯著降低,細胞凋亡明顯減少[24]。本研究亦發現,miR-133b、miR-23b 在DM 大鼠視網膜中明顯升高,益景湯干預后其表達降低,提示miR-133b、miR-23b 可能是益景湯保護視網膜血管內皮細胞,干預早期DR 的治療靶點。前期細胞實驗[6]發現,益景湯能減少高糖環境中大鼠視網膜微血管周細胞凋亡、人腦微血管內皮細胞的凋亡[25]對此亦有所提示。
miR-214、miR-451 在缺血狀態下的視網膜細胞中顯著升高,miR-31 水平下降,眼內注射miR-31能夠減少視網膜新生血管形成[26-27]。但KAUR P等[28]的研究表明,miR-31 在DM 患者視網膜細胞中表達水平上調,通過介導VEGF在DR中發揮不良作用;同時miR-31 水平與白細胞滾動速度呈正相關,與促炎因子和黏附分子誘導的白細胞向血管壁聚集有關[29]。CAO Q 等[30]發現,miR-673 靶向抑制非受體型酪氨酸蛋白激酶2(Janus kinase 2,JAK2)表達調節輔助性T 細胞(T helper cell 17,Th17)分化,誘導胎盤生長因子分泌,促進新生血管形成,上調miR-673可抑制新生血管形成。本研究發現,DM大鼠視網膜組織中miR-31a 表達升高,miR-673 表達降低,益景湯干預后miR-31a表達降低,miR-673表達升高,和上述研究結果一致。團隊前期臨床研究和動物研究[3-4]都發現益景湯能改善DR 患者視網膜微循環,調控VEGF 的水平;緩解DR 大鼠視網膜毛細血管床的減少,改善視網膜血供,減輕視網膜滲漏[7,31-32]。因此,通過調節miR-31a、miR-673 表達,改善視網膜微循環,抑制新生血管形成,可能是益景湯干預早期DR微血管損害的分子機制之一。
綜上所述,益景湯干預大鼠早期DR 微血管損害,miR-673、miR-1、miR-133、miR-214、miR-23b、miR-31a、miR-451 等可能是其作用靶點,其機制與緩解視網膜組織缺血,調節視網膜微血管內皮細胞異常增殖和凋亡,抑制視網膜新生血管形成以及抗炎等過程有關,但更深入機制還有待進一步研究。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55
