NRG1基因修飾的骨髓基質(zhì)干細(xì)胞移植對大鼠脊髓半橫切損傷的修復(fù)作用及其機(jī)制
2024-03-25 09:35:20符禹玄陳君趙富生李媛媛張可心武庚
解放軍醫(yī)學(xué)雜志 2024年2期
關(guān)鍵詞:模型
符禹玄,陳君,趙富生,李媛媛,張可心,武庚*
1牡丹江醫(yī)學(xué)院第一臨床醫(yī)學(xué)院骨科,黑龍江牡丹江 157000;2牡丹江醫(yī)學(xué)院組織學(xué)與胚胎學(xué)教研室,黑龍江牡丹江157000
脊髓損傷(spinal cord injury,SCI)是一種由交通事故、高處墜落等引起的嚴(yán)重中樞神經(jīng)系統(tǒng)疾病,主要表現(xiàn)為脊髓受損平面以下感覺、運(yùn)動和自主神經(jīng)功能障礙或喪失,可嚴(yán)重影響生活質(zhì)量,同時給患者家庭和社會帶來沉重的經(jīng)濟(jì)負(fù)擔(dān)[1]。SCI可分為原發(fā)性和繼發(fā)性損傷兩個連續(xù)的病理階段[2]。原發(fā)性SCI 多因脊柱骨折壓迫脊髓引起,繼發(fā)性損傷是由水腫、炎癥反應(yīng)、氧化應(yīng)激、鈣超載、神經(jīng)元凋亡等繼發(fā)性損害因素引起的一系列病理變化[3-4]。因此有效抑制神經(jīng)元壞死和凋亡,恢復(fù)神經(jīng)傳導(dǎo)功能,成為臨床治療SCI 的重點(diǎn)和難題[5]。神經(jīng)調(diào)節(jié)蛋白1(neureglin1,NRG1)是一種含表皮生長因子樣結(jié)構(gòu)域的跨膜蛋白,屬于神經(jīng)生長因子家族,參與突觸塑形、髓鞘形成、神經(jīng)細(xì)胞發(fā)育及成熟等多種生理過程[6]。研究發(fā)現(xiàn),NRG1 與其下游信號受體ErbBs 結(jié)合,形成同源二聚體或異二聚體,通過激活胞內(nèi)酪氨酸激酶,參與調(diào)控神經(jīng)元和膠質(zhì)細(xì)胞的增殖和分化過程[6]。NRG1還與阿爾茨海默病、帕金森病等多種神經(jīng)退行性疾病的發(fā)生發(fā)展相關(guān)[7]。骨髓基質(zhì)干細(xì)胞(bone marrow mesenchymal stem cells,BMSCs)是骨髓中的非造血干細(xì)胞,具有高度自我復(fù)制能力和多向分化潛能[8-10]。BMSCs可分泌多種神經(jīng)細(xì)胞營養(yǎng)因子,如血管內(nèi)皮生長因子、腦源性神經(jīng)營養(yǎng)因子等,改善損傷局部微環(huán)境,促進(jìn)損傷細(xì)胞修復(fù)[8]。BMSCs 易于進(jìn)行基因修飾,并能夠穩(wěn)定表達(dá)外源基因[11],且取材方便,易于擴(kuò)增,可降低免疫排斥反應(yīng),因而成為較理想的組織工程種子細(xì)胞[12]。有研究發(fā)現(xiàn),在大鼠大腦中動脈閉塞模型中,BMSCs 移植可減輕卒中后腦組織損傷[13]。本研究通過建立大鼠SCI模型,移植并觀察NRG1-BMSCs對SCI的修復(fù)作用,以期為臨床治療SCI提供新思路。
1 材料與方法
1.1 主要試劑及儀器 pcDNA3.1(+)-Nrg1-IRES/eGFP 質(zhì)粒由廣州賽業(yè)生物科技有限公司構(gòu)建及鑒定;DMEM 培養(yǎng)液、胎牛血清、ECL 化學(xué)發(fā)光試劑盒(上海碧云天生物技術(shù)有限公司);青鏈霉素雙抗溶液(SV30010,美國Hyclone 公司);FuGene6(E269,美國Promega 公司);NRG1 ELISA 試劑盒(上海康朗生物科技有限公司);兔抗鼠葡萄糖調(diào)節(jié)蛋白78(glucose regulated protein 78,GRP78)、轉(zhuǎn)錄激活因子4(activating transcription factor 4,ATF4)、ATF6、C/EBP同源蛋白(C/EBP homologous protein,CHOP)、X-box結(jié)合蛋白1(X-box binding protein 1,XBP1)、B 淋巴細(xì)胞瘤-2 蛋白(B-cell lymphoma-2,Bcl-2)、Bcl-2 相關(guān)X蛋白(Bcl-2-asslciated protein X,Bax)抗體(武漢三鷹生物技術(shù)有限公司)。PVDF膜(上海碧云天生物技術(shù)有限公司);CO2細(xì)胞培養(yǎng)箱(MCO-15AC,日本SANYO公 司);微 量 注 射 器(UMC4 Micro4TMController)、Model ELX800 酶標(biāo)儀(ELX800,美國Bio-Tek 公司);冷凍切片機(jī)(CM3050S,德國Leica 公司);體視顯微鏡(SMZ745T,日本Nikon公司);激光共聚焦顯微鏡(LSM700,德國Zeiss 公司);凝膠成像系統(tǒng)(Gel Doc XR+,美國Bio-Rad 公司);Image-Pro PLUS 圖像分析軟件(美國Meyer Instruments公司)。
1.2 方法
1.2.1 實驗動物 4 周齡SPF 級SD 大鼠4 只,體重(100±5) g,用于BMSCs 分離培養(yǎng);8 周齡SPF 級SD大鼠43 只,雌22 只,雄21 只,體重(280±20) g,由牡丹江醫(yī)學(xué)院實驗動物中心提供[實驗動物使用許可證號:SYXK(黑)2019-006]。實驗前適應(yīng)性飼養(yǎng)1周,環(huán)境溫度20~24 ℃,濕度42%~46%,晝夜12 h 交替光照,自由攝食飲水。實驗過程符合國家及單位實驗動物的管理和使用規(guī)定。
1.2.2 BMSCs 分離培養(yǎng) 將4周齡SD 大鼠頸椎脫臼處死,置70%乙醇中浸泡10 min,無菌條件下分離股骨和脛骨,剪除骨骺,暴露骨髓腔,用20 ml DMEM 培養(yǎng)液沖洗骨髓腔,經(jīng)200 目無菌篩網(wǎng)過濾后,將細(xì)胞接種于50 ml 培養(yǎng)瓶中,置于37 ℃、5%CO2培養(yǎng)箱中培養(yǎng),72 h后更換培養(yǎng)液,去除未貼壁細(xì)胞,之后每隔3 d更換1次培養(yǎng)液,于倒置相差顯微鏡下觀察細(xì)胞形態(tài)變化,并采集圖像。當(dāng)細(xì)胞融合度達(dá)80%~90%時,進(jìn)行傳代培養(yǎng),取第3 代BMSCs進(jìn)行后續(xù)實驗。
1.2.3 NRG1 質(zhì) 粒 轉(zhuǎn) 染BMSCs 將BMSCs 按1×105個/ml接種于6孔板,當(dāng)細(xì)胞融合度達(dá)80%時,按照FuGene6 說明書將NRG1 質(zhì)粒或空載質(zhì)粒轉(zhuǎn)染入BMSCs,具 體 操 作 如 下:取91 μl DMEM、6 μl FuGene6、3 μl NRG1 質(zhì)粒混勻后加入BMSCs 培養(yǎng)液中,培養(yǎng)48 h 后更換含G418 的培養(yǎng)液,收集G418抗性NRG1-BMSCs 擴(kuò)增培養(yǎng),在熒光顯微鏡下觀察BMSCs 轉(zhuǎn)染情況,并采集圖像。按照ELISA 試劑盒說明書步驟分別檢測BMSCs 和NRG1-BMSCs 裂解液及培養(yǎng)液中NRG1蛋白含量。
1.2.4 細(xì)胞增殖檢測 取生長狀態(tài)良好的BMSCs 和NRG1-BMSCs,制備細(xì)胞懸液,調(diào)整細(xì)胞密度為1×105個/ml,以每孔相同的細(xì)胞數(shù)接種于24 孔板,連續(xù)培養(yǎng)14 d,每天取3孔細(xì)胞消化,應(yīng)用細(xì)胞計數(shù)板計數(shù),取平均值繪制細(xì)胞生長曲線。
1.2.5 動物實驗分組及脊髓半橫切損傷模型制備取8周齡SPF級SD大鼠隨機(jī)分為對照組(n=10)、SCI模型組(n=10)、空載體-BMSCs(BMSCs)組(n=10)和NRG1-BMSCs 組(n=13,其中3 只用于細(xì)胞遷移檢測)。SCI 模型組、BMSCs 組和NRG1-BMSCs 組大鼠參照趙富生等[14]的方法,建立脊髓半橫切模型。具體操作如下:用1.5%戊巴比妥鈉(30 mg/kg)腹腔注射麻醉大鼠,俯臥位固定于手術(shù)臺上,背部手術(shù)區(qū)去毛、消毒,以T8棘突為中心,行背部正中縱行切口,長約3 cm,用眼科彎鑷鈍性分離筋膜及雙側(cè)椎旁肌,暴露T7-T9三個棘突、雙側(cè)椎板及橫突,用止血鉗固定T8并輕輕向上提起,張開椎間隙,剪除T8棘突及棘突下椎板暴露脊髓,在體式顯微鏡下,用眼科虹膜刀緊貼脊髓正中動脈右側(cè)垂直刺入脊髓至椎管底部,刀鋒向右劃動至右側(cè)脊髓完全橫斷,大鼠即刻出現(xiàn)一過性鼠尾連續(xù)性抽動和后肢持續(xù)性痙攣,表示造模成功。術(shù)后使用無菌明膠海綿填塞術(shù)區(qū),逐層縫合硬脊膜、肌肉、筋膜和皮膚,注射青霉素(20 萬U/kg),2 次/d,連續(xù)3 d,將飼料、飲水置于大鼠可及范圍,保持墊料干燥。
造模成功后2 h,BMSCs 組和NRG1-BMSCs 組大鼠在脊髓損傷中心(垂直進(jìn)針)以及損傷中心上、下0.5 cm 處傾斜45°進(jìn)針,刺入深度為1.5 mm,使用微量注射器以5 μl/min 勻速注入5 μl BMSCs 或NRG1-BMSCs 懸液(1×107個/ml)至皮質(zhì)脊髓側(cè)束(圖1),注射后留針3 min;SCI 模型組以同樣方法注入15 μl PBS。對照組僅行T8棘突及椎板切除術(shù)。

圖1 大鼠脊髓半橫切損傷模型及細(xì)胞移植示意圖Fig.1 Schematic diagram of rat spinal cord hemisection injury model and cell transplantation
1.2.6 大鼠術(shù)后運(yùn)動功能和脊髓組織病理學(xué)及相關(guān)蛋白表達(dá)水平觀察
1.2.6.1 NRG1-BMSCs 遷移及分布觀察 細(xì)胞移植第7天,隨機(jī)選取NRG1-BMSCs組3只大鼠,用1.5%戊巴比妥鈉過量麻醉后,左心室插管灌注4%多聚甲醛溶液,切取損傷節(jié)段脊髓用OCT包埋,冰凍切片機(jī)常規(guī)切片(厚度6~10 μm)。采用DAPI 染細(xì)胞核,用抗熒光淬滅劑封片后,于熒光顯微鏡下觀察NRG1-BMSCs遷移及分布情況。
1.2.6.2 大鼠后肢運(yùn)動功能評估 造模后第1、7、14、21、28 天,各組隨機(jī)選取6 只大鼠進(jìn)行BBB 評分和斜板實驗。BBB 評分范圍為0~21 分,0 分表示癱瘓,21 分表示后肢運(yùn)動功能正常。斜板實驗將大鼠置于平坦、不光滑的斜板上,斜板由0°緩慢上升,每次升高5°,大鼠堅持時間至少5 s不從斜板上跌落為成功,使用量角器測量斜板角數(shù)。實驗采用雙盲、雙人獨(dú)立完成評價,最終結(jié)果取平均值。
1.2.6.3 HE 染色、Nissl 染色和TUNEL 染色 完成BBB 評分和斜板實驗后,各組隨機(jī)選取5 只大鼠經(jīng)1.5%戊巴比妥鈉過量麻醉后,剖胸行左心室插管,灌注4%多聚甲醛溶液固定,切取0.5 cm損傷節(jié)段脊髓,以損傷為中心橫斷為兩段,一段制備石蠟切片行HE 染色和Nissl 染色,另一段制備冰凍切片行TUNEL染色。
HE染色:石蠟切片經(jīng)二甲苯脫蠟和梯度乙醇水化后,蘇木精染液染色10 min,1%酸性乙醇浸泡5 s,蒸餾水漂洗,置于伊紅溶液中染色5 min,經(jīng)梯度乙醇脫水、中性香脂封片后,于光學(xué)顯微鏡下觀察組織形態(tài)結(jié)構(gòu),并采集圖像。
Nissl 染色:切片經(jīng)脫蠟水化后,置于Nissl 染液中染色,于光學(xué)顯微鏡下觀察脊髓組織神經(jīng)元形態(tài)結(jié)構(gòu)以及尼氏體密度和分布情況。
TUNEL 染色:取冰凍切片,按照TUNEL 試劑盒說明書進(jìn)行染色,在激光共聚焦顯微鏡下觀察,TUNEL 陽性細(xì)胞呈現(xiàn)紅色熒光,采用ImageJ 軟件分析TUNEL 陽性細(xì)胞率。TUNEL 陽性細(xì)胞率(%)=(TUNEL陽性細(xì)胞數(shù)/DAPI染色細(xì)胞數(shù))×100%。
1.2.6.4 Western blotting 檢測脊髓組織中內(nèi)質(zhì)網(wǎng)應(yīng)激相關(guān)蛋白及凋亡相關(guān)蛋白的表達(dá) 完成BBB 評分和斜板實驗后,取各組剩余5 只大鼠的脊髓組織,經(jīng)RIPA 裂解、離心,取上清采用BCA 法測定蛋白含量。通過SDS-PAGE 凝膠電泳分離蛋白,經(jīng)轉(zhuǎn)膜、封閉后,加入XBP1(1∶1000)、CHOP(1∶800)、ATF4(1∶1000)、ATF6(1∶1000)、GRP78(1∶800)、Bcl-2(1∶500)、Bax(1∶400)和β-actin(1∶2000)一抗,4 ℃孵育過夜,次日TBST清洗3次,加入相應(yīng)二抗37 ℃孵育2 h,滴加ECL發(fā)光試劑,置于成像儀中成像,應(yīng)用ImageJ軟件分析目的蛋白條帶相對灰度值。
1.3 統(tǒng)計學(xué)處理 應(yīng)用SPSS 18.0 軟件進(jìn)行統(tǒng)計分析。所有數(shù)據(jù)以±s表示,兩組間比較采用獨(dú)立樣本t 檢驗,多組間比較采用單因素方差分析,方差齊時,進(jìn)一步兩兩比較采用Bonferroni 校正t 檢驗;方差不齊時,進(jìn)一步兩兩比較采用Tamhane's T2檢驗。P<0.05為差異有統(tǒng)計學(xué)意義。
2 結(jié) 果
2.1 BMSCs 培養(yǎng)及NRG1 基因修飾 倒置相差顯微鏡觀察顯示,培養(yǎng)第1 天,BMSCs 呈圓形、不規(guī)則形或多角形,分布均勻(圖2A);培養(yǎng)第3 天,BMSCs 形成多個散在集落,緊密排列呈聚集生長(圖2A);傳代培養(yǎng)后,BMSCs 分布均勻呈長梭形或束狀生長(圖2A)。熒光顯微鏡觀察顯示,轉(zhuǎn)染NRG1基因后,BMSCs呈明亮綠色熒光(圖2B)。
ELISA 法檢測結(jié)果顯示,BMSCs 培養(yǎng)液和裂解液中NRG1 含量在轉(zhuǎn)染24 h 略有升高,但差異無統(tǒng)計 學(xué) 意 義(P>0.05),轉(zhuǎn) 染48 h 明 顯 升 高(P<0.05,圖2C、D)。
細(xì)胞增殖檢測結(jié)果顯示,BMSCs 和NRG1-BMSCs生長曲線呈S形(圖2E)。接種后第1~3天為滯留期,第6~8天進(jìn)入對數(shù)生長期,第12天達(dá)到高峰,此后細(xì)胞生長緩慢,進(jìn)入平臺期,且NRG1-BMSCs增殖速度明顯快于BMSCs(P<0.05,圖2E)。
2.2 NRG1-BMSCs 遷移和分布情況 移植第7 天,NRG1-BMSCs 大鼠脊髓組織切片中可觀察到呈現(xiàn)綠色熒光的移植細(xì)胞,細(xì)胞呈圓形、橢圓形或不規(guī)則形狀,多聚集在損傷中心,偶見少量NRG1-BMSCs向損傷中心周圍區(qū)域遷移擴(kuò)散(圖3)。

圖3 GFP標(biāo)記的NRG1-BMSCs在脊髓組織內(nèi)示蹤觀察(A-C,×50;D,×100)Fig.3 Trace observation of GFP-labeled NRG1-BMSCs in spinal cord tissues
2.3 各組大鼠后肢運(yùn)動功能比較 術(shù)前各組大鼠BBB 評分均為21 分。術(shù)后第1、7、14、21、28 天,SCI模型組、BMSCs組和NRG1-BMSCs組BBB評分均明顯低于對照組(P<0.05)。術(shù)后第7、14 天,SCI 模型組、BMSCs 組和NRG1-BMSCs 組BBB 評分均較術(shù)后第1 天增高,但組間比較無統(tǒng)計學(xué)差異(P>0.05);術(shù)后第21、28 天,NRG1-BMSCs 組BBB 評分明顯高于SCI 模型組和BMSCs 組(P<0.05),BMSCs 組與SCI模型組BBB評分無統(tǒng)計學(xué)差異(P>0.05,圖4A)。

圖4 移植后不同時間點(diǎn)各組大鼠BBB評分和斜板傾斜角度比較Fig.4 Comparison of the BBB scores and the slope angle of rats in each group at different time points after transplantation
術(shù)前各組大鼠斜板實驗均為60°,術(shù)后第1、7、14、21、28 天,SCI 模 型 組、BMSCs 組 和NRG1-BMSCs 組斜板傾斜角度均明顯低于對照組(P<0.05)。術(shù)后第7、14 天,SCI 模型組、BMSCs 組和NRG1-BMSCs 組斜板傾斜角度均較術(shù)后第1 天增高,但組間差異無統(tǒng)計學(xué)意義(P>0.05);術(shù)后第21、28 天,NRG1-BMSCs組斜板傾斜角度明顯高于SCI模型組和BMSCs組(P<0.05),BMSCs組與SCI模型組斜板傾斜角度無統(tǒng)計學(xué)差異(P>0.05)(圖4B)。
2.4 各組大鼠脊髓組織病理學(xué)改變 HE 染色結(jié)果顯示,移植第28天,對照組脊髓組織結(jié)構(gòu)清晰,神經(jīng)元形態(tài)結(jié)構(gòu)正常,細(xì)胞核大而圓,核仁明顯;SCI模型組脊髓灰質(zhì)中有炎性細(xì)胞浸潤,空洞較多,可見神經(jīng)元核固縮、凋亡;BMSCs 組脊髓組織結(jié)構(gòu)紊亂,神經(jīng)元形態(tài)不規(guī)則,空洞明顯;NRG1-BMSCs組脊髓組織中炎性細(xì)胞浸潤明顯減少,神經(jīng)元核固縮、壞死減輕,空洞減少(圖5)。

圖5 各組大鼠脊髓組織HE染色(×200)和Nissl染色(×200)Fig.5 HE staining and Nissl staining of spinal cord tissues of rats in each group
Nissl 染色結(jié)果顯示,對照組神經(jīng)元胞質(zhì)中尼氏體豐富、染色清晰;SCI 模型組神經(jīng)元溶解、破壞,尼氏體密度降低;BMSCs組神經(jīng)元排列紊亂、皺縮,尼氏體減少;NRG1-BMSCs組神經(jīng)元形態(tài)結(jié)構(gòu)完整,尼氏體密度明顯增加(圖5)。
2.5 各組大鼠脊髓組織中內(nèi)質(zhì)網(wǎng)應(yīng)激相關(guān)蛋白表達(dá)水平比較 Western blotting檢測結(jié)果顯示,與對照組相比,SCI 后大鼠脊髓組織中內(nèi)質(zhì)網(wǎng)應(yīng)激相關(guān)蛋白XBP1、CHOP、ATF4、ATF6 和GRP78 表達(dá)水平明顯升高(P<0.05);經(jīng)NRG1-BMSCs移植治療后,內(nèi)質(zhì)網(wǎng)應(yīng)激相關(guān)蛋白表達(dá)水平明顯降低(P<0.05),但未恢復(fù)到對照組水平(P<0.05);SCI模型組與BMSCs組脊髓組織中XBP1、CHOP、ATF4、ATF6 和GRP78 蛋白表達(dá)水平差異無統(tǒng)計學(xué)意義(P>0.05)(圖6)。

圖6 各組大鼠脊髓組織內(nèi)質(zhì)網(wǎng)應(yīng)激相關(guān)蛋白XBP1、CHOP、ATF4、ATF6和GRP78表達(dá)水平比較Fig.6 Comparison of expression levels of endoplasmic reticulum stress-related proteins XBP1, CHOP, ATF4, ATF6 and GRP78 in rat spinal cord tissues
2.6 各組大鼠脊髓組織TUNEL 染色陽性細(xì)胞率比較 細(xì)胞移植第28 天,SCI 模型組脊髓損傷區(qū)域可見大量TUNEL 陽性細(xì)胞(細(xì)胞核呈紅色),而NRG1-BMSCs 組TUNEL 陽性細(xì)胞明顯減少(圖7)。NRG1-BMSCs 組TUNEL 陽性細(xì)胞率明顯低于SCI 模型組(P<0.05),但 高 于 對 照 組(P<0.05);SCI 模 型 組 與BMSCs 組TUNEL 陽性細(xì)胞率差異無統(tǒng)計學(xué)意義(P>0.05)(圖7)。

圖7 各組大鼠脊髓組織TUNEL陽性細(xì)胞率比較(TUNEL ×200)Fig.7 Comparison of TUNEL-positive cells rate in spinal cord tissues of rats in each group (TUNEL ×200)
2.7 各組大鼠脊髓組織中凋亡相關(guān)蛋白表達(dá)水平比較 Western blotting 檢測結(jié)果顯示,細(xì)胞移植第28天,與對照組相比,SCI 后大鼠脊髓組織中Bcl-2 蛋白表達(dá)水平明顯降低(P<0.05),Bax 蛋白表達(dá)水平明顯升高(P<0.05);NRG1-BMSCs組損傷節(jié)段脊髓組織中Bcl-2 蛋白表達(dá)水平明顯高于BMSCs 組和SCI 模型組(P<0.05),而Bax 蛋白表達(dá)水平明顯低于BMSCs 組和SCI 模型組(P<0.05);BMSCs 組與SCI 模型組Bcl-2、Bax 蛋白表達(dá)水平比較差異無統(tǒng)計學(xué)意義(P>0.05)(圖8)。
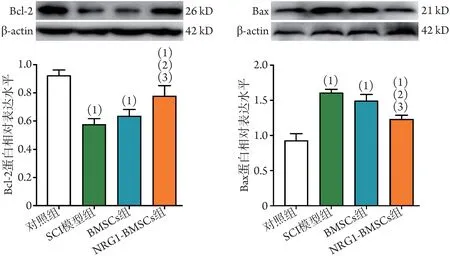
圖8 各組大鼠脊髓組織中凋亡相關(guān)蛋白Bcl-2、Bax表達(dá)水平比較Fig.8 Comparison of expression levels of apoptosis-related proteins Bcl-2 and Bax in spinal cord tissues of rats in each group
3 討 論
SCI通常是由脊柱創(chuàng)傷引起的破壞性中樞神經(jīng)系統(tǒng)疾病,目前其治療方法主要包括藥物治療、手術(shù)減壓、康復(fù)理療等手段,但均未取得滿意效果。本研究采用脊髓半橫切損傷模型,探討移植NRG1 基因修飾的BMSCs 對SCI 的保護(hù)作用,結(jié)果顯示,NRG1能夠提高BMSCs的治療效果,促進(jìn)大鼠SCI后運(yùn)動功能恢復(fù),其機(jī)制可能與增加BMSCs 增殖活性、抑制脊髓組織細(xì)胞凋亡、減輕內(nèi)質(zhì)網(wǎng)應(yīng)激有關(guān)[15]。
BMSCs 具有在特定條件下分化為神經(jīng)細(xì)胞的特性,并可自體移植、促進(jìn)軸突再生,因而在中樞神經(jīng)系統(tǒng)疾病治療中應(yīng)用前景廣闊[16]。研究發(fā)現(xiàn),BMSCs 中MHC-I 類分子表達(dá)水平較低,且缺乏MHC-Ⅱ類分子和共刺激分子表達(dá),這些特性致使BMSCs 移植后極少引起同種異體免疫排斥反應(yīng)[17]。此外,BMSCs 通過分泌白細(xì)胞抗原G5、T 淋巴細(xì)胞發(fā)育抑制因子、白血病抑制因子以及γ 干擾素,可使其免受T淋巴細(xì)胞和NK細(xì)胞的損害[17]。有研究發(fā)現(xiàn),將BMSCs 移植于SCI 模型大鼠中,大鼠損傷部位M2 型巨噬細(xì)胞數(shù)量明顯增多,M1 型巨噬細(xì)胞數(shù)量明顯減少,且脊髓組織中TNF-α、IL-6 含量也明顯減少,提示BMSCs 可減輕SCI 炎癥反應(yīng)。同時,BMSCs 能夠減輕SCI 后神經(jīng)性疼痛,阻止血脊髓屏障破壞,抑制星形膠質(zhì)細(xì)胞增生引起的瘢痕形成[18]。但研究發(fā)現(xiàn),單純移植BMSCs 治療效果有限,聯(lián)合其他治療方式或進(jìn)行基因修飾能夠顯著提高BMSCs的治療效果[17]。有研究發(fā)現(xiàn),將KLF7 基因?qū)隑MSCs,可明顯上調(diào)BMSCs 中KLF7 的表達(dá),提高BMSCs的增殖活性,抑制BMSCs凋亡,增高BMSCs中神經(jīng)生長因子分泌水平[19]。本研究成功分離培養(yǎng)BMSCs,并轉(zhuǎn)染NRG1基因。在NRG1修飾的BMSCs培養(yǎng)液和細(xì)胞裂解液中,NRG1 表達(dá)水平呈時間依賴性增高,并能夠顯著增加BMSCs 的增殖活性,提示將NRG1-BMSCs 移植到脊髓組織可能增加損傷組織中NRG1 含量,一定程度改善局部微環(huán)境,促進(jìn)神經(jīng)修復(fù)。同時,NRG1 也可能促進(jìn)BMSCs 存活以及向損傷部位遷移,改善神經(jīng)功能。有研究發(fā)現(xiàn),應(yīng)用NRG1 基因轉(zhuǎn)染人脂肪源性基質(zhì)干細(xì)胞,能夠上調(diào)該細(xì)胞中NRG1的表達(dá);將NRG1高表達(dá)的脂肪基質(zhì)干細(xì)胞注射到大鼠紋狀體14 d 后,可增高大鼠腦組織中NRG1 蛋白表達(dá)水平,促進(jìn)卒中大鼠肢體運(yùn)動功能恢復(fù)[20]。NRG1 是神經(jīng)調(diào)節(jié)蛋白家族中信號蛋白,含有表皮生長因子樣結(jié)構(gòu)域,通過與其下游ErbB2、ErbB3 和ErbB4 受體結(jié)合,介導(dǎo)神經(jīng)元遷移分化、髓鞘形成、神經(jīng)肌肉接頭發(fā)育,參與調(diào)節(jié)神經(jīng)元興奮性、突觸塑形、神經(jīng)傳遞,以及維持腦穩(wěn)態(tài),并在學(xué)習(xí)和記憶過程中發(fā)揮重要作用[21]。有研究發(fā)現(xiàn),鞘內(nèi)注射NRG1蛋白可減輕SCI后炎癥反應(yīng),減少硫酸軟骨素聚糖蛋白生成,抑制損傷區(qū)膠質(zhì)瘢痕形成,促進(jìn)SCI后大鼠神經(jīng)功能恢復(fù)[22]。
研究發(fā)現(xiàn),BMSCs 可合成包括外泌體在內(nèi)的多種細(xì)胞因子,抑制損傷局部炎癥反應(yīng)、細(xì)胞凋亡,進(jìn)而促進(jìn)SCI后神經(jīng)功能修復(fù)[23]。有臨床研究報道,經(jīng)椎管內(nèi)移植或靜脈輸注BMSCs,2 年后核磁共振成像顯示SCI 患者脊髓組織結(jié)構(gòu)改善,肢體感覺和運(yùn)動功能明顯恢復(fù),且無感染、疼痛及腫瘤形成等并發(fā)癥發(fā)生[24]。體視學(xué)檢查發(fā)現(xiàn),經(jīng)靜脈移植BMSCs 可阻止SCI 后脊髓灰質(zhì)和白質(zhì)體積減小,抑制損傷區(qū)神經(jīng)元凋亡[25]。本研究HE 染色和Nissl 染色結(jié)果顯示,NRG1-BMSCs 移植后大鼠脊髓組織灰質(zhì)和白質(zhì)結(jié)構(gòu)清晰,壞死區(qū)明顯縮小,脊髓灰質(zhì)中神經(jīng)細(xì)胞形態(tài)清晰可見,大部分神經(jīng)元形態(tài)結(jié)構(gòu)正常,提示移植NRG1-BMSCs 有利于抑制SCI 引起的核固縮,恢復(fù)受損神經(jīng)元尼氏體密度,減輕SCI 誘導(dǎo)的神經(jīng)元損傷,進(jìn)而促進(jìn)SCI 后神經(jīng)電生理功能恢復(fù)。有研究發(fā)現(xiàn),BMSCs 移植結(jié)合超短波治療可抑制SCI 后空洞形成,減輕損傷組織炎癥反應(yīng),促進(jìn)大鼠運(yùn)動誘發(fā)電位恢復(fù)[26]。本研究通過原位移植NRG1-BMSCs治療SCI,經(jīng)BBB運(yùn)動功能評分和斜板實驗觀察顯示,NRG1-BMSCs能明顯改善大鼠SCI后肢體運(yùn)動功能。由此推測,移植攜帶外源性NRG1基因的BMSCs 可能通過改善損傷局部微環(huán)境,抑制細(xì)胞凋亡過程,從而促進(jìn)大鼠SCI 后肢體運(yùn)動功能恢復(fù)。
研究發(fā)現(xiàn),內(nèi)質(zhì)網(wǎng)應(yīng)激介導(dǎo)的細(xì)胞凋亡參與SCI的病理過程[27]。內(nèi)質(zhì)網(wǎng)是真核生物細(xì)胞中一種重要細(xì)胞器,具有合成、修飾和轉(zhuǎn)運(yùn)蛋白質(zhì)的功能。內(nèi)質(zhì)網(wǎng)應(yīng)激為細(xì)胞的一種防御和自我保護(hù)機(jī)制,但劇烈而持久的內(nèi)質(zhì)網(wǎng)應(yīng)激反應(yīng)可導(dǎo)致細(xì)胞損傷甚至凋亡[28-29]。在SCI過程中,機(jī)體在炎癥、氧化應(yīng)激或其他致病因素刺激下,內(nèi)質(zhì)網(wǎng)功能可能受到影響,導(dǎo)致錯誤折疊蛋白在內(nèi)質(zhì)網(wǎng)中堆積,繼而誘發(fā)內(nèi)質(zhì)網(wǎng)應(yīng)激反應(yīng)。研究發(fā)現(xiàn),當(dāng)內(nèi)質(zhì)網(wǎng)應(yīng)激持續(xù)存在時,機(jī)體可通過啟動內(nèi)質(zhì)網(wǎng)信號途徑激活凋亡相關(guān)因子而誘發(fā)細(xì)胞凋亡[30]。蛋白激酶類RNA 內(nèi)質(zhì)網(wǎng)激酶(protein kinase RNA-like endoplasmic reticulum kinase,PERK)是位于內(nèi)質(zhì)網(wǎng)膜上的Ⅰ型跨膜蛋白,也是參與機(jī)體適應(yīng)性應(yīng)答的關(guān)鍵蛋白受體。正常生理條件下,PERK與GRP78結(jié)合處于無活性狀態(tài);當(dāng)內(nèi)質(zhì)網(wǎng)應(yīng)激發(fā)生時,PERK與GRP78解離磷酸化而激活,促使eIF2α 活化并上調(diào)ATF4 的表達(dá),進(jìn)而促進(jìn)CHOP蛋白合成,最終激活下游信號分子轉(zhuǎn)導(dǎo),誘發(fā)細(xì)胞凋亡[31]。研究發(fā)現(xiàn),內(nèi)質(zhì)網(wǎng)應(yīng)激在SCI 早期即被激活,在脊髓組織中內(nèi)質(zhì)網(wǎng)應(yīng)激相關(guān)蛋白GRP78、PERK、eIF2α、CHOP表達(dá)明顯上調(diào)[32],與本研究結(jié)果一致。本研究發(fā)現(xiàn),SCI 大鼠脊髓組織中GRP78、ATF4、CHOP、XBP1、ATF6 蛋白表達(dá)水平明顯增高,經(jīng)NRG1-BMSCs 治療后,脊髓組織中內(nèi)質(zhì)網(wǎng)應(yīng)激相關(guān)蛋白表達(dá)水平明顯降低,提示NRG1-BMSCs能夠調(diào)控內(nèi)質(zhì)網(wǎng)應(yīng)激信號通路,減輕內(nèi)質(zhì)網(wǎng)應(yīng)激引起的組織損傷,進(jìn)而發(fā)揮神經(jīng)保護(hù)作用。Bax基因是機(jī)體中主要的凋亡基因,具有促進(jìn)細(xì)胞凋亡的作用。Bcl-2 主要定位于線粒體外膜上,通過調(diào)控線粒體膜通透性,發(fā)揮抑制細(xì)胞凋亡的作用[33]。本研究結(jié)果表明,NRG1-BMSCs 移植能夠減少脊髓損傷區(qū)TUNEL 陽性細(xì)胞數(shù),同時降低脊髓組織中Bax 蛋白水平,提高Bcl-2 蛋白水平。由此推測,NRG1-BMSCs 可能通過調(diào)節(jié)內(nèi)質(zhì)網(wǎng)應(yīng)激介導(dǎo)的凋亡途徑,抑制SCI誘導(dǎo)的細(xì)胞凋亡。
綜上所述,移植NRG1-BMSCs能夠促進(jìn)大鼠SCI后運(yùn)動功能恢復(fù),其作用機(jī)制可能包括:(1)NRG1促進(jìn)BMSCs 在脊髓組織內(nèi)增殖、存活及遷移,參與受損神經(jīng)組織修復(fù),進(jìn)而促進(jìn)大鼠運(yùn)動功能恢復(fù);(2)NRG1-BMSCs 參與改善損傷局部微環(huán)境,促進(jìn)軸突再生和髓鞘形成;(3)NRG1-BMSCs 可能通過分泌NRG1 及多種神經(jīng)營養(yǎng)因子,促進(jìn)神經(jīng)組織結(jié)構(gòu)重構(gòu);(4)NRG1-BMSCs 可能通過調(diào)節(jié)內(nèi)質(zhì)網(wǎng)應(yīng)激介導(dǎo)的凋亡通路,抑制細(xì)胞凋亡。本研究結(jié)果表明,移植NRG1-BMSCs 能夠減輕SCI,改善大鼠運(yùn)動功能,為其用于臨床治療SCI 提供了實驗依據(jù)。但本研究未深入探討NRG1-BMSCs 對大鼠脊髓組織分化、軸突再生及髓鞘化、膠質(zhì)細(xì)胞增生和膠質(zhì)瘢痕形成的影響,以及對脊髓損傷局部血管形成和炎癥反應(yīng)的作用,后續(xù)需進(jìn)一步探討NRG1-BMSCs 的神經(jīng)保護(hù)作用及分子機(jī)制。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網(wǎng)絡(luò)安全與數(shù)據(jù)管理(2022年1期)2022-08-29 03:15:20
導(dǎo)航定位學(xué)報(2022年4期)2022-08-15 08:27:00
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:36
成都醫(yī)學(xué)院學(xué)報(2021年2期)2021-07-19 08:35:14
新世紀(jì)智能(數(shù)學(xué)備考)(2020年9期)2021-01-04 00:25:14
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
