西藏油桃褐腐病菌生物學特性測定及室內藥劑篩選
2024-03-28 07:58:24孫晨曦毛維興相棟徐秉良張樹武
西北農業學報 2024年1期
孫晨曦 毛維興 相棟 徐秉良 張樹武
摘 要 旨在明確西藏油桃上褐腐病菌(Monilia yunnanensis)的生物學特性及6種殺菌劑對其菌落生長的室內毒力。通過“菌落生長速率法”評價不同培養基成分溫度、光照、pH及殺菌劑對M.yunnanensis菌落生長的影響。結果表明,在固體培養基上培養7 d后,M.yunnanensis最適生長碳源為淀粉,其生長速率為? 0.696 cm/d,其次是乳糖和蔗糖,其生長速率分別為0.558和0.417 cm/d,在葡萄糖和山梨醇上的生長速率為0.343和0.328 cm/d,但在甘露醇培養基上具有顯著的抑制作用;最適生長氮源為苯丙氨酸,生長速率為0.374 cm/d,蛋氨酸、硝酸銨及硝酸鈉次之,其生長速率分別為0.290、0.248和0.246 cm/d,但是硫酸銨和尿素對其生長具有抑制作用;最適光照條件為全黑暗,其菌絲生長速率為0.721 cm/d,較其他光照條件具有顯著性差異;最適溫度20~25 ℃,但當溫度高于40 ℃則不能正常生長;最適pH為6~8。室內毒力測定結果表明,6種殺菌劑對西藏油桃褐腐病菌生長均具有不同的抑制效果,其中丁子香酚和戊唑醇的抑菌效果最佳,其EC50值分別為0.032和0.043? μg/mL。
關鍵詞 西藏;油桃褐腐病;生物學特性;殺菌劑;生長速率
西藏自治區位于青藏高原西南部,是中國氣候資源最多樣與果樹資源最為豐富的區域之一[1]。油桃是西藏主要的水果種類之一,截止2016年,桃樹栽培面積和產量分別占全自治區的18%和16%,居水果類第二位[2]。西藏油桃具有口感香甜、肉質細脆、耐貯運及耐長途運輸,且富含蛋白質、粗纖維、糖及礦物質等多種人體所必需的物質, 被稱為“天下第一果”[3],但是隨著油桃樹栽培面積的不斷增加,油桃褐腐病的發生愈來愈嚴重,對西藏地區油桃產業的發展造成了嚴重影響。桃褐腐病是一種能嚴重危害核果和仁果類水果生產的真菌性病害[4],是桃樹上發生的一種常見病害[5],并且其可在桃樹整個生長期進行危害,不僅造成花腐、果腐和枝條潰瘍等癥狀,導致果樹產量和質量下降,還可引起儲藏期果實腐爛,造成嚴重的經濟損失[6],嚴重時還對翌年桃樹成花坐果不利,對桃樹造成毀滅性損失[7]。
目前,世界上已報道的引起果樹褐腐病的鏈核盤菌屬親緣關系較近的有6個種[8],中國的褐腐病菌種群與歐美種群有所不同,目前中國已確定的種有4個,主要3個種為Monilinia fructicola、Monilia mumecola和Monilia? yunnanensis[9-10],其中M.yunnanensis主要分布于云南、北京、陜西和沈陽等地區[4,9],植物病害生物防治課題組前期研究發現引起了西藏油桃褐腐病的病原為M.yunnanensis,首次報道了其造成西藏油桃褐腐病[11],但是關于油桃褐腐菌生物學特性的研究較少。同時,目前關于桃褐腐病的防治仍以化學手段為主,但果園的噴藥量沒有一定的規范標準,噴施藥物量大且種類雜,田間防效參差不齊[12],導致了桃表面農藥殘留超標、病菌對多種藥物產生抗性、危害人體健康的同時還對環境造成了污染[13]。因此,篩選出低毒高效的化學藥劑和安全高效的植物源藥劑是防治該病害的一種重要方法。目前有關西藏地區油桃褐腐菌生物學特性及其藥劑篩選尚未報道。因此,本試驗開展了西藏地區油桃褐腐病菌的生物學特性測定,以及其植物源和低毒高效化學藥劑的篩選,旨在明確西藏地區油桃褐腐病菌的生物學特性,并篩選獲得防效較好的藥劑,為西藏地區油桃褐腐病菌的防治提供理論依據。
1 材料與方法
1.1 供試菌株
以課題組前期分離自西藏林芝市巴宜區,林芝鎮圣域公司示范園區油桃褐腐病病害標本的病原菌Monilia yunnanensis作為供試材料。
1.2 供試培養基
PDA培養基:去皮馬鈴薯200 g、葡萄糖? 20 g、瓊脂15 g、蒸餾水1? 000 mL。
查氏(Czapek)培養基:KNO3 2 g、MgSO4·7H2O 0.5 g、FeSO4 0.01 g、KCL 0.5 g、KH2PO4 1 g、瓊脂15 g、蔗糖30 g、蒸餾水1? 000 mL。
1.3 供試藥劑
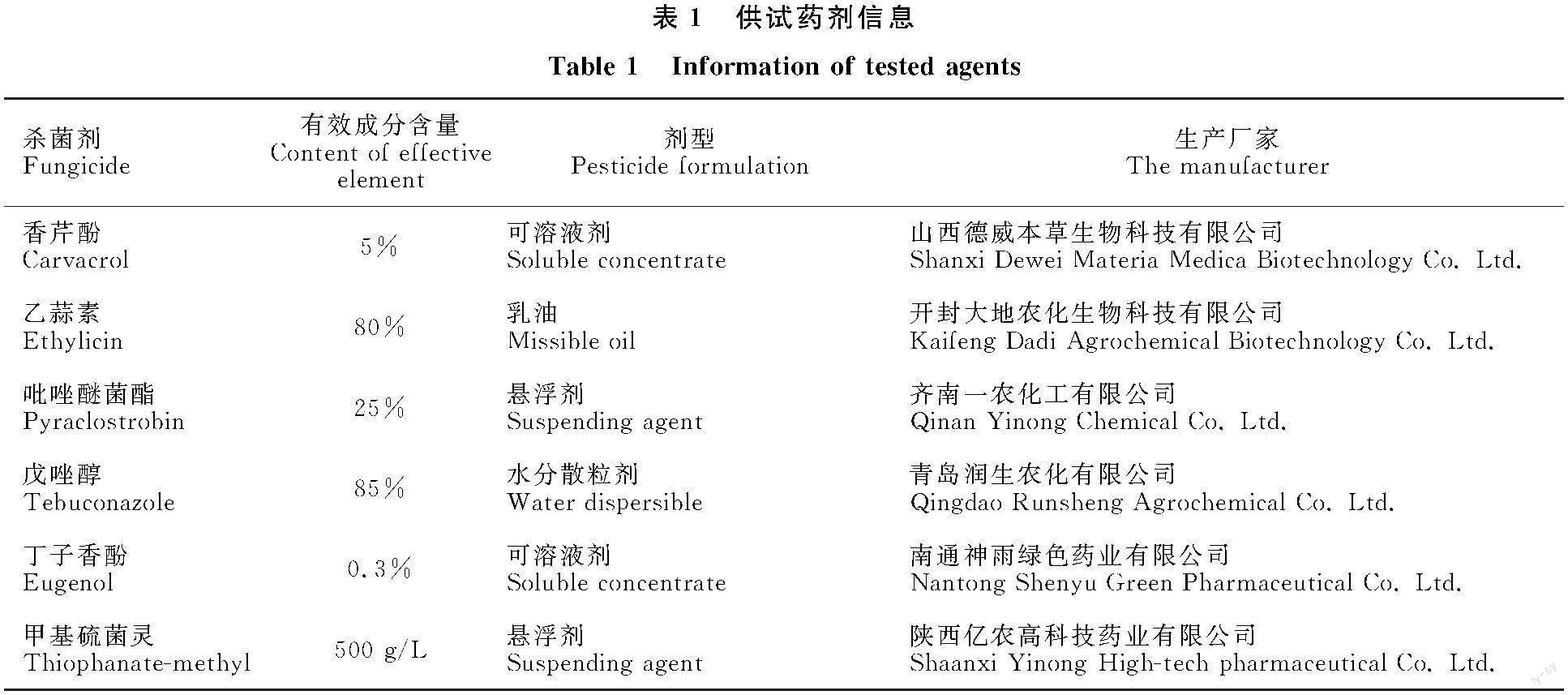
藥劑篩選中供試藥劑見表1。
1.4 試驗方法
1.4.1 Monilia yunnanensis活化與培養 將? 4 ℃低溫保存于斜面培養基上的油桃褐腐病菌接種于PDA培養基,并置于溫度為25 ℃,光照時間為16/8 h的培養箱中進行活化培養,培養3 d后備用。
1.4.2 不同碳源對Monilia yunnanensis生長的影響 試驗以查氏培養基作為供試培養基,供試碳源選用等質量的葡萄糖、乳糖、甘露醇、蔗糖、可溶性淀粉和山梨醇代替查氏培養基中的蔗糖,測定不同碳源對油桃褐腐病菌生長的影響。試驗以不加碳源的查氏培養基作為對照,每個處理和對照分別重復6次。待接種培養后,每隔1 d采用“十字交叉法”測定M.yunnanensis的菌落直徑,并計算其生長速率。
1.4.3 不同氮源對Monilia yunnanensis生長的影響 試驗以查氏培養基作為供試培養基,供試氮源選用質量相等的尿素、苯丙氨酸、硝酸銨、硝酸鈉、硫酸銨和蛋氨酸替代查氏培養基中的NaNO3,測定不同氮源對油桃褐腐病菌生長的影響。試驗以不加氮源的查氏培養基作為對照培養基,每個處理和對照分別重復6次。待接種培養后,每隔1 d采用“十字交叉法”測定M.yunnanensis菌落直徑,并計算其生長速率。
1.4.4 不同溫度對Monilia yunnanensis生長的影響 試驗以PDA培養基作為供試培養基,供試溫度條件選用5 ℃、15 ℃、20 ℃、25 ℃、28 ℃、30 ℃、35 ℃和40 ℃,測定不同溫度對油桃褐腐病菌生長的影響。試驗每個處理和對照分別重復6次。待接種培養后,每隔1 d采用“十字交叉法”測定M.yunnanensis菌落直徑,并計算其生長? 速率。
1.4.5 不同pH對Monilia yunnanensis生長的影響 試驗以PDA培養基作為供試培養基,供試pH條件分別設為4、5、6、7、8和9,測定不同pH對油桃褐腐病菌生長的影響。試驗每個處理和對照分別重復6次。待接種培養后,每隔1 d采用“十字交叉法”測定M.yunnanensis的菌落直徑,并計算其生長速率。
1.4.6 不同光照對Monilia yunnanensis生長的影響 試驗以PDA培養基作為供試培養基,供試光照處理條件分別設為25 ℃持續光照24 h、? 25 ℃持續黑暗24 h和25 ℃光照/黑暗交替處理 (12/12 h)。試驗每個處理和對照分別重復6次。待接種培養后,每隔1 d采用“十字交叉法”測定M.yunnanensis菌落直徑,并計算其生長速率。
1.4.7 6種殺菌劑對Monilia yunnanensis的室內毒力測定 試驗供試藥劑香芹酚的濃度梯度設置為1、2、4、8、16 mg/L;乙蒜素的濃度梯度設置為1、2、4、8、16 mg/L;吡唑醚菌酯的濃度梯度設置為0.15、0.3、0.6、1.2、2.4 mg/L;戊唑醇的濃度梯度設置為0.01、0.02、0.04、0.08、0.1 mg/L;丁子香酚的濃度梯度設置為0.02、0.03、0.04、? 0.05、0.06 mg/L;甲基硫菌靈的濃度梯度設置為? 0.08、0.16、0.24、0.32、0.40 mg/L。將不同農藥分別配制成10 mL其濃度梯度10倍的濃縮液,然后將濃縮液分別加入到90 mL PDA中,制成相應濃度梯度的含藥培養基。以加入10 mL無菌水于90 mL PDA中制成培養基作為對照培養基。然后,利用經滅菌的直徑為0.5 cm打孔器打取活化3 d的M.yunnanensis菌餅,分別接種于各含藥培養基中央。試驗以PDA培養基作為對照,每個處理重復6次,并置于溫度為25 ℃,光照時間為16/8 h的溫培養箱中培養。待培養7 d后,采用“十字交叉法”測定不同含藥培養基上菌落直徑,并計算不同藥劑對M.yunnanensis菌落生長抑制率。以濃度對數值(x)與抑制率幾率值(y)之間的線性回歸關系求出毒力回歸方程和EC50值[14]。參考紀兆林等[12]文獻評價殺菌劑毒力的方法,根據不同藥劑EC50值評價其殺菌劑毒力:EC50<0.01 μg/mL,很強;EC50為0.01~0.1 μg/mL,強;EC50為 0.1~1.0 μg/mL,較強;EC50為1.0~5.0 μg/mL,中等;EC50為5.0~10.0 μg/mL,較弱;EC50>10.0 μg/mL,弱。
1.5 數據處理
利用Microsoft Excel 2016軟件進行數據處理與圖表制作,并采用SPSS 25.0軟件進行Probit分析與卡方檢驗。采用單因素方差分析各處理平均數的差異及其差異顯著性檢驗(Duncan新復極差法)。
2 結果與分析
2.1 不同碳源對Monilia yunnanensis生長的? 影響
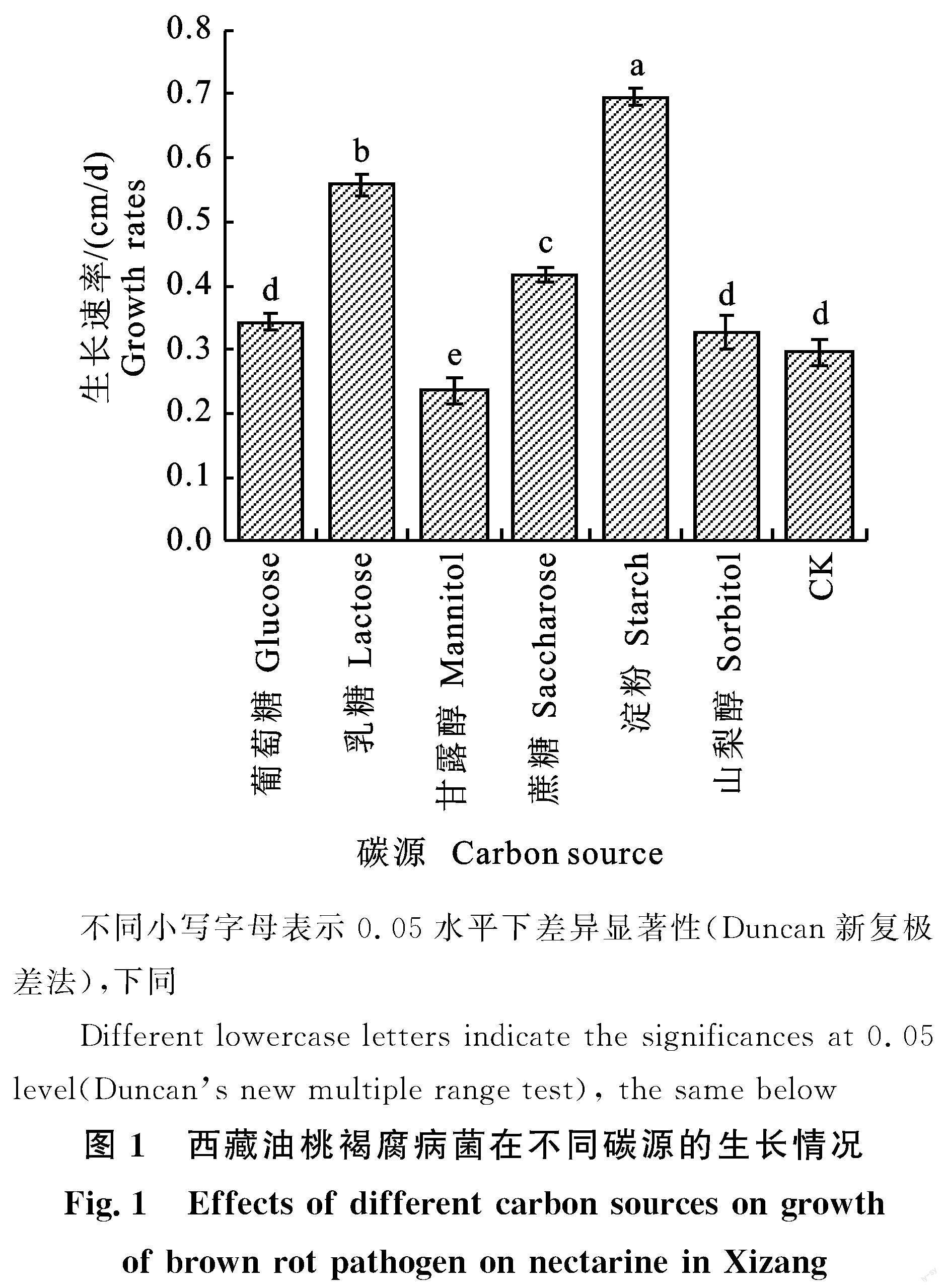
結果發現,培養7 d后,以葡萄糖、乳糖、甘露醇、蔗糖、可溶性淀粉和山梨醇等作為唯一碳源培養的M.yunnanensis生長速率具有不同程度差異。與對照相比,淀粉、乳糖和蔗糖對M.yunnanensis菌落的生長具有顯著的促進作用,其中以淀粉作為唯一碳源的培養基對其菌落的生長促進作用最顯著,其生長速率為0.696 cm/d,而以葡萄糖和山梨醇作為唯一碳源的培養基促進作用則不顯著,其生長速率分別為0.343 cm/d和? 0.328 cm/d。另外,當以甘露醇作為唯一碳源的培養基對其菌落生長具有顯著的抑制作用,其生長速率為0.235 cm/d(圖1)。
2.2 不同氮源對Monilia yunnanensis生長的影響
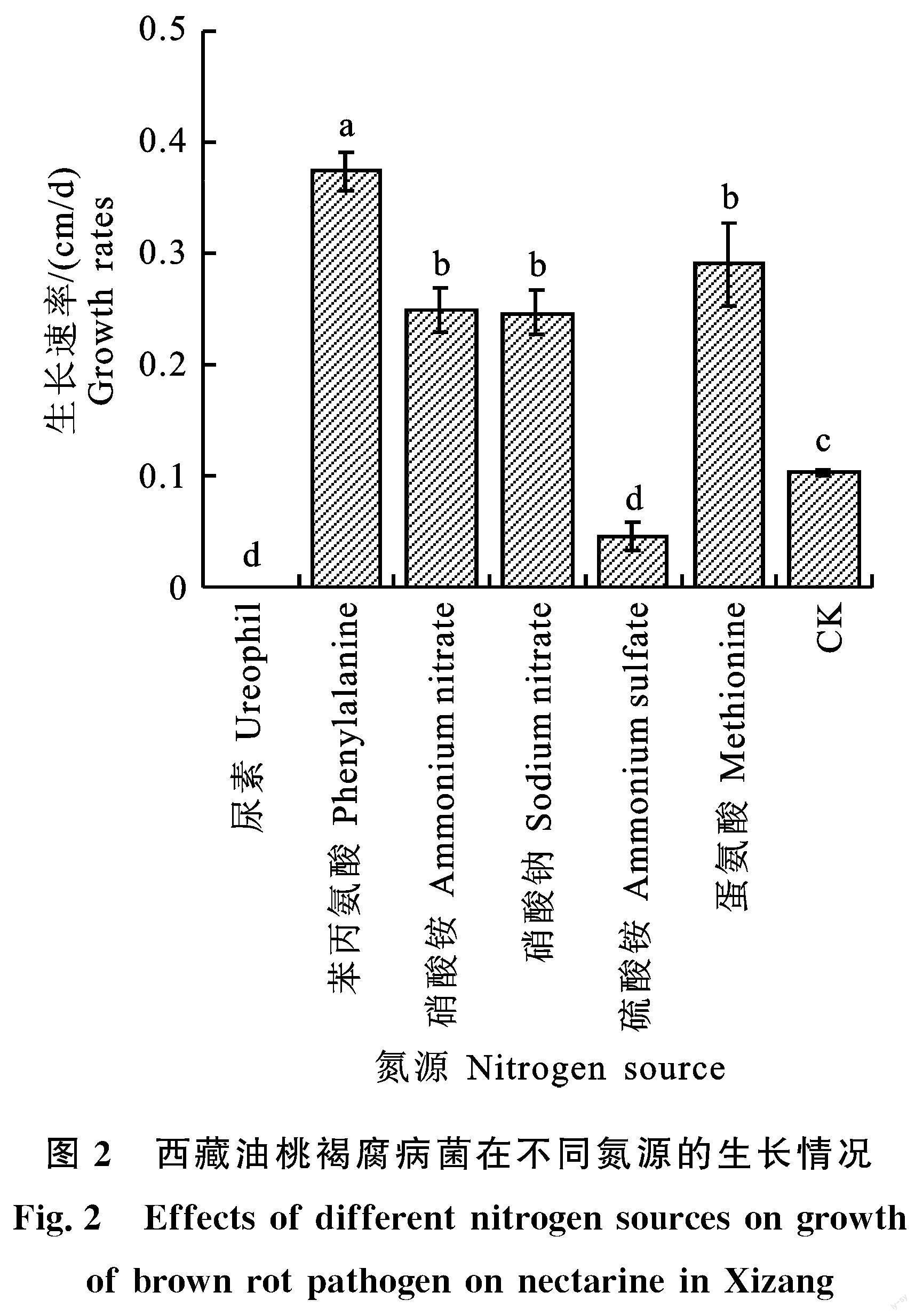
與對照相比,培養7 d后,以苯丙氨酸、蛋氨酸、硝酸銨、硝酸鈉作為唯一氮源的培養基對M.yunnanensis菌落生長具有明顯的促進作用,其中苯丙氨酸促進作用顯著高于蛋氨酸、硝酸銨和硝酸鈉,其生長速率為0.374 cm/d。同時,與對照相比,硫酸銨和尿素對M.yunnanensis菌落的生長具有顯著抑制作用,且尿素抑制作用最強,使得菌落不能正常生長,其次為為硫酸銨,其生長速率為0.045 cm/d(圖2)。
2.3 不同溫度對Monilia yunnanensis生長的影響
結果表明,M.yunnanensis菌落生長速率隨著溫度(5~40 ℃)的升高呈先增加后降低的趨勢,并且不同溫度對其生長速率的影響存在顯著差異。在5~20 ℃的條件下,隨著溫度的升高其生長速率逐漸上升, 培養7 d后其在5 ℃、10 ℃、15 ℃和20 ℃時生長速率分別為0.168、0.391、? 0.686 和1.071 cm/d,尤其在20 ℃條件下菌落生長速率最大,并且與其它溫度相比差異顯著。然而,在25 ℃、28 ℃、30 ℃、35 ℃和40 ℃條件下,其生長速率隨著溫度的升高逐漸降低,生長速率分別為0.986、0.843、0.631、0.419和0 cm/d? (圖3)。
2.4 不同光照對Monilia yunnanensis生長的影響
結果表明,不同光照條件對M.yunnanensis菌落的生長速率具有不同程度的影響,待培養7 d后,在全黑暗條件下M.yunnanensis菌落生長最快,其生長速率為0.721 cm/d,與全光照和光照交替條件下菌落生長速率相比,全黑暗條件下其菌落生長速率顯著高于全光照和光照交替條件,分別增長了0.116和0.059 cm/d。然而,全光照條件下M.yunnanensis菌落生長速率較低,僅為0.605 cm/d。從表2可以看出,每天的光照條件隨著黑暗時間的增長,其菌落生長速率增高。
2.5 不同pH對Monilia yunnanensis生長的影響
結果表明,不同pH對M.yunnanensis菌落的生長速率具有不同程度的影響。培養7 d后,當pH為6~8時其菌落生長速度較快,尤其當pH為6時最佳為0.982 cm/d。然而,當pH=5和9時其生長速率較低,分別為0.713和0.681 cm/d(表3)。
2.6 不同殺菌劑對Monilia yunnanensis的室內毒力測定
毒力測定結果表明,生物源殺菌劑丁子香酚對M.yunnanensis菌落生長的抑制作用最強,其EC50為0.032 μg/mL。其次為化學殺菌劑戊唑醇,其EC50為0.043 μg/mL。甲基硫菌靈和吡唑醚菌酯的抑菌能力為較強,其EC50分別為0.294和0.467 μg/mL。香芹酚抑菌能力較弱,其EC50為7.84 μg/mL。乙蒜素的抑菌能力最弱,其EC50為17.944 μg/mL。
根據其毒力回歸方程斜率可以看出,M.yunnanensis對供試6種殺菌劑較為敏感性,其中對甲基硫菌靈的敏感性最高,其斜率為3.734,其余殺菌劑的斜率為1.029~2.409。同時,試驗中6種殺菌劑對M.yunnanensis生長的抑制作用與其濃度呈正相關(表4)。
3 討? 論
前期對果實褐腐病菌的研究主要集中在櫻桃[15]、桃[16]、蘋果[17]等果實上,有關油桃褐腐菌的報道與研究相對較少,而西藏油桃褐腐菌生物學特性的相關研究還未見報道。本試驗發現Monilia yunnanensis在固體選擇培養基上最適碳源為淀粉,這與紀兆林等[18]對不同產區的桃褐腐病菌的生物學特性進行探究,發現其菌絲生長的最適生長碳源為淀粉結果一致。同時,本試驗發現Monilia yunnanensis在固體培養基上最適生長氮源為苯丙氨酸,在溫度為5~35 ℃的條件下均能生長,其生長特性整體呈現出先升高再降低的趨勢,高于40 ℃則不能生長,其最適生長溫度為20~25 ℃,這與宋丹丹等[19]研究結果基本一致,其表明蟠桃褐腐病菌Monilinia? fructicola菌絲的最適生長溫度范圍20~28 ℃,但是高于36 ℃時則不能正常生長。與郝曉娟等[20]研究發現的歐李褐腐病菌Monilinia? fructicola菌絲在? 7~27 ℃均能正常生長稍有差異,其原因可能與病原菌種類和菌株分離來源的地域氣候條件有關。Monilia yunnanensis最適的光照條件為全黑暗,這與劉志恒等[15]研究發現大櫻桃褐腐病菌Monilinia? fructigena在光照、光暗交替、黑暗3種條件下菌絲都能正常生長,但在黑暗條件下菌絲生長稍快的結果一致。Monilia yunnanensis最適生長pH范圍在6~8之間,且在中性及弱酸堿性條件下生長速度稍快,這與郝曉娟等[20]發現在pH 6~8時,適宜于歐李褐腐病菌孢子萌發的結果基本一致。
另外,本試驗發現丁子香酚和戊唑醇對Monilia yunnanensis菌落生長抑制作用效果最佳,但甲基硫菌靈對Monilia yunnanensis菌落生長抑制作用效果低于戊唑醇,這可能與Monilia yunnanensis對DMI類殺菌劑和苯并咪唑類藥劑產生抗性有關[21-22]。丁子香酚作為一種生物源藥劑,不僅對Monilia yunnanensis菌的生長抑制效果最好,并且在自然環境中極易分解轉化,對環境友好,對人畜安全,是一種理想的殺菌劑。本研究所用殺菌劑為不同劑型的制劑,相較于原藥有更好的田間應用價值,但制劑不同的助劑和劑型都可能影響測定結果,其影響有待進一步研究。本試驗為幾種殺菌劑對Monilia yunnanensis的室內毒力測定,而在田間會受到氣候條件等多種不可控因素的影響,因此在田間的應用效果還需要通過田間試驗進一步探究。
參考文獻 Reference:
[1] 曾秀麗,扎 西,王玉霞,等.西藏桃生產的現狀和問題[J].西藏科技,2012(6):14-15.
ZENG X L,ZHA X,WANG Y X,et al.The current situation and problems of peach production in Tibet[J].Tibet Science and Technology,2012(6):14-15.
[2] 曾秀麗,張姍姍,鄧 嵐,等.3個油桃品種在拉薩露地栽培的果實發育初探[J].西藏農業科技,2016,38(4):38-41.
ZENG X L,ZHANG SH SH,DENG L,et al.Growth and development patterns of three nectarines in Lhasa[J].Tibet Journal of Agricultural Sciences,2016,38(4):38-41.
[3] 殷曉慧,王慶國,張 暢,等.桃果實褐腐病拮抗菌的篩選、鑒定及其拮抗活性[J].食品工業科技,2017,38(9):128-132.
YIN X H,WANG Q G,ZHANG CH,et al.Isolation and?? identification of antagonistic bacteria against peach brown rot and its antibacterial activity[J].Science and Technology of Food Industry,2017,38(9):128-132.
[4] 羅朝喜.果樹褐腐病的研究現狀及其展望[J].植物病理學報,2010,5(2):4-12.
LUO CH X.Advances and prospects on researches of brown rot disease on fruits[J].Acta Phytopathologica Sinica,2010,5(2):4-12.
[5] 李世訪,陳 策.桃褐腐病的發生和防治[J].植物保護,2009,35(2):134-139.
LI SH F,CHEN C.Incidence and management of the peach fruit brown rot[J].Plant Protection,2009,35(2):134-139.
[6] BYTHER R S.Brown rot of stone fruits[J].Extension Bulletin-Cooperative Extension,College of Agriculture & Home Economics,Washington State University,1991,1047:2.
[7] 馬靖艷.桃褐腐病綜合防控技術[J].林業與生態,2021(2):41-42.
MA J Y.Comprehensive prevention and control technology of peach brown rot[J].Forestry and Ecology,2021(2):41-42.
[8] 尹良芬.中國核果/仁果類果樹褐腐病菌種群結構及Cyt b基因的遺傳進化研究[D].武漢:華中農業大學,2015.
YIN L F.Characterization of populations of Monilinia spp.on stone and pome fruits in China and genetic evolution of the Cyt b genes[D].Wuhan:Huazhong Agricultural University,2015.
[9] HU M J,COX K D,SCHNABEL G,et al.Monilinia species causing brown rot of peach in China [J].PLoS One,2011,? 6(9):e24990.
[10]YIN L F,CHEN S,CHEN G K,et al.Identification and characterization of three Monilinia species from plum in China [J].Plant Disease,2015,99:1775-1783.
[11] ZHANG S W,XIANG D,LI T,et al.First report of brown rot of nectarine caused by Monilia yunnanensis in Tibet[J].Plant Disease,2021.
[12] 紀兆林,蔣長根,戴慧俊,等.不同殺菌劑對桃褐腐病菌的毒力測定[J].中國南方果樹,2013,42(5):95-97.
JI ZH L,JIANG CH G,DAI H J,et al.Toxicity of different fungicides to peach brown rot fungus[J].South China Fruits,2013,42(5):95-97.
[13] 莫熙禮,趙同貴,鄧 偉,等.花椒提取物對桃褐腐病菌的抑制作用及酶活性的影響[J].北方園藝,2014(20):129-132.
MO X L,ZHAO T G,DENG W,et al.Effect of zanthoxylum bungeanum on pathogen inhibition and activities of antioxidative enzyme of Monilinia fructicolain peach[J].Northern Horticulture,2014(20):129-132.
[14] 周 瑩,嚴 紅, 關海春,等.幾種殺菌劑對桃褐腐病的毒力測定及田間控制作用[J].農藥, 2013, 52(10):771-777.
ZHOU Y,YAN H,GUAN H CH,et al.Toxicity and field effi ciency of several fungicides against peach brown rot[J].Agrochemicals,2013,52(10):771-777.
[15] 劉志恒,白海濤,楊 紅,等.大櫻桃褐腐病菌生物學特性研究[J].果樹學報,2012,29(3):423-427.
LIU ZH H,BAI H T,YANG H,et al.Biological characteristics of Monilia fructigena as pathogen of brown rot in sweet cherry[J].Journal of Fruit Science,2012,29(3):423-427.
[16] 紀兆林,談 彬,朱 薇,等.我國不同產區桃褐腐病病原鑒定與分析[J].微生物學通報,2019,46(4):869-878.
JI ZH L,TAN B,ZHU W,et al.Identification and analysis of the peach brown rot pathogens from different peach-growing areas in China[J].Microbiology China,2019,?? 46(4):869-878.
[17] 岳海梅, 莊 華, 鞏文峰,等.蘋果褐腐病拮抗毛殼菌的篩選及鑒定[J].果樹學報,2017,34(8):1025-1032.
YUE H M,ZHUANG H,GONG W F,et al.Screening and identification of antagonistic Chaetomium spp. against Monilinia fructigena[J].Journal of Fruit Science,2017,34(8):1025-1032.
[18] 紀兆林,朱 薇,談 彬,等.我國不同產區桃褐腐病菌生物學特性研究[J].中國南方果樹,2018,47(5):101-107.
JI ZH? L,ZHU W,TAN B,et al.Biological characteristics of peach brown rot pathogens from different peach-growing areas in China[J].South China Fruits,2018,47(5):101-107.
[19] 宋丹丹,孔慶軍,李鴻彬,等.蟠桃褐腐病原菌生物學特性及細胞壁降解酶活性[J].食品科學,2017,38(10):42-48.
SONG D D,KONG Q J,LI H B,et al.Biological characteristics and cell wall degrading enzyme activities of flat peach brown rot pathogen[J].Food Science,2017,? 38(10):42-48.
[20] 郝曉娟,高 瑩,李新鳳,等.歐李褐腐病病原菌生物學特性及其寄主范圍[J].果樹學報,2014,? 31(1):101-104.
HAO X J,GAO Y,LI X F,et al.Biological characteristics and host range of the pathogen causing brown rot of Chinese dwarf cherry[J].Journal of Fruit Science,2014,31(1):101-104.
[21] CHEN F P,LIU X L,CHEN S,et al.Characterization of Monilinia fructicola strains resistant to both Propiconazole and Boscalid[J].Plant Disease,2013,97:645-651.
[22] 張艷婷,仇智靈,李阿根,等.浙江省櫻桃褐腐病病原菌種類及其對常見藥劑的抗性檢測[J].果樹學報,2020,37(9):1394-1403.
ZHANG Y T,QIU ZH? L, LI A G,et al.Species of pathogens causing cherry brown rot and their resistance to common fungicides in Zhejiang province[J].Journal of Fruit Science,2020,37(9):1394-1403.
Determination of Biological Characteristics of Nectarine Brown Rot Fungus in Xizang and Indoor Pharmacy Screening
Abstract In order to clarify the biological characteristics of the pathogen of nectarine brown rot (Monilia yunnanensis) in Xizang, and determine the toxic effect of six fungicides on its colony growth, we evaluated the effect of different composition of culture media and conditions as well as fungicides on the M.yunnanensis colony growth by the method of colony growth rate.The results showed that the most suitable carbon source for M.yunnanensis growth was starch with a growth rate of 0.696 cm/d on the solid medium at 7 days after incubation, followed by lactose, sucrose, glucose and sorbitol, and with the growth rates of 0.558, 0.417, 0.343 and 0.328 cm/d, respectively. In contrast, a significant inhibitory effect was observed on the mannitol medium; the most suitable nitrogen source was phenylalanine, and the growth rate was 0.374 cm/d, followed by methionine, ammonium nitrate and sodium nitrate, and the growth rates were 0.290, 0.248 and 0.246 cm/d, respectively. However, the ammonium sulfate and urea had significant inhibitory effect on M.yunnanensis growth; the most suitable light condition was darkness, and the colony growth rate was 0.721 cm/d, which was significantly different from other light conditions; the optimum temperature was 20 to 25 ℃, whereas the temperature cannot grow normally was higher than 40 ℃; the optimum pH was 6-8. In addition, The test results also showed that the six fungicides had different inhibitory effects on the growth of?? M.yunnanensis. The eugenol and tebuconazole exhibited the best antifungal effect, and their EC50 values were 0.032? and 0.043? μg/mL, respectively.
Key words Xizang; Nectarine brown rot; Biological characteristics; Fungicides; Growth rate
