葡萄籽提取物、茶多酚可食性活性包裝膜對羅非魚的保鮮作用
2024-04-03 02:13:36蔣雨心孫悅溪楊曉玲范方宇
食品與發酵工業 2024年6期
蔣雨心,孫悅溪,楊曉玲,范方宇,2,3*
1(西南林業大學 生命科學學院,云南 昆明,650224)
2(西南地區生物多樣性保育國家林業和草原局重點實驗室,云南 昆明,650224)
3(云南省森林災害預警與控制實驗室,云南 昆明,650224)
羅非魚營養豐富、肉質鮮嫩、味道鮮美,廣受消費者喜愛,但因內源蛋白酶活性較強,在加工、運輸、貯藏等過程中易水解和氧化[1]。傳統的低溫保鮮技術易出現冰晶堆積,造成肌肉收縮及蛋白聚集變性等,導致魚肉品質變差[2]。通過添加天然抗氧化劑是抑制魚肉脂肪和蛋白質氧化的有效方法,但一些水溶性天然活性物質(如茶多酚)難以直接添加到油脂食品體系中,限制了其應用[3],為此,各種新型活性包裝膜應運而生。研究者們將天然抗氧化劑與可食性包裝膜結合制備活性包裝膜,可達到有效的抗氧化濃度閾值,并在食品與膜界面實現抗氧化、抑菌作用,保持食品新鮮度,延長貨架期[4]。傳統方法制備活性包裝膜主要以單一抗氧化劑為主,因其時效短、廣譜性差以及合成復雜等問題,使用過程中無法達到理想的效果。研究表明,通過將不同抗菌、抗氧化劑復配可得到兼具安全性、廣譜性、穩定性和高效抗菌性的復合型保鮮劑[2]。
葡萄籽提取物(grape seed extraction,GSE)是食品加工過程中的副產品,含豐富的酚類物質[5],其抗氧化能力分別是維生素E的20倍、維生素C的50倍,是廉價的天然抗氧化劑來源[6]。GSE抗氧化活性的多重機制主要表現在清除自由基的能力、金屬螯合作用以及與其他抗氧化劑的協同作用等方面。茶多酚(tea polyphenols,TP)類物質是茶葉中多羥基酚類化合物的總稱,具有抗氧化、抗輻射等多種功能,是一種新型的天然抗氧化、抗菌劑[7]。二者均在食品保鮮領域具有較高的應用價值。許寶琛等[5]在牛肉餡中添加茶多酚、葡萄籽提取物和海藻糖提高了肉餡的保水性,質構變化程度小,抑制了感官品質劣變。余小亮等[8]以茶多酚、肉桂精油復配,所制備出的復合保鮮劑具較強抗氧化活性,并對金黃色葡萄球菌有較好抑菌效果。都津銘等[9]將丁香精油和茶多酚復合得到復合抗菌液,結果表明二者以一定比例的復配具有協同抑菌效果。
目前,關于GSE和TP兩者復配制備可食性活性包裝膜的研究鮮有報道。基于此,本文以海藻酸鈉(sodium alginate,SA)、納米纖維(nanofiber,NCC)作為膜基材,GSE、TP復合制備可食性活性包裝用于羅非魚保鮮,研究貯藏過程中魚肉品質指標及肌原纖維蛋白氧化指標的變化,并分析復合膜對魚肉的保鮮效果,為新型可食性活性包裝膜的開發及魚肉的貯藏保鮮提供新途徑。
1 材料與方法
1.1 材料與試劑
新鮮羅非魚,云南省昆明市盤龍區農貿市場;葡萄籽提取物(食品級),西安齊岳生物科技有限公司;納米纖維(20~50 nm),中山納纖絲新材料有限公司;茶多酚(HPLC≥97%)、海藻酸鈉、硫代巴比妥酸(thiobarbituric acid,TBA)、2,4-二硝基苯肼、2-二硝基苯甲酸、溴酚藍、鹽酸胍、考馬斯亮藍、酒石酸鉀鈉、乙酸乙酯、三氯乙酸、乙二胺四乙酸二鈉均為分析純,上海麥克林生化科技有限公司;丙三醇、磷酸氫二鈉、磷酸二氫鉀、氯化鈉、甲基紅、亞甲基藍均為分析純,廣東光華科技股份有限公司;平板計數瓊脂,國藥集團化學試劑有限公司。
1.2 儀器與設備
FJ200-SH高速分散均質機,上海滬析實業有限公司;A300Plus電子攪拌器,上海歐河機械設備有限公司;HSP-80B恒溫恒濕箱,上海坤天實驗室儀器有限公司;101-2AB電熱鼓風干燥箱,天津市泰斯特儀器有限公司;TGL16M離心機,上海赫田科學儀器有限公司;UV-2600紫外可見分光光度計,日本島津公司。
1.3 實驗方法
1.3.1 可食性活性包裝膜的制備
取4.5 g NCC于燒杯中,加入50 mL蒸餾水,450 W超聲20 min得NCC懸浮液;同時取1.0 g SA于50 mL蒸餾水中,60 ℃,1 200 r/min攪拌均勻得SA溶液。將上述2種溶液以等體積比混勻后加入0.8% (質量分數)的復合保鮮溶液[m(GSE)∶m(TP)=2∶1,后文簡稱為GT],1 200 r/min磁力攪拌至混合均勻,超聲30 min并靜置消泡得成膜液。取15.0 g成膜液倒入9 cm聚四氟乙烯(poly tetra fluoroethylene,PTFE)皿中流延成膜,置于45 ℃烘箱干燥15 h得GT-SA-NCC復合膜。干燥后的膜樣品置于恒溫恒濕培養箱(25 ℃、相對濕度50%)平衡48 h。以相同方法制備不含復合保鮮液的NCC-SA膜。
1.3.2 樣品預處理
將新鮮羅非魚宰殺,去除頭、鱗、內臟(“三去”),切片后洗凈,置于紫外超凈工作臺上,用無菌水洗滌,并用濾紙瀝干。稱取40.0 g魚肉裝于70 mm×40 mm稱量皿中,用復合膜密封皿口,4 ℃冰箱貯藏8 d,每隔2 d測定魚肉各項生理指標的變化。試驗設3個處理,分別為對照(CK)組(無密封,敞口保存)、SA-NCC膜處理組以及GT-SA-NCC膜處理組。
1.3.3 魚肉持水力的變化
魚肉持水力根據貯藏期間質量差按公式(1)計算[3]:
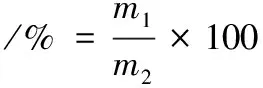
(1)
式中:m1為貯藏后魚肉重量,g;m2為新鮮魚肉重量,g。
1.3.4 魚肉pH值的測定
魚肉pH值測定參考GB 5009.237—2016《食品pH值的測定》。準確稱取5 g魚肉絞碎后置于錐形瓶中,加入45 mL蒸餾水,靜置30 min后測定pH值。
1.3.5 魚肉TBA值的測定
TBA值的測定參考竇川林[3]的方法。準確稱取10 g魚肉樣品,絞碎,加入25 mL 200 g/L三氯乙酸溶液,20 mL蒸餾水,10 000 r/min勻質60 s,4 ℃離心10 min(5 500 r/min)。吸取2.00 mL上清液于25 mL比色管中,加入2 mL TBA溶液(0.02 mol/L),沸水浴20 min,冷卻至室溫,532 nm和600 nm處測定吸光值。以2 mL三氯乙酸-水(體積比為1∶1)為空白。TBA值按公式(2)計算:
(2)
式中:A532為待測液在532 nm處的吸光值;A600為待測液在600 nm處的吸光值;155為丙二醛摩爾消光系數,L/(mol·cm);72.6為丙二醛相對分子質量;10為樣品質量,g。
1.3.6 魚肉揮發性鹽基氮(total volatile basic nitrogen,TVB-N)的測定
TVB-N測定參考GB 5009.228—2016《食品中揮發性鹽基氮的測定》。準確稱取10 g魚肉攪碎,加入100 mL 20 g/L三氯乙酸溶液(2%),搖勻并靜置15 min,8 000 r/min離心10 min,取上清液備用。測定時安裝好半微量凱式定氮裝置,清洗并做好密封性檢查。準確吸取10 mL樣品液注入反應室,以10 mL蒸餾水洗滌管壁,再注入5 mL 10 g/L氯化鎂混懸液,塞緊棒狀玻塞,玻杯內注入少量水以防漏氣。接收瓶內加入10 mL硼酸溶液(2%,體積分數)及3~5滴甲基紅-亞甲基藍混合指示劑,冷凝管下端插入液面下,夾緊螺旋夾,開始蒸餾,5 min后移動接收瓶,使冷凝管離開液面,繼續蒸餾1 min。少量水沖洗冷凝管下端,取下接收瓶,以0.01 mol/L鹽酸標準滴定液滴定至藍紫色。魚肉TVB-N含量按公式(3)計算:
(3)
式中:V1為測定樣液消耗鹽酸標準液的體積,mL;V2為空白消耗鹽酸標準液的體積,mL;c為鹽酸標準液的濃度,mol/L;m為試樣質量,g;14為與1 mL鹽酸標準滴定溶液相當的氮的含量,mg;10為樣品質量,g;100為三氯乙酸溶液體積,100 mL。
1.3.7 魚肉菌落總數的測定
魚肉菌落總數的測定參考GB 4789.2—2016《食品微生物學檢驗菌落總數測定》。稱取10 g魚肉絞放于滅菌燒杯中并加入90 mL滅菌生理鹽水,均質30 s,以10倍梯度依次稀釋,選擇3個適宜的稀釋度,用移液槍各取1 mL稀釋液分別涂布于已滅菌的計數瓊脂平板,36 ℃恒溫培養48 h,平板計數法測定菌落總數。以滅菌的生理鹽水做空白對照實驗。
1.3.8 魚肉肌原纖維蛋白的提取
取5 g魚肉絞碎樣品,加入20 mL Tris-HCl緩沖提取液(20 mmol/L,pH 7.2),高速均質2 min,4 ℃冷凍離心20 min(8 000 r/min),去上清液。沉淀與20 mL提取液混合,離心,重復2次。取沉淀加入15 mL含0.6 mol/L NaCl的Tris-HCl緩沖液(20 mmol/L,pH 7.2),4 ℃靜置提取60 min后,冷凍離心20 min(8 000 r/min),上清液即為魚肉纖維蛋白溶液[10-11]。
肌原纖維蛋白含量的測定:取1 mL肌原纖維蛋白溶液于10 mL比色管中,加入4 mL雙縮脲試劑,混勻,室溫靜置30 min,540 nm處測定吸光值。參照蛋白標準曲線(Y=0.048 5X+0.118 4,R2=0.999 3)計算其肌原纖維蛋白溶液濃度。
1.3.9 魚肉肌原纖維蛋白溶解度的測定
溶解度的測定參考藍蔚青等[12]的方法。準確稱取1 g肌原纖維蛋白,加入20 mL含0.6 mol/L NaCl的Na2HPO4緩沖液(50 mmol/L,pH 6.25),混勻使其充分溶解,測定蛋白溶液濃度,4 ℃冷凍離心15 min(5 000 r/min),取上清液測定蛋白溶液質量濃度。魚肉肌原纖維蛋白溶解度按公式(4)計算:
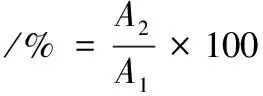
(4)
式中:A1為離心前魚肉蛋白溶液質量濃度,mg/mL;A2為離心后蛋白溶液質量濃度,mg/mL。
1.3.10 魚肉肌原纖維蛋白巰基含量的測定
巰基含量的測定參考WANG等[13]的方法,略作修改。取1 mL肌原纖維蛋白溶液(含量為5 mg/g)于25 mL比色管中,加入9 mL含0.6 mol/L KCl,10 mmol/L EDTA,8 mol/L尿素的磷酸鹽緩沖液(50 mmol/L,pH 7.0),旋渦振蕩搖勻,取5 mL于10 mL比色管中,加入0.5 mL DTNB溶液(準確稱取396 mg DTNB,用0.1 mol/L,pH 7.0的磷酸鹽緩沖液定容至100 mL),25 ℃靜置25 min,412 nm處測定吸光值。魚肉肌原纖維蛋白巰基含量按公式(5)計算:
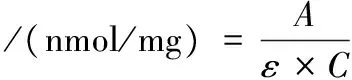
(5)
式中:A為吸光值;ε為巰基濃度的摩爾吸光系數,13 600 L/(mol·cm);C為肌原纖維蛋白質量濃度,5 mg/mL。
1.3.11 魚肉肌原纖維蛋白羰基含量的測定
羰基含量的測定參考張慧蕓等[14]的方法,略作修改。取1 mL肌原纖維蛋白溶液(質量濃度為2 mg/mL)于10 mL比色管中,加入1 mL DNPH溶液(10 mmol/L),旋渦振蕩搖勻,室溫避光靜置60 min,加入1 mL 200 g/L三氯乙酸溶液,12 000 r/min冷凍離心3 min,去上清液,沉淀用1 mL等體積比混合的乙酸乙酯-乙醇溶液洗滌3次,加入3 mL鹽酸胍溶液(6 mol/L),37 ℃水浴15 min使沉淀溶解,12 000 r/min離心3 min,取上清液370 nm處測定吸光值。魚肉肌原纖維蛋白羰基含量按公式(6)計算:
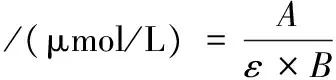
(6)
式中:A為吸光值;ε為羰基濃度的摩爾吸光系數,22 000 L/(mol·cm);B為吸收池光程長度,10 mm。
1.3.12 魚肉肌原纖維蛋白表面疏水性的測定
表面疏水性的測定參考張雪春等[15]的方法,略作修改。取1 mL肌原纖維蛋白樣液于25 mL比色管中,加入200 μL溴酚藍溶液(1 mg/mL),旋渦振蕩搖勻,8 000 r/min離心10 min,取上清液稀釋10倍,595 nm處測定吸光值A1,磷酸鹽緩沖液代替樣液測定吸光值A2。魚肉肌原纖維蛋白表面疏水性按公式(7)計算:
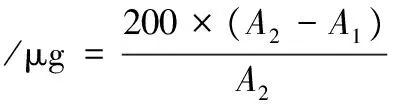
(7)
1.4 數據處理
所有試驗均設3組平行,利用軟件IBM SPSS Statistics 26進行數據分析,軟件GraphPad Prism 9.3.1進行繪圖。
2 結果與分析
2.1 貯藏過程中魚肉的品質特性
2.1.1 貯藏過程中魚肉持水力的變化
貯藏時間內CK組、SA-NCC組、GT-SA-NCC組魚肉持水率的變化如圖1所示。隨著貯藏時間的延長,魚肉持水率均持續降低。主要是因為隨著時間的延長,魚肉體內微生物的生長繁殖及復雜的生化反應,導致魚肉蛋白氧化分解,肌肉組織結構破壞,魚肉彈性、持水力降低[16]。與CK組相比,SA-NCC組、GT-SA-NCC組持水力的下降趨勢較慢。貯藏初期(0 d),CK組、SA-NCC組、GT-SA-NCC組持水率均為100%,貯藏第8天分別降至66.19%、80.70%、85.18%。原因為膜具有一定阻隔性能,可有效減少水蒸氣、氧氣透過率。GT-SA-NCC組由于GSE、TP具有一定的抗氧化、抗菌能力,可延緩魚肉脂質和蛋白氧化分解的速度,延緩了持水率的下降。
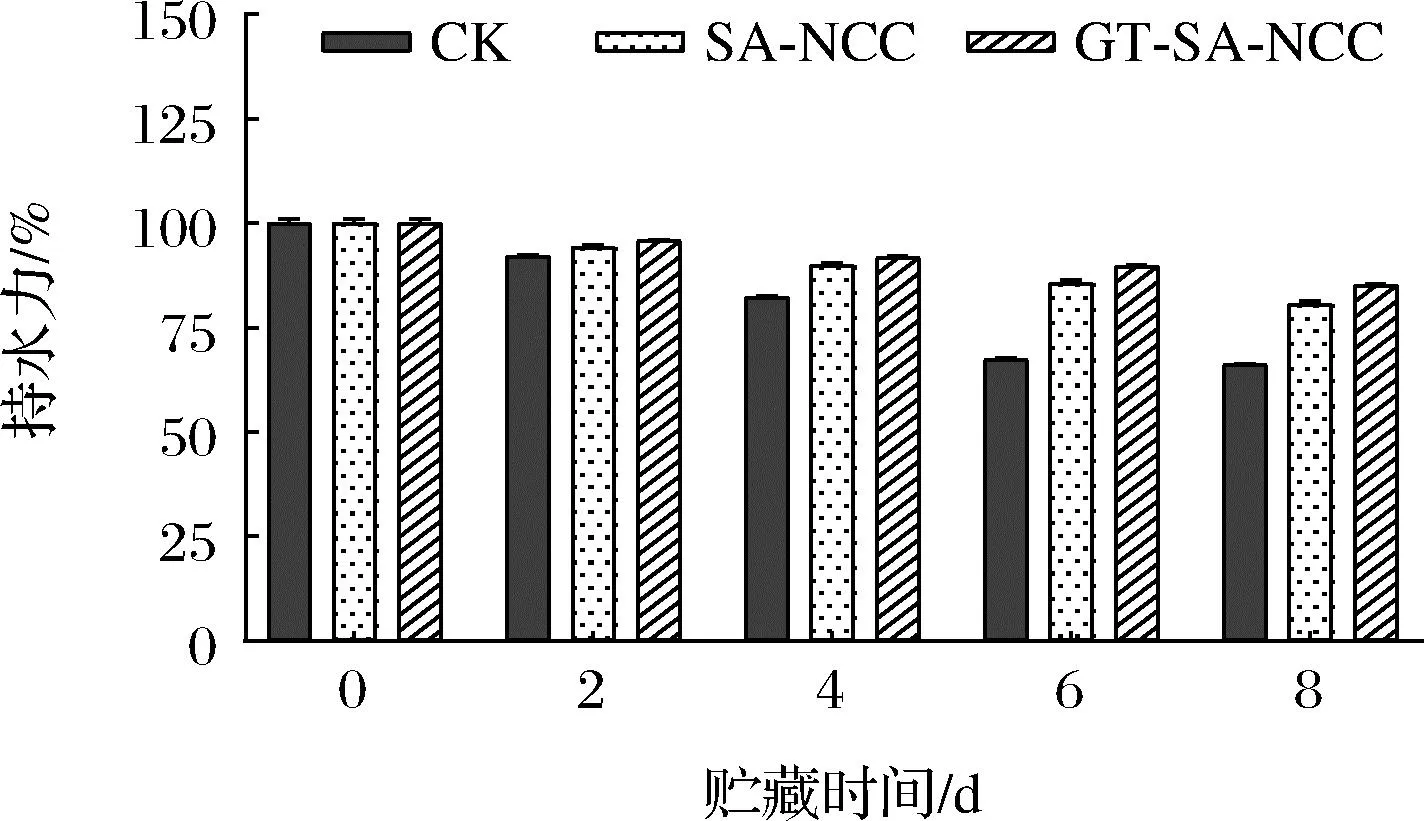
圖1 貯藏過程中魚肉持水力的變化
2.1.2 貯藏過程中魚肉pH值的變化
貯藏時間內CK組、SA-NCC組、GT-SA-NCC組魚肉pH值的變化如圖2所示。隨著貯藏時間的延長,魚肉pH值均呈先減小后增加的趨勢。貯藏初期(0~2 d),由于魚肉體內糖原無氧酵解產生乳酸,以及腺苷三磷酸(adenosine triphosphoric acid,ATP)經三羧酸循環產生的有機酸,導致pH值下降[17]。貯藏后期(2~8 d),蛋白質在內源酶的作用下分解為小分子肽和堿性氨基酸,后又因微生物作用分解為氨與三甲胺等堿性物質,導致pH上升,加速魚體腐敗[18]。與CK組相比,SA-NCC組pH值上升緩慢,貯藏8 d后pH值為6.69低于CK組(6.96)。原因為經膜覆蓋可有效降低氧氣與魚體的接觸,延緩魚體腐敗。GT-SA-NCC組緩慢下降第4天時pH達到最低,且上升速度最慢,貯藏8 d后pH值為6.58。原因為TP、GSE的加入可抑制糖原酵解中磷酸化酶的酶活,降低乳酸的生成。此外,TP、GSE具有抗菌、抗氧化作用,有利于抑制微生物生長繁殖,延緩魚體蛋白及脂質氧化,從而降低堿性物質的生成。
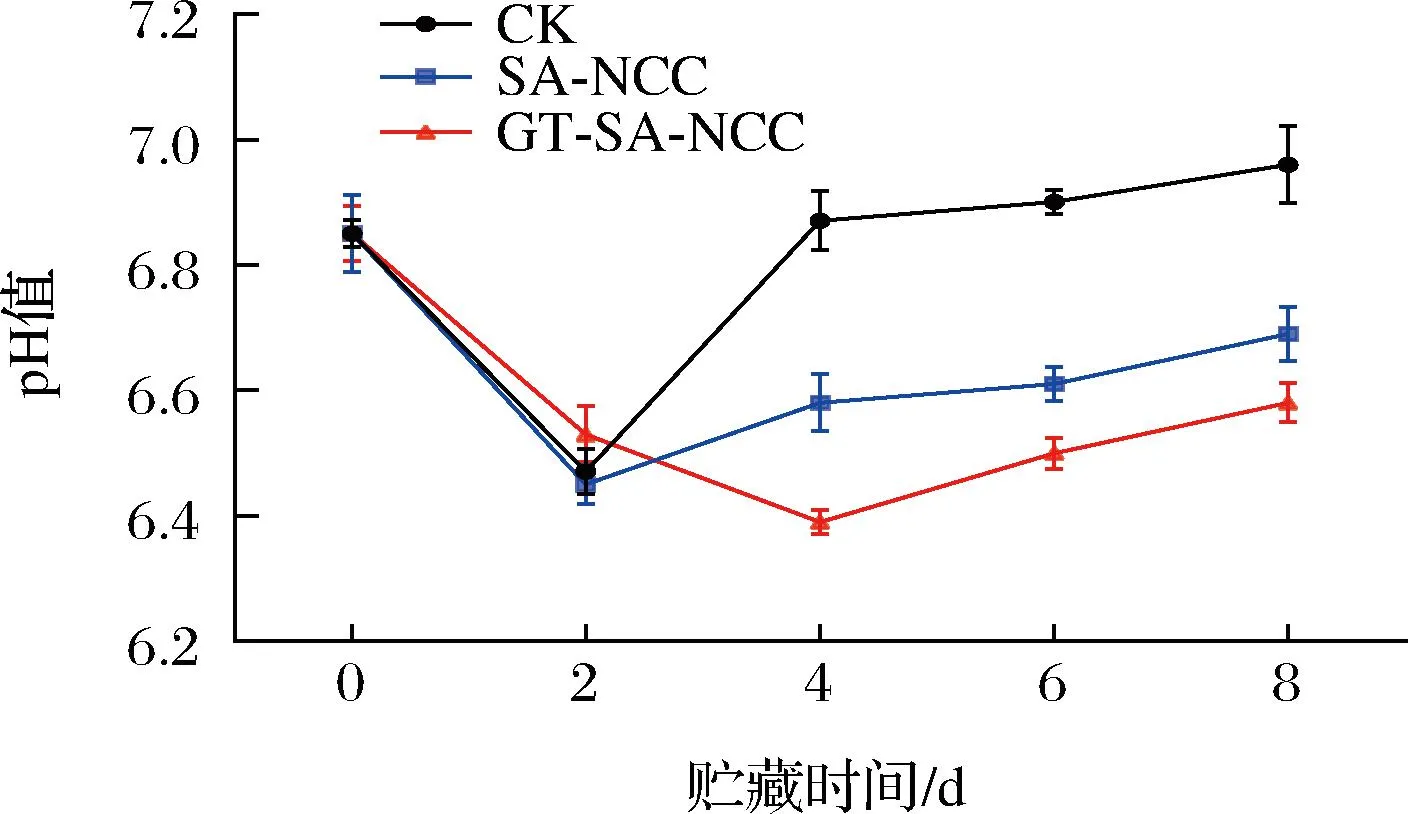
圖2 貯藏過程中魚肉pH值的變化
2.1.3 貯藏過程中魚肉TBA值的變化
魚肉在貯藏過程中由于微生物、酶、空氣等因素導致不飽和脂肪酸氧化,產生油脂氧化產物,并進一步降解為醛、酮等物質,導致魚肉酸敗、TBA值上升[19]。因此,TBA值是評價脂肪氧化程度的重要指標。貯藏過程中CK組、SA-NCC組、GT-SA-NCC組魚肉TBA值的變化如圖3所示。隨著貯藏時間的延長,所有魚肉處理組TBA值均持續上升。CK組魚肉的TBA上升最快,經膜覆蓋處理魚肉的TBA上升速度明顯減緩。新鮮魚肉的TBA值為2.38 mg/100 g,CK組貯藏8 d后TBA值上升至18.14 mg/100 g,此時魚體已經腐敗,不可食用。而其他處理組貯藏期間TBA值均低于CK組,GT-SA-NCC組始終保持最低(2.38~11.37 mg/100 g)。分析認為,GSE、TP加入使膜具有抗氧化和抗菌性能,能抑制魚體內脂肪的氧化,抑制魚肉變質腐敗,保證魚肉的優良品質,延長其貨架期。
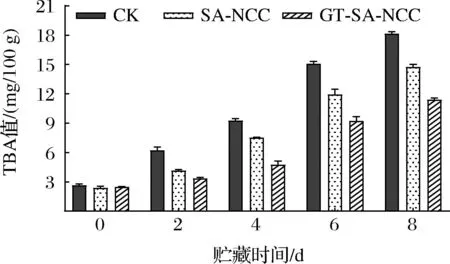
圖3 貯藏過程中魚肉TBA值的變化
2.1.4 貯藏過程中魚肉TVB-N值的變化
貯藏過程中,由于微生物及內源酶作用,魚體蛋白氧化分解為氨與胺類化合物,即TVB-N,其含量變化與新鮮度相關性較高,已被用于我國水產品新鮮度的鑒定標準[1]。GB/T 18108—2019《食品安全國家標準 鮮海水魚通則》指出,鮮海水魚肉TVB-N值≤15 mg/100 g時為優級品,合格品不超過30 mg/100 g。貯藏時間內CK組、SA-NCC組、GT-SA-NCC組魚肉TVB-N值的變化如圖4所示。新鮮魚肉的TVB-N值為5.367 mg/100 g,為優級品。隨著貯藏時間的延長,所有處理組魚肉的TVB-N均呈上升趨勢。貯藏第8天,CK組TVB-N值接近合格品限值,有明顯的胺臭味,說明魚體已腐敗變質失去食用價值。SA-NCC組、GT-SA-NCC組TVB-N值仍維持在優級品限值內,GT-SA-NCC組TVB-N值上升最慢。主要是由于TP、GSE的加入使膜具有一定的抑菌和抗氧化作用,在一定時間內能有效抑制微生物的生長代謝及魚體蛋白的氧化分解,延緩魚體品質的劣變。
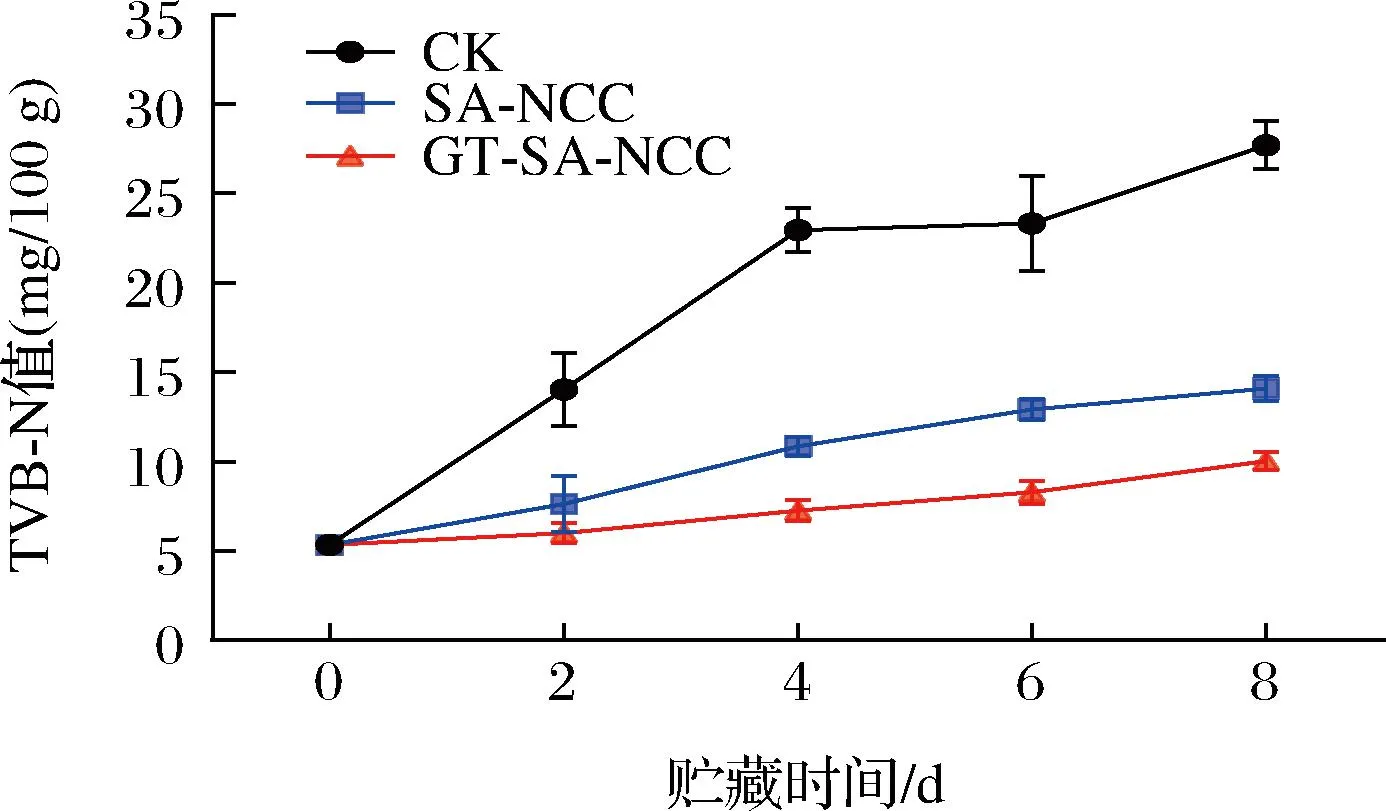
圖4 貯藏過程中魚肉TVB-N的變化
2.1.5 貯藏過程中魚肉菌落總數的變化
菌落總數是用來判定食品被細菌污染的程度及衛生質量的一個重要參數,它能夠反映食品在生產過程中是否符合衛生要求,以便對食品做出適當的衛生學評價。在水產品中菌落總數是判斷新鮮程度的一個重要指標。貯藏時間內CK組、SA-NCC組、GT-SA-NCC組魚肉菌落總數的變化如圖5所示,隨著貯藏時間的延長,所有處理組的鰱魚肉的菌落總數呈上升趨勢,且各組在0~4 d上升緩慢,4~8 d上升幅度較大。CK組、SA-NCC組在貯藏4 d時菌落總數分別達到7.46、6.79 lg CFU/g,均超過SC/T 3117—2006中規定菌落總數限值(4 lg CFU/g),魚肉已經失去食用價值。GT-SA-NCC組在貯藏6 d后菌落總數僅4.08 lg CFU/g。由菌落總數這一指標可判定,普通冷藏魚肉的貯藏期限為3~4 d,GT-SA-NCC膜包封處理后貯藏期限為5~6 d。說明GSE和TP在一定時間內能有效抑制微生物的生長繁殖,延緩魚體品質的劣變。對比其他理化指標,SA-NCC組菌落總數與CK組區別不大,這是因為SA-NCC膜具有一定阻隔作用,可一定程度延緩魚肉因環境造成的氧化變質,但本身無抗菌作用,對微生物的抑制作用較小。
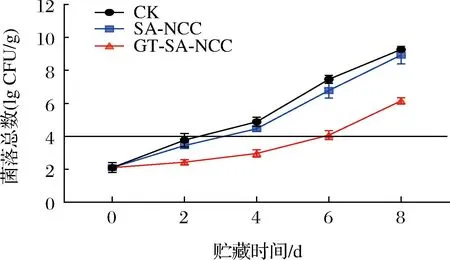
圖5 貯藏過程中魚肉菌落總數的變化
2.2 貯藏過程中魚肉肌原纖維蛋白的氧化變性
2.2.1 貯藏過程中魚肉肌原纖維蛋白溶解度的變化
魚肉蛋白質的功能性質主要由肌原纖維蛋白決定,其溶解度的變化可一定程度反映魚肉蛋白變性的情況。貯藏時間內CK組、SA-NCC組、GT-SA-NCC組肌原纖維蛋白溶解度的變化如圖6所示。隨著貯藏時間的延長,魚肉肌原纖維蛋白溶解度均持續下降。其原因為,貯藏過程中,魚肉肌原纖維蛋白氧化變性形成堿溶性大分子蛋白聚集體,這些大分子蛋白在高離子的微環境作用下溶解度降低。同時,由于蛋白氧化,巰基形成二硫鍵導致肌球蛋白重鏈聚合,降低其溶解度[20]。此外,在冷藏過程中,蛋白質與結合水間的結合狀態被破壞,蛋白質分子間作用增強,蛋白質與水間的相互作用減弱,導致蛋白質疏水基團暴露,疏水相互作使其產生凝聚和沉淀,導致溶解度降低。貯藏期間,SA-NCC組、GT-SA-NCC組溶解度均低于CK組,GT-SA-NCC組始終保持最低。貯藏8 d后,CK組、SA-NCC組、GT-SA-NCC組溶解度分別為40.54%、43.96%、49.31%,分別下降了25.59%、22.17%、16.82%。說明活性物質GSE、TP的加入,使膜本身具有一定的抗氧化和抑菌作用,對抑制魚體自身氧化及微生物的生長有較好的效果,從而延緩魚肉肌原纖維蛋白溶解度的降低。
2.2.2 貯藏過程中魚肉肌原纖維蛋白巰基含量的變化
巰基是魚肉蛋白質分子中反應活性較高的基團之一,蛋白質巰基上的硫外層具有較多的孤對電子,親核性很強,對氧化反應較為敏感。此外,巰基還與ATPase活性、肌球蛋白重鏈的氧化、二聚物的形成、以及蛋白質的聚合和變性關系密切。因此,巰基含量的變化是反映魚肉肌原纖維蛋白氧化變性程度的一個重要指標。貯藏時間內CK組、SA-NCC組、GT-SA-NCC組魚肉肌原纖維蛋白巰基含量的變化如圖7所示。隨著貯藏時間的延長,肌原纖維蛋白中巰基含量均呈不同程度的下降趨勢。這是因為貯藏期間,魚肉因體內微生物生長代謝及自溶酶作用,促使蛋白空間構象變化,隱藏在分子內部及分布在肌球蛋白頭部的巰基暴露并氧化形成二硫鍵[21]。貯藏期間,SA-NCC組、GT-SA-NCC組巰基含量均低于CK組,GT-SA-NCC組疏基含量始終保持最低。原因為經膜密封處理后,可一定程度隔絕氧氣、水蒸氣延緩魚體自身蛋白氧化。貯藏8 d后,CK組、SA-NCC組、GT-SA-NCC組巰基含量分別為5.70、8.09、11.35 nmol/mg,分別降低了15.33、11.95、15.33 nmol/mg。說明活性物質GSE、TP的加入,使膜本身具有一定的抗氧化和抑菌作用,同時隨著貯藏時間的延長,活性物質緩慢釋放,可一定程度抑制魚體自身氧化及微生物的生長,從而延緩魚體巰基氧化。
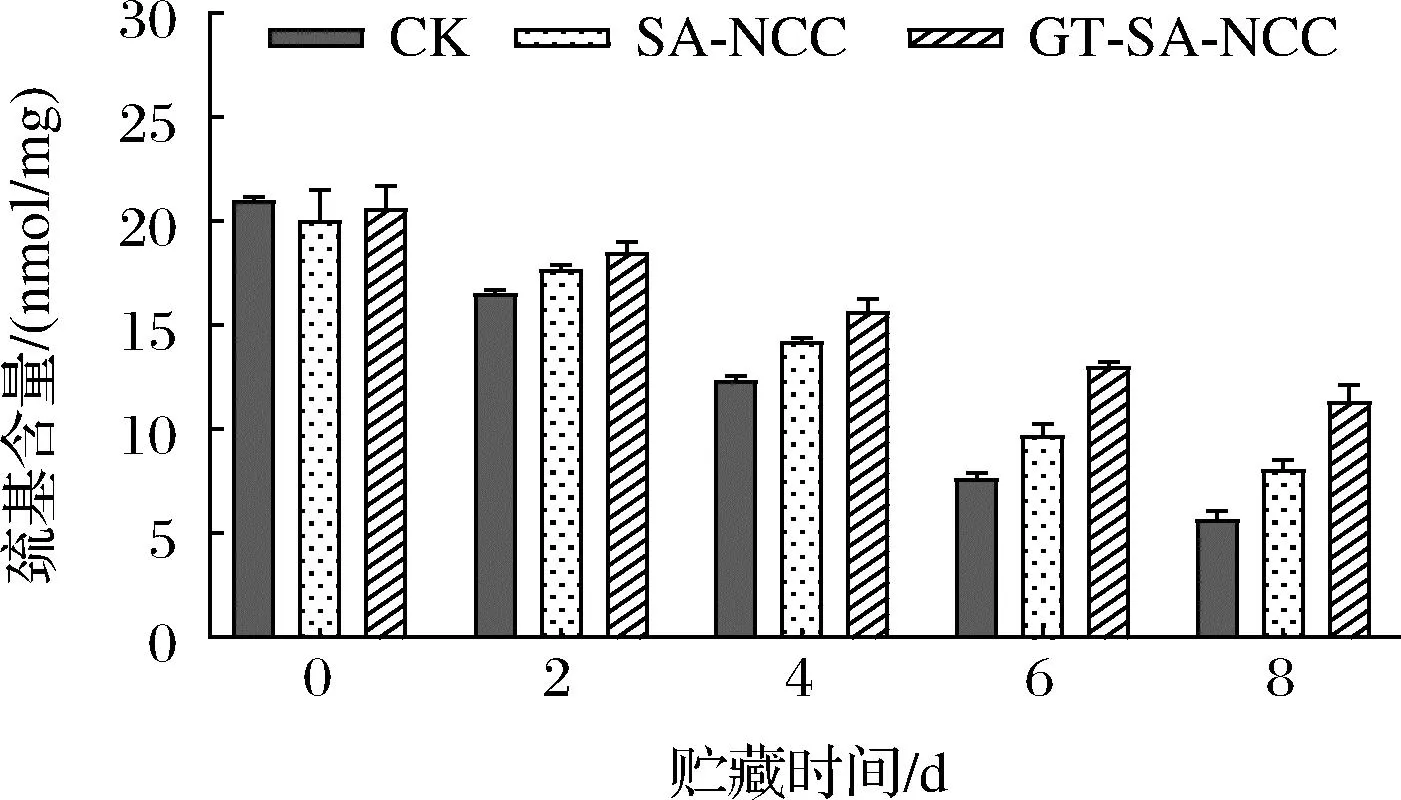
圖7 貯藏過程中魚肉肌原纖維蛋白巰基含量的變化
2.2.3 貯藏過程中魚肉肌原纖維蛋白羰基含量的變化
羰基化反應是蛋白質發生的一種不可逆非酶修飾,其中氧化應激和其他機制也能誘導羰基分子的形成。羰基含量變化也是用來反映蛋白氧化變性程度的重要指標之一[14]。貯藏時間內CK組、SA-NCC組、GT-SA-NCC組魚肉肌原纖維蛋白羰基含量的變化如圖8所示。隨著貯藏時間的延長,所有處理組肌原纖維蛋白中羰基含量均呈不同程度的上升趨勢。原因主要是隨著貯藏時間的延長,氨基酸側鏈直接氧化、氨基酸與次級氧化產物如丙二醛共價結合以及多肽骨架的氧化斷裂等多種脫氨反應途徑生成羰基化合物(醛和酮)[13]。貯藏期間,SA-NCC組、GT-SA-NCC組羰基含量均低于CK組,GT-SA-NCC組羰基含量始終保持最低。貯藏8 d后,CK組、SA-NCC組、GT-SA-NCC組羰基含量分別為29.39、26.40、23.86 μmol/L,分別增加了14.12、11.16、8.62 μmol/L。說明膜的阻隔性能可以有效延緩魚體自身蛋白氧化,同時活性物質GSE、TP的加入,使膜本身具有一定的抗氧化和抑菌作用,同時隨著貯藏時間的延長,活性物質緩慢釋放,可一定程度抑制魚體自身氧化及微生物的生長,從而延緩羰基化合物的生成。
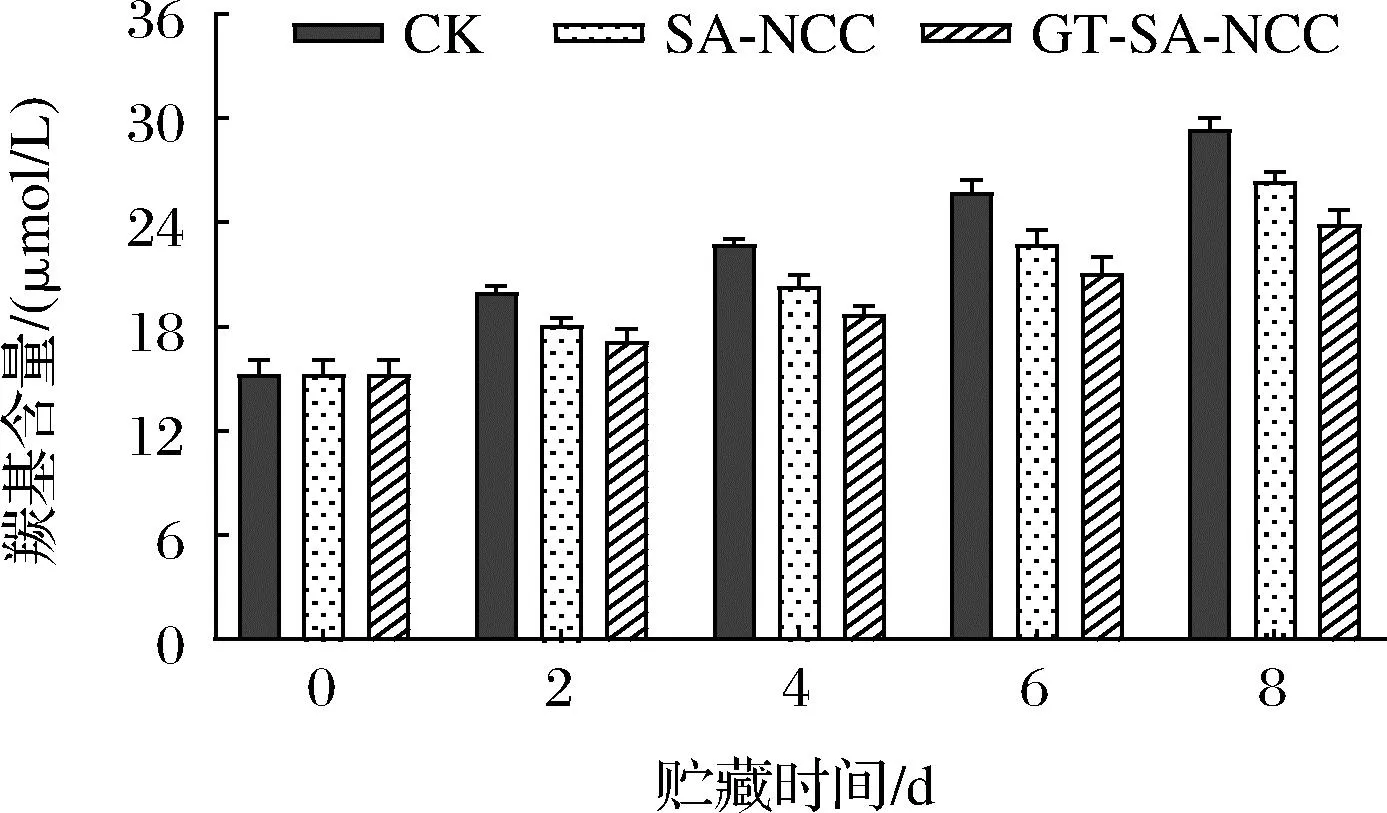
圖8 貯藏過程中魚肉肌原纖維蛋白羰基含量的變化
2.2.4 貯藏過程中魚肉肌原纖維蛋白表面疏水性的變化
表面疏水性是指蛋白質分子內部疏水性氨基酸聚集所形成的疏水力。研究指出,因蛋白質變性致使空間構象改變,原先隱藏于蛋白質分子內部的疏水性氨基酸暴露在分子表面,導致蛋白質表面疏水性變化[22]。因此,表面疏水性可用于表征蛋白的構象和氧化變性程度。貯藏時間內CK組、SA-NCC組、GT-SA-NCC組魚肉肌原纖維蛋白表面疏水性的變化如圖9所示。隨著貯藏時間的延長,所有處理組魚肉肌原表面疏水性均持續上升。這可能是由于在冷藏過程中,魚肉蛋白氧化引起分子內部空間構想改變,隱藏于蛋白質分子內部的疏水性氨基酸持續暴露引起。這與盧涵[23]研究的鳙魚肌原纖維蛋白表面疏水性變化趨勢一致。CK組上升最快,GT-SA-NCC組較為緩慢,且貯藏期間表面疏水性保持最低。貯藏8 d后,CK組、SA-NCC組、GT-SA-NCC組溴酚藍結合量分別為64.61、58.63、64.61 μg,分別增加了37.35、31.37、24.01 μg。說明GSE、TP的加入使膜有效抑制蛋白氧化變性,減少內部的疏水性氨基酸的暴露,延緩蛋白表面疏水性上升,延長了產品貨架期。
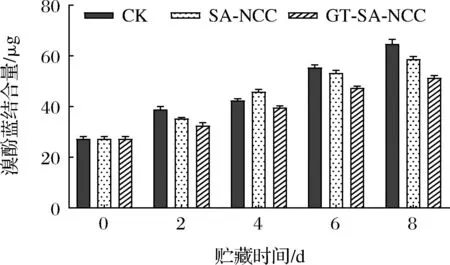
圖9 貯藏過程中魚肉肌原纖維蛋白表面疏水性的變化
3 結論
本試驗以海藻酸鈉(SA)、納米纖維(NCC)為基材,添加葡萄籽提取物(GSE)、茶多酚(TP)復合保鮮劑制備活性包裝膜用于羅非魚保鮮。結果表明,與CK、SA-NCC組相比,GT-SA-NCC處理有利于保持魚肉水分,抑制pH值、TBA值、TVB-N值升高,降低魚肉的菌落總數;同時,抑制魚肉肌原纖維蛋白地氧化變性、延緩溶解度、抑制巰基含量的降低以及羰基含量、表面疏水性的增加。說明添加GSE、TP所制備的活性包裝膜有利于延緩羅非魚脂質及蛋白氧化,對維持魚肉貯藏過程中的品質特性及蛋白的功能性質具有較理想的效果,在水產品及肉制品加工貯藏過程中具有一定的運用潛力。
