蘇紫豬血清細(xì)胞因子含量對育肥性狀的影響
2024-04-08 11:22:17付言峰程金花廖超戴超輝涂楓王學(xué)敏
江蘇農(nóng)業(yè)科學(xué) 2024年3期
付言峰 程金花 廖超 戴超輝 涂楓 王學(xué)敏

摘要:為研究育肥期蘇紫豬血清中細(xì)胞因子含量對育肥性狀的影響,本研究選取18頭蘇紫豬進(jìn)行生長育肥測定,然后屠宰采集血樣,進(jìn)行CCL4、IL-6和FGF-2這3 種細(xì)胞因子的檢測,同時(shí)統(tǒng)計(jì)其日齡、體質(zhì)量、瘦肉率和皮下脂肪,分析兩者之間的關(guān)系。結(jié)果發(fā)現(xiàn),CCL4、IL-6和FGF-2的標(biāo)準(zhǔn)曲線擬合度分別為0.996 3、0.995 8、0.995 3,均非常接近于1,說明后面檢測的結(jié)果可信度高。育肥期蘇紫豬血清中,IL-6含量最高,其次為CCL4含量,F(xiàn)GF-2 含量最低(P<0.05)。IL-6含量與日增質(zhì)量、瘦肉率呈負(fù)相關(guān)(相關(guān)系數(shù)分別為-0.41、-0.21),與皮下脂肪率呈正相關(guān)(相關(guān)系數(shù)為0.08);CCL4含量與瘦肉率呈負(fù)相關(guān)(相關(guān)系數(shù)為-0.44);FGF-2含量最高的豬日增質(zhì)量最快。研究提示,優(yōu)質(zhì)豬血清細(xì)胞因子含量對育肥性狀有較大的影響,其中,育肥期IL-6和CCL4含量可反映出豬瘦肉率的高低,有利于豬高瘦肉率性狀的高效選育。
關(guān)鍵詞:蘇紫豬;趨化因子C-C配體4;成纖維細(xì)胞生長因子2;成纖維細(xì)胞生長因子2;生長育肥
中圖分類號(hào):S828.6? 文獻(xiàn)標(biāo)志碼:A
文章編號(hào):1002-1302(2024)03-0201-06
我國是一個(gè)生豬養(yǎng)殖和消費(fèi)大國,《中國畜禽種業(yè)發(fā)展報(bào)告2022》數(shù)據(jù)顯示,2021年12月,全國能繁母豬存欄量為4 329萬頭,全國生豬存欄量為44 922萬頭,出欄量67 128萬頭,豬肉產(chǎn)量為 5 296萬t (肉類占比59.6%),人均豬肉占有量37.49 kg[1]。
蘇紫豬是江蘇省農(nóng)業(yè)科學(xué)院畜牧研究所豬育種與生產(chǎn)創(chuàng)新團(tuán)隊(duì)正在培育的快長節(jié)糧型優(yōu)質(zhì)豬新品種[2],含蘇鐘豬[3-4]、淮豬[5]和巴克夏豬[6]血統(tǒng)。蘇紫豬全身被毛黑色,生產(chǎn)性能優(yōu)異,除了含有人體生理活動(dòng)所需的優(yōu)質(zhì)蛋白質(zhì)和脂肪主成分以外,還富含維生素、肌苷酸、氨基酸和其他風(fēng)味物質(zhì),加工后肉質(zhì)鮮、香、嫩、口感好,在市場上深受消費(fèi)者的青睞[7]。所以,屠宰性狀的選育是其一個(gè)重要選育方向。
另外,在種豬選育中,生長性狀(料質(zhì)量比、日增質(zhì)量)也是當(dāng)前非常重要的育種性狀,此性狀屬于中等遺傳力性狀[8]。及時(shí)篩選到具有優(yōu)良生長性狀的種豬,能快速提高整個(gè)生豬體系的生產(chǎn)效率,實(shí)現(xiàn)更高的生豬養(yǎng)殖效益[9]。隨著人民生活水平的提高,消費(fèi)者對肉質(zhì)的需求也要不斷增加[10]。因此,有必要針對優(yōu)質(zhì)豬種豬[11]開展上述生長育肥性狀的相關(guān)研究,為豬場的引種、生豬改良工作提供理論依據(jù)。
趨化因子C-C配體4(C-C motif chemokine 4,CCL4)屬于炎癥性CC 類趨化因子亞家族的成員,又稱巨噬細(xì)胞炎癥蛋白1β(macrophage inflammatory protein-1β,MIP-1β)[12],是多種類型細(xì)胞,包括巨噬細(xì)胞、自然殺傷細(xì)胞、單核細(xì)胞、未成熟樹突狀細(xì)胞和冠狀動(dòng)脈內(nèi)皮細(xì)胞等的趨化劑[13]。分子質(zhì)量一般為8~12 ku,為細(xì)胞因子蛋白樣物質(zhì),是體內(nèi)促炎趨化因子,參與細(xì)胞分化、遷移和組織定位等過程[14]。對于CCL4的作用,在人類疾病中的研究較多[15]。大鱗副泥鰍(Paramisgurnus dabryanus) CCL4在上頜觸須發(fā)育的晚期高度表達(dá),表明CCL4可能作為調(diào)節(jié)基因參與上頜觸須的發(fā)育[16]。
豬白細(xì)胞介素6(IL-6)在1980年被發(fā)現(xiàn),主要由活化的T細(xì)胞、成纖維細(xì)胞、脂肪細(xì)胞分泌和合成,是一種功能廣泛的、具有多種生物學(xué)效應(yīng)的炎癥因子[17]。肥胖(體質(zhì)量過高)會(huì)導(dǎo)致炎癥因子TNF-α、IL-6表達(dá)水平上升[18],血液中大約1/3的循環(huán)IL-6來自脂肪組織,IL-6水平升高與肥胖,葡萄糖耐受不良,胰島素抵抗和T2DM呈正相關(guān)[19]。人孕期外周血IL-6水平與子代出生體質(zhì)量無明顯相關(guān),原因可能是研究對象經(jīng)濟(jì)水平較好,炎性因子平均水平較低所致[20]。IL-6基因的遺傳變異與民豬、長白豬哺乳仔豬腹瀉存在不同程度的相關(guān)[21]。
成纖維細(xì)胞生長因子2(fibroblast growth factor 2,F(xiàn)GF-2)由多種細(xì)胞類型產(chǎn)生,是一種重要的促血管生成因子[22]和肌肉因子[23],并通過其受體與各種硫酸乙酰肝素結(jié)合和組成調(diào)節(jié)其活性的蛋白聚糖[24]。FGF-2可引起內(nèi)皮細(xì)胞的遷移和生長,又能夠促進(jìn)腫瘤血管生成,參與豬胚胎附植過程[25],還能夠通過NDY1/KDM2B-miR-101-EZH2途徑來調(diào)控細(xì)胞增殖等過程[26]。肥胖小鼠皮下脂肪組織中FGF-2轉(zhuǎn)錄水平表達(dá)量升高,且FGF-2劑量依賴性抑制早期脂肪細(xì)胞分化[27]。
1 材料與方法
1.1 試驗(yàn)動(dòng)物
試驗(yàn)地點(diǎn)在江蘇省農(nóng)業(yè)科學(xué)院合作單位(江蘇融港良種繁育有限公司)豬場,試驗(yàn)動(dòng)物包括:蘇紫豬35頭。按性別、血統(tǒng)分成5欄飼養(yǎng),飼養(yǎng)在種豬測定站上。試驗(yàn)開始日期:2022年7月6日,結(jié)束日期:2023年12月31日。2022年11月14日,對18頭豬進(jìn)行屠宰測定,采血后分析血清細(xì)胞因子含量。
1.2 試劑儀器
主要試劑為3種試劑盒,分別為:MM-0418O1 豬白細(xì)胞介素6(IL-6)ELISA試劑盒 96T、MM-77683O1 豬趨化因子C-C-基元配體4(CCL4)ELISA試劑盒 96T、MM-78107O1豬成纖維細(xì)胞生長因子2(FGF-2)ELISA試劑盒96T,均為酶免(MEIMIAN)品牌,來自江蘇酶免實(shí)業(yè)有限公司(Jiangsu Meimian industrial Co.,Ltd)。
主要儀器包括:酶標(biāo)儀(芬蘭,Labsystems Multiskan MS,352型)、洗板機(jī)(芬蘭,Thermo Labsystems,AC8型)、微量高速離心機(jī)(中國,湘儀,TG16W型)、隔水式恒溫培養(yǎng)箱(中國,江東,GNP-9080型)和全自動(dòng)生化分析儀(美國,貝克曼庫爾特,AU480型)。
1.3 細(xì)胞因子測定方法
使用ELISA方法進(jìn)行測定。首先進(jìn)行標(biāo)準(zhǔn)品的稀釋,1、2、3、4、5號(hào)標(biāo)準(zhǔn)品的濃度分別稀釋至50、100、200、400、800 ng/L,即取150 μL各號(hào)標(biāo)準(zhǔn)品,加入150 μL標(biāo)準(zhǔn)品稀釋液。其次進(jìn)行以下操作:(1)加樣,分別設(shè)空白孔(空白對照孔不加樣品及酶標(biāo)試劑,其余各步操作相同)、標(biāo)準(zhǔn)孔、待測樣品孔。在酶標(biāo)包被板上標(biāo)準(zhǔn)品準(zhǔn)確加樣50 μL,待測樣品孔中先加樣品稀釋液40 μL,然后再加待測樣品 10 μL(樣品最終稀釋度為5倍)。將樣品加于酶標(biāo)板孔底部,盡量不觸及孔壁,輕輕晃動(dòng)混勻。(2)溫育:用封板膜封板后置于37 ℃溫育30 min。(3)配液:將30 倍濃縮洗滌液用蒸餾水30倍稀釋后備用。(4)洗滌:小心揭掉封板膜,棄去液體,甩干,每孔加滿洗滌液,靜置30 s后棄去,如此重復(fù)5 次,拍干。(5)加酶:每孔加入酶標(biāo)試劑50 μL,空白孔除外。(6)再重復(fù)進(jìn)行1次溫育和洗滌。(7)顯色:每孔先加入顯色劑A 50 μL,再加入顯色劑B 50 μL,輕輕振蕩混勻,37 ℃避光顯色10 min。(8)終止:每孔加終止液 50 μL,終止反應(yīng)(此時(shí)藍(lán)色立轉(zhuǎn)黃色)。最后進(jìn)行測定,以空白孔調(diào)零,450 nm 波長依序測量各孔的吸光度(D值),測定應(yīng)在加終止液后15 min以內(nèi)進(jìn)行。
1.4 豬育肥性狀測定方法
采用種豬自動(dòng)測定站進(jìn)行測定育肥性狀。先將試驗(yàn)豬打上電子耳豬,統(tǒng)一趕入測定舍,分成5欄飼喂,每個(gè)飼養(yǎng)欄內(nèi)設(shè)置1臺(tái)測定站,每天利用測定站自動(dòng)記錄和上傳蘇紫豬在測定日齡內(nèi)的采食量數(shù)據(jù)、增質(zhì)量數(shù)據(jù)、生產(chǎn)性能數(shù)據(jù)。其中,每一條采食量數(shù)據(jù)包括:欄位、測定站號(hào)、電子耳牌號(hào)、開始日期、結(jié)束日期、測定天數(shù)、采食量、采食次數(shù)、采食時(shí)間和平均日采食量;每一條增重?cái)?shù)據(jù)包括:欄位、測定站號(hào)、電子耳牌號(hào)、開始日期、結(jié)束日期、開始體質(zhì)量、結(jié)束體質(zhì)量、增質(zhì)量、測定天數(shù)、平均日增質(zhì)量、每個(gè)測定日體質(zhì)量和日增質(zhì)量;每一條生產(chǎn)性能數(shù)據(jù)包括:欄位、測定站號(hào)、電子耳牌號(hào)、采食量、起始體質(zhì)量、增質(zhì)量、平均日采食量、平均日增質(zhì)量和飼料報(bào)酬(料質(zhì)量比)。
瘦肉率和皮下脂肪率是在采血當(dāng)天對試驗(yàn)豬進(jìn)行屠宰后現(xiàn)場測定,具體為左半胴體分割成前、中、后3段,每段進(jìn)行皮、皮下脂肪、瘦肉和骨頭4個(gè)部分的手工剝離,依次稱質(zhì)量記錄,4個(gè)部分相加為100%(不計(jì)算分割過程中的損耗,不包括板油、腎),最后分別采用瘦肉和皮下脂肪除以4個(gè)部分總質(zhì)量,獲得各部分所占的比例。
1.5 數(shù)據(jù)統(tǒng)計(jì)
數(shù)量儲(chǔ)存、篩選、分類匯總等整理是通過Excel和Access軟件分析。不同組別間測定數(shù)值的差異性顯著水平通過SSPS 9.1統(tǒng)計(jì)軟件包進(jìn)行GLM分析,結(jié)果以“最小二乘均值±標(biāo)準(zhǔn)誤”表示。
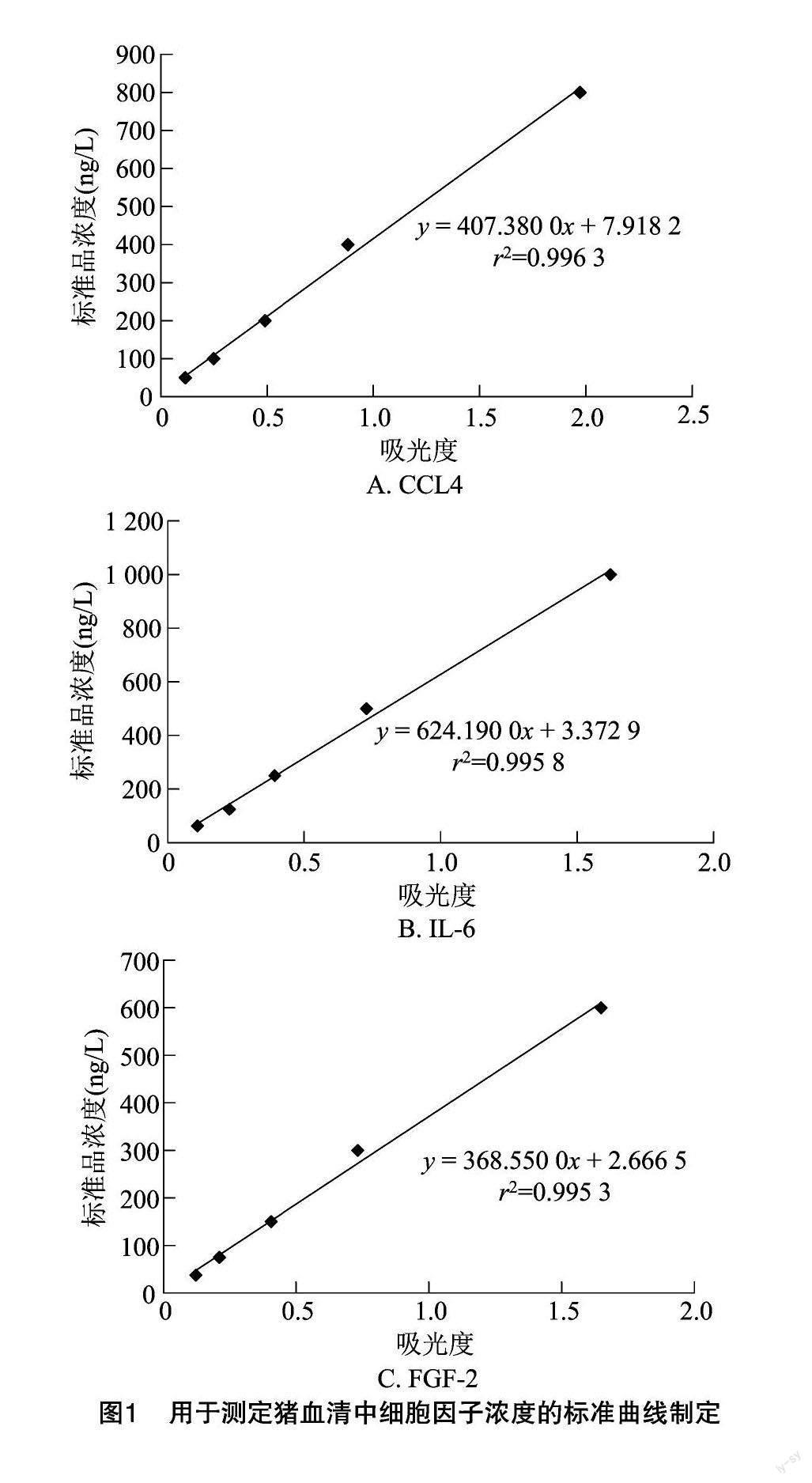
細(xì)胞因子含量是通過標(biāo)準(zhǔn)曲線方法計(jì)算出來的,即用各自的5種標(biāo)準(zhǔn)物(濃度已知)和空白對照,采用酶標(biāo)儀測定出各標(biāo)準(zhǔn)物的吸光度(D值)。然后以D值為橫坐標(biāo),以標(biāo)準(zhǔn)物濃度為縱坐標(biāo),繪制出各細(xì)胞因子的標(biāo)準(zhǔn)曲線。最后,待測樣品根據(jù)測定的D值,在標(biāo)準(zhǔn)曲線上找到對應(yīng)的濃度。
2 結(jié)果與分析
2.1 標(biāo)準(zhǔn)曲線制定結(jié)果
3種細(xì)胞因子CCL4、IL-6和FGF-2,分別對應(yīng)3條標(biāo)準(zhǔn)曲線,同時(shí)獲得了標(biāo)準(zhǔn)曲線的線性擬合回歸方程式和線性擬合度(r2)。結(jié)果表明,CCL4的標(biāo)準(zhǔn)曲線擬合的回歸方程為y=407.380 0x+7.918 2,擬合度為r2=0.996 3。IL-6標(biāo)準(zhǔn)曲線擬合的回歸方程為y=624.190 0x+3.372 9,擬合度為 r2=0.995 8,斜率(624.19)>CCL4(407.38),擬合度略低于CCL4。FGF-2標(biāo)準(zhǔn)曲線擬合的回歸方程為 y=368.550 0x+2.666 5,擬合度為r2=0.995 3,斜率(368.55)最低,小于CCL4和IL-6,擬合度也最低,小于CCL4和IL-6。由圖1可知,3條標(biāo)準(zhǔn)曲線擬合度均非常接近于1,說明擬合效果均非常理想,故根據(jù)各自線性擬合回歸方程式計(jì)算出的待測樣品濃度準(zhǔn)確性均非常高。
2.2 細(xì)胞因子測定結(jié)果
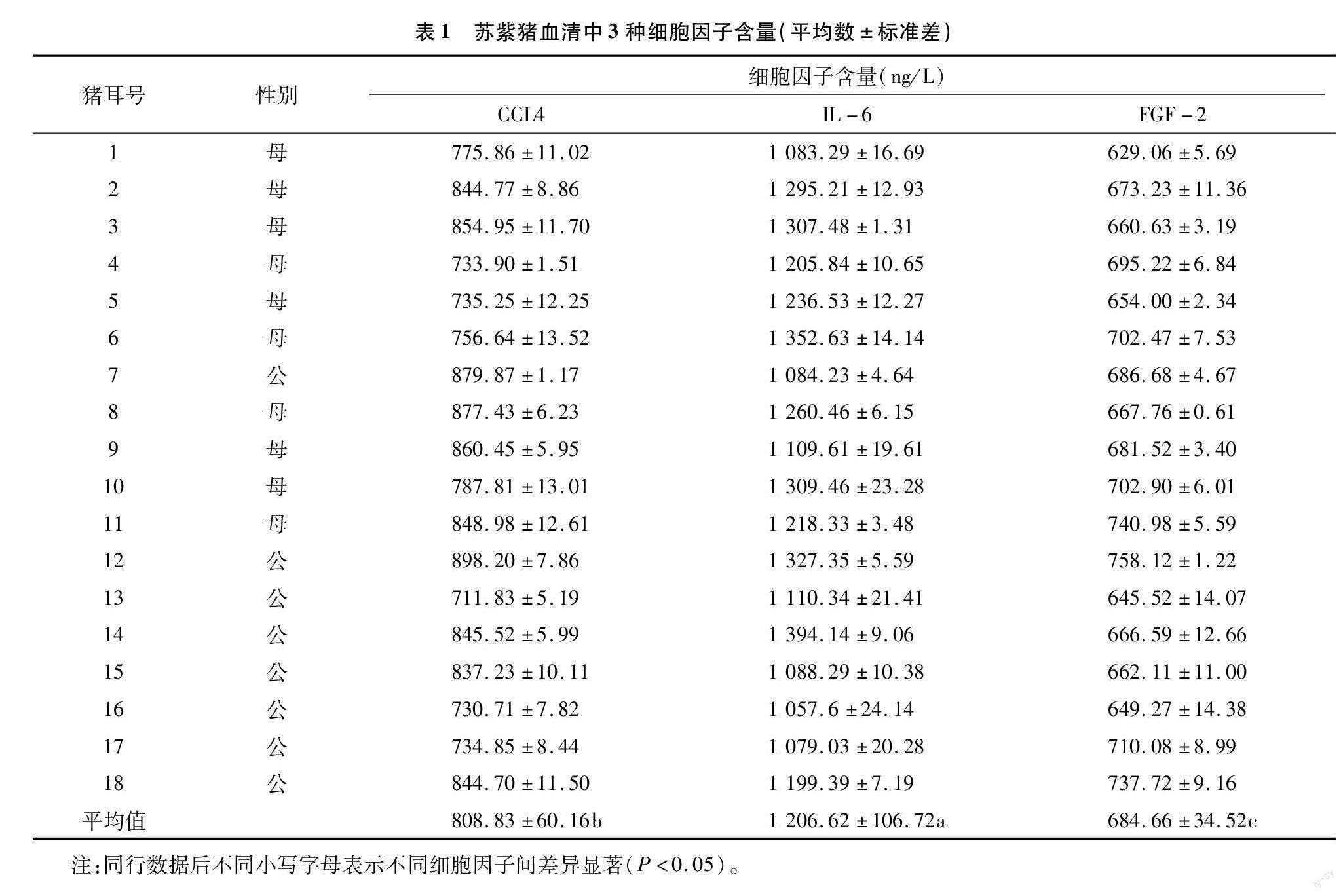
由表1可知,細(xì)胞因子的檢測有3次重復(fù),取其平均值。每頭豬根據(jù)上述標(biāo)準(zhǔn)曲線計(jì)算出的試驗(yàn)豬同一時(shí)間點(diǎn)的CCL4、IL-6和FGF-2平均含量分別為808.83、1 206.62、684.66 ng/L,且3種細(xì)胞因子的平均含量兩兩差異顯著(P<0.05),其中,IL-6含量最高,其次為CCL4,最低的為FGF-2。3種細(xì)胞因子含量最高值均出現(xiàn)在不同的豬上,最低值也均出現(xiàn)在不同的豬上。其中,CCL4最高值為898.2 ng/L,最低值為711.83 ng/L;IL-6最高值為 1 394.14 ng/L 最低值為1 057.6 ng/L;FGF-2最高值為737.72 ng/L,最低值為629.06 ng/L。
2.3 不同育肥性狀間的細(xì)胞因子差異性分析
對試驗(yàn)豬不同性別、體質(zhì)量、日齡間細(xì)胞因子進(jìn)行差異性比較,由表2可知:(1)性別間差異,IL-6 含量母豬顯著高于公豬(P<0.05),CCL4和FGF-2含量公母豬間無顯著差異。(2)體質(zhì)量間差異,CCL4含量在90 kg豬中顯著高于80、100 kg豬中(P<0.05),80、100 kg豬間差異不顯著;IL-6含量在90 kg豬中最高,其次為80 kg豬和100 kg豬,其中,90、100 kg豬間差異顯著(P<0.05)。(3)日齡間差異,CCL4含量在220 d豬中顯著高于190、200、210 d豬中(P<0.05),其中190 d豬中的CCL4含量最低;IL-6含量在220 d豬中顯著低于190、200 d豬中,其中,190 d豬中的IL-6含量最高。(4)
日增質(zhì)量間差異,IL-6含量在500 g日增質(zhì)量豬中顯著高于700 g豬中(P<0.05);FGF-2在800 g日增質(zhì)量豬中顯著高于500、600、700 g日增質(zhì)量豬中(P<0.05),其他組間差異不顯著。(5)瘦肉率間差異,CCL4在50%和55%瘦肉率豬中的含量顯著高于60%和65%的豬中(P<0.05);IL-6在50%瘦肉率豬中的含量最高,在65%瘦肉率豬中的含量最低,且這個(gè)最低值顯著低于50%、60%瘦肉率的豬(P<0.05);FGF-2在50%瘦肉率豬中的含量最高,在55%瘦肉率豬中的含量最低,且這個(gè)最低值顯著低于50%瘦肉率的豬(P<0.05)。(6)皮下脂肪率間差異,只有IL-6含量在不同皮下脂肪率間有顯著差異,IL-6在30%皮下脂肪率豬中的含量最高,在15%和25%皮下脂肪率豬中的含量最低,且二者間差異顯著(P<0.05)。
2.4 育肥性狀和細(xì)胞因子關(guān)聯(lián)性分析
對豬細(xì)胞因子含量和育肥性狀進(jìn)行關(guān)聯(lián)性分析,結(jié)果(表3)表明,3種細(xì)胞因子含量間均呈正相關(guān),相關(guān)系數(shù)分別為0.30、0.36、0.31;6個(gè)育肥性狀間有8個(gè)正相關(guān)(最高值0.62)、6個(gè)負(fù)相關(guān)(絕對值最高值-0.69)、1個(gè)不相關(guān)(0)(不含自身);3種細(xì)胞因子含量與6個(gè)育肥性狀間有9個(gè)正相關(guān)(最高值0.33)、9個(gè)負(fù)相關(guān)(絕對值最高值-0.44)。其中,細(xì)胞因子含量與育肥性狀間的相關(guān)系數(shù)中,CCL4與瘦肉率呈負(fù)相關(guān),且相關(guān)系數(shù)的絕對值最高(-0.44),與日增質(zhì)量、皮下脂肪率均呈正相關(guān);IL-6 與日增質(zhì)量呈負(fù)相關(guān),且相關(guān)系數(shù)的絕對值最高(-0.41),與瘦肉率也呈負(fù)相關(guān),與皮下脂肪率呈正相關(guān);FGF-2與日增質(zhì)量、瘦肉率、皮下脂肪率均呈正相關(guān),但相關(guān)系數(shù)均不高(≤0.1)。
3 討論與結(jié)論
本研究中,3種細(xì)胞因子為趨化因子C-C-基元配體4(CCL4)[12]、豬白細(xì)胞介素6(IL-6)[21]和豬成纖維細(xì)胞生長因子2(FGF-2)[26],豬育肥性狀主要包含日增質(zhì)量、瘦肉率和皮下脂肪率[28]。研究結(jié)果表明,同一批豬屠宰采血后,測定的3種細(xì)胞因子含量對比,IL-6顯著高于CCL4,CCL4又顯著高于FGF-2。IL-6含量最高可能是分泌IL-6的細(xì)胞較多且進(jìn)行血液循環(huán)的也較多有關(guān),IL-6由活化的 T 細(xì)胞、成纖維細(xì)胞、脂肪細(xì)胞分泌和合成[17];FGF-2含量最低可能與分泌的細(xì)胞較少有關(guān),F(xiàn)GF-2 主要由血管內(nèi)的內(nèi)皮細(xì)胞、平滑肌細(xì)胞、巨噬細(xì)胞分泌,且主要作用是促血管生成[29]。
細(xì)胞因子含量對豬育肥性狀中的生長速度(日增質(zhì)量)有一定影響,本研究發(fā)現(xiàn),IL-6含量最高的豬,日增質(zhì)量最慢,且隨著IL-6含量降低,日增質(zhì)量變快,提示IL-6含量與日增質(zhì)量呈負(fù)相關(guān),原因可能是炎癥因子IL-6含量高的豬,會(huì)需要更高的能量用于保持健康[19],導(dǎo)致生長速度變慢。FGF-2 含量最高的豬中,生長速度最快,原因可能是生長速度快的豬,同樣時(shí)間內(nèi)會(huì)產(chǎn)生更多的肌肉,故肌肉因子FGF-2含量也較高[23]。
細(xì)胞因子含量對豬育肥性狀中的瘦肉率也有一定影響,表現(xiàn)為CCL4、IL-6和FGF-2 3 種細(xì)胞因子含量最高的豬中,瘦肉率最低;反之,CCL4和IL-6含量最低的豬中,瘦肉率最高,說明CCL4和IL-6含量與瘦肉率呈負(fù)相關(guān)。CCL4能在促炎因子的刺激下通過吸引白細(xì)胞等免疫細(xì)胞定向趨化到炎癥部位發(fā)揮作用[30],且CCL4注射配合高脂日糧的飼喂能加速小鼠的肥胖和脂肪肝形成[31],說明CCL4能促進(jìn)脂肪形成,降低瘦肉率,與本研究結(jié)果一致。IL-6與瘦肉率呈負(fù)相關(guān),可能與其功能有關(guān),有研究表明,肥胖會(huì)導(dǎo)致炎癥因子TNF-α、IL-6表達(dá)水平上升[18],脂肪增加后,瘦肉和瘦肉率就會(huì)相應(yīng)下降。
細(xì)胞因子含量對豬育肥性狀中的皮下脂肪率影響如下:皮下脂肪率和瘦肉率在一定程度上是相反的;所以,IL-6含量最高的豬,皮下脂肪率也最高;反之,IL-6含量最低的豬,皮下脂肪率也最低,說明IL-6細(xì)胞因子含量與皮下脂肪率呈正相關(guān)。研究表明,人血液中大約1/3的循環(huán) IL-6 來自脂肪組織,IL-6 含量升高與肥胖呈正相關(guān)[19],這與本研究結(jié)果類似。
綜上所述,育肥期蘇紫豬血清中,IL-6含量最高,其次為CCL4含量,F(xiàn)GF-2含量最低(P<0.05)。IL-6含量與生長速度(日增質(zhì)量)、瘦肉率呈負(fù)相關(guān),與皮下脂肪率呈正相關(guān);CCL4含量與瘦肉率呈負(fù)相關(guān);FGF-2含量最高的豬生長速度最快。
參考文獻(xiàn):
[1]農(nóng)業(yè)農(nóng)村部種業(yè)管理司,全國畜牧總站. 中國畜禽種業(yè)發(fā)展報(bào)告2022[R]. 北京:中國農(nóng)業(yè)科學(xué)技術(shù)出版社,2022.
[2]王學(xué)敏,涂 楓,任守文,等. 蘇紫黑豬1~5世代繁殖與生長性能分析[J]. 江蘇農(nóng)業(yè)科學(xué),2019,47(21):247-248.
[3]任守文,王學(xué)敏,葛云山,等. 蘇鐘豬在云南省的引種、擴(kuò)繁和推廣情況[J]. 江蘇農(nóng)業(yè)科學(xué),2011,39(5):281-282.
[4]付言峰,李碧俠,李 蘭,等. 蘇鐘豬脂蛋白脂肪酶的組織表達(dá)譜及其生物信息學(xué)分析[J]. 畜牧獸醫(yī)學(xué)報(bào),2012,43(12):1871-1878.
[5]吳云鶴. 淮豬若干福利飼養(yǎng)技術(shù)研究[D]. 南京:南京農(nóng)業(yè)大學(xué),2020.
[6]李玉蓮,吳買生,王建偉,等. 大約克、杜洛克、巴克夏豬肥育和胴體性狀比較分析[J]. 家畜生態(tài)學(xué)報(bào),2018,39(3):44-47.
[7]付言峰,王澤平,廖 超,等. 雜交導(dǎo)入巴克夏血統(tǒng)蘇紫豬生長發(fā)育性能分析[J]. 江蘇農(nóng)業(yè)科學(xué),2022,50(17):169-173.
[8]趙云翔,鄺偉鍵,高 寧,等. 杜洛克公豬背膘厚度、日增重、日采食量和飼料效率相關(guān)性狀的遺傳參數(shù)估計(jì)[J]. 家畜生態(tài)學(xué)報(bào),2019,40(11):18-21.
[9]徐 忠,肖 倩,張 哲,等. 金華豬雜交組合繁殖、育肥、胴體與肉質(zhì)性能的研究[J]. 畜牧與獸醫(yī),2018,50(3):6-9.
[10]李碧俠,趙為民,付言峰,等. 苜蓿草粉對蘇山豬屠宰性能、胴體品質(zhì)和肉質(zhì)性狀的影響[J]. 動(dòng)物營養(yǎng)學(xué)報(bào),2020,32(3):1090-1098.
[11]任守文,李碧俠,葛云山,等. 蘇山豬選育研究[J]. 中國畜牧雜志,2020,56(11):47-51.
[12]丁祝進(jìn),崔虎軍,谷昭天. 魚類趨化因子家族的研究進(jìn)展[J]. 中國水產(chǎn)科學(xué),2021,28(9):1227-1237.
[13]Maurer M,von Stebut E. Macrophage inflammatory protein-1[J]. The International Journal of Biochemistry & Cell Biology,2004,36(10):1882-1886.
[14]Nomiyama H,Osada N,Yoshie O. The evolution of mammalian chemokine genes[J]. Cytokine & Growth Factor Reviews,2010,21(4):253-262.
[15]王秋霞,王 芳,楚富茗,等. 禽源趨化因子CCL4基因的表達(dá)及多克隆抗體的制備[J]. 中國獸醫(yī)科學(xué),2020,50(1):93-98.
[16]Tan K,Geng R J,Wang Z Q,et al. Anatomical structure,and expression of CCL4 and CCL13-like during the development of maxillary barbel in Paramisgurnus dabryanus[J]. Organogenesis,2019,15(1):13-23.
[17]Weissenbach M,Clahsen T,Weber C,et al. Interleukin-6 is a direct mediator of T cell migration[J]. European Journal of Immunology,2004,34(10):2895-2906.
[18]王麗萍. 低氧運(yùn)動(dòng)對食源性肥胖大鼠體重及血清炎癥因子TNF-α、IL-6的影響[D]. 南昌:江西師范大學(xué),2019.
[19]Hunter C A,Jones S A. IL-6 as a keystone cytokine in health and disease[J]. Nature Immunology,2015,16(5):448-457.
[20]李宇思,劉慧慧,沈松英,等. 母體外周血TNF-α、IL-6水平與子代出生體重的關(guān)系[J]. 中國生育健康雜志,2021,32(4):306-310.
[21]查安東,邢桂玲,高曉雯,等. IL6基因內(nèi)含子3多態(tài)性與仔豬腹瀉、生長和公豬繁殖性狀的關(guān)聯(lián)分析[J]. 畜牧獸醫(yī)學(xué)報(bào),2017,48(9):1674-1682.
[22]何 巖,劉 沛,韓廣業(yè),等. 成纖維細(xì)胞生長因子2通過AKT和ERK信號(hào)促進(jìn)腎癌細(xì)胞增殖[J]. 現(xiàn)代泌尿外科雜志,2019,24(3):227-232.
[23]方 幸. 運(yùn)動(dòng)介導(dǎo)肌肉因子IGF-1、FGF-2對小鼠骨的影響[D]. 上海:華東師范大學(xué),2018.
[24]Sanderson R D,Yang Y,Kelly T,et al. Enzymatic remodeling of heparan sulfate proteoglycans within the tumor microenvironment:growth regulation and the prospect of new cancer therapies[J]. Journal of Cellular Biochemistry,2005,96(5):897-905.
[25]Meyer A E,Pfeiffer C A,Brooks K E,et al. New perspective on conceptus estrogens in maternal recognition and pregnancy establishment in the pig[J]. Biology of Reproduction,2019,101(1):148-161.
[26]Kottakis F,Polytarchou C,F(xiàn)oltopoulou P,et al. FGF-2 regulates cell proliferation,migration,and angiogenesis through an NDY1/KDM2B-miR-101-EZH2 pathway[J]. Molecular Cell,201 3(2):285-298.
[27]王 睿. FGF9、miR-122在肥胖發(fā)生及預(yù)測中的作用[D]. 上海:上海交通大學(xué),2016.
[28]陳 鑫,公紅斌,李清春,等. 不同品系杜洛克后代商品豬育肥性能和屠宰性能的比較分析[J]. 石河子大學(xué)學(xué)報(bào)(自然科學(xué)版),2022,40(6):735-740.
[29]Sahni A,Altland O D,F(xiàn)rancis C W. FGF-2 but not FGF-1 binds fibrin and supports prolonged endothelial cell growth[J]. Journal of Thrombosis and Haemostasis,2003,1(6):1304-1310.
[30]陳東平,羅 茜,黃 佩,等. 趨化因子CCL3、CCL4在兒童免疫性血小板減少癥中的表達(dá)及臨床意義[J]. 臨床兒科雜志,2022,40(2):95-100.
[31]鄭琳琳. 透穴埋線對肥胖脂肪肝大鼠血清TC、TG、LDL-C、HDL-C及肝細(xì)胞的影響[D]. 鄭州:河南中醫(yī)學(xué)院,2012.
