豬輪狀病毒G1P[7]株分離鑒定及多克隆抗體制備
2024-04-11 06:10:32李素芬湯學(xué)超周金柱王丹丹朱雪蛟陶然李運(yùn)川郭容利張雪寒李彬
畜牧與獸醫(yī) 2024年4期
關(guān)鍵詞:檢測(cè)
李素芬,湯學(xué)超,周金柱,王丹丹,朱雪蛟,陶然,李運(yùn)川,郭容利,張雪寒*,李彬*
(1. 南京農(nóng)業(yè)大學(xué)動(dòng)物醫(yī)學(xué)院,江蘇 南京 210095;2. 江蘇省農(nóng)業(yè)科學(xué)院獸醫(yī)研究所,江蘇 南京 210014)
輪狀病毒(rotavirus,RV)感染與多種哺乳動(dòng)物和鳥(niǎo)類的腸炎和腹瀉有關(guān)[1]。RV是非囊膜二十面體病毒顆粒,包含11段雙鏈 RNA基因組[1]。病毒粒子由3層病毒結(jié)構(gòu)蛋白組成,VP2構(gòu)成核心,VP6構(gòu)成內(nèi)層,VP7和VP4構(gòu)成外殼,外殼包含血清型/基因型特異性表位。根據(jù)VP6蛋白的抗原性可將RV分類為12個(gè)血清群(A~L)[2-3],A、B、C、E和H群RV已經(jīng)被證實(shí)可感染豬,其中感染人和動(dòng)物最常見(jiàn)的為A群輪狀病毒(RVA)。
豬輪狀病毒A群(PoRVA)感染仔豬呈世界流行,各國(guó)豬場(chǎng)中 PoRVA 的陽(yáng)性率呈明顯走高趨勢(shì),報(bào)道最高感染率可達(dá)70%以上,引起世界各國(guó)重視[4-5],常與豬流行性腹瀉病毒(PEDV)等其他腹瀉病原混合感染,給世界養(yǎng)豬業(yè)造成嚴(yán)重的經(jīng)濟(jì)損失[6-7]。2種外衣殼蛋白(VP7和VP4)刺激中和抗體的產(chǎn)生,形成了RVA菌株G(糖蛋白)和P(蛋白酶敏感)雙基因分型系統(tǒng)的基礎(chǔ)。這種雙基因分型系統(tǒng)通常用于RVA分類,并由輪狀病毒分類工作組不斷更新。根據(jù)這一分類體系,全球已描述了42種G和58種P基因型[8-9]。G/P雙基因組合的多樣性是免疫保護(hù)的重要決定因素,與疫苗研制高度相關(guān)[8,10]。因此,持續(xù)監(jiān)測(cè)流行的RVA毒株對(duì)于了解區(qū)域流行病學(xué)信息和更新新型疫苗毒株非常重要。
小鼠EDIM毒株和猿猴SA-11毒株是能夠在細(xì)胞培養(yǎng)中繁殖的第一批RV毒株[11-12]。牛和人源RV分別于1971年和1980年在細(xì)胞培養(yǎng)中首次分離出來(lái)[13-14]。本研究旨在從臨床發(fā)病豬樣品中分離PoRV,對(duì)分離毒株的VP4和VP7基因進(jìn)行序列和遺傳進(jìn)化分析,為疫苗研制儲(chǔ)備生物學(xué)材料,并且制備該分離毒株的多克隆抗體,為PoRV致病機(jī)制研究提供可靠的工具。
1 材料與方法
1.1 病料來(lái)源
從江蘇省南京市附近某豬場(chǎng)采集腹瀉豬糞便,將3份糞便與1×MEM(含雙抗,其中青霉素濃度為 200 U/mL,鏈霉素終濃度為200 mg/mL),按1∶3進(jìn)行均質(zhì)化,反復(fù)凍融2次,低溫12 000 r/min離心5 min,取上清液先后使用0.45 μm和0.22 μm濾器過(guò)濾,其中吸取部分用于 RNA 提取和RT -PCR檢測(cè),剩余部分用于病毒分離。
1.2 主要試劑、細(xì)胞和動(dòng)物
RNA提取試劑盒(Fast Pure Cell/Tissue Total RNA Isolation Kit V2),HiScript Ⅲ 1st Strand cDNA Synthesis Kit(+gDNA wiper)和2×GreenTaqMix 預(yù)混液購(gòu)自南京諾唯贊生物科技股份有限公司。MA104細(xì)胞由江蘇省農(nóng)業(yè)科學(xué)院獸醫(yī)所保存。MEM和胰酶購(gòu)自上海源培生物科技股份有限公司;四季青胎牛血清購(gòu)自浙江天杭生物科技股份有限公司;PoRVA 的VP6單克隆抗體由本實(shí)驗(yàn)室制備[15];His-sumo-VP4*P7(26~231 aa)重組蛋白由本實(shí)驗(yàn)室制備,是將VP4基因完整閱讀框的78~693位核苷酸序列克隆到pCold-sumo質(zhì)粒,大腸桿菌表達(dá)純化所得,濃度為2 mg/mL;FITC-羊抗小鼠二抗、FITC-羊抗兔二抗和HRP-羊抗兔二抗購(gòu)自武漢博士德生物科技有限公司。新西蘭大白兔2只,體重3 kg/只,采購(gòu)自金陵種兔場(chǎng)。
1.3 引物設(shè)計(jì)與合成
本研究使用引物主要用于檢測(cè)PEDV、傳染性胃腸炎(TGEV)、豬德?tīng)査跔畈《?PDCoV)、RV、偽狂犬病毒(PRV)和牛病毒性腹瀉病毒(BVDV),委托南京金斯瑞生物科技有限公司合成,如表1。
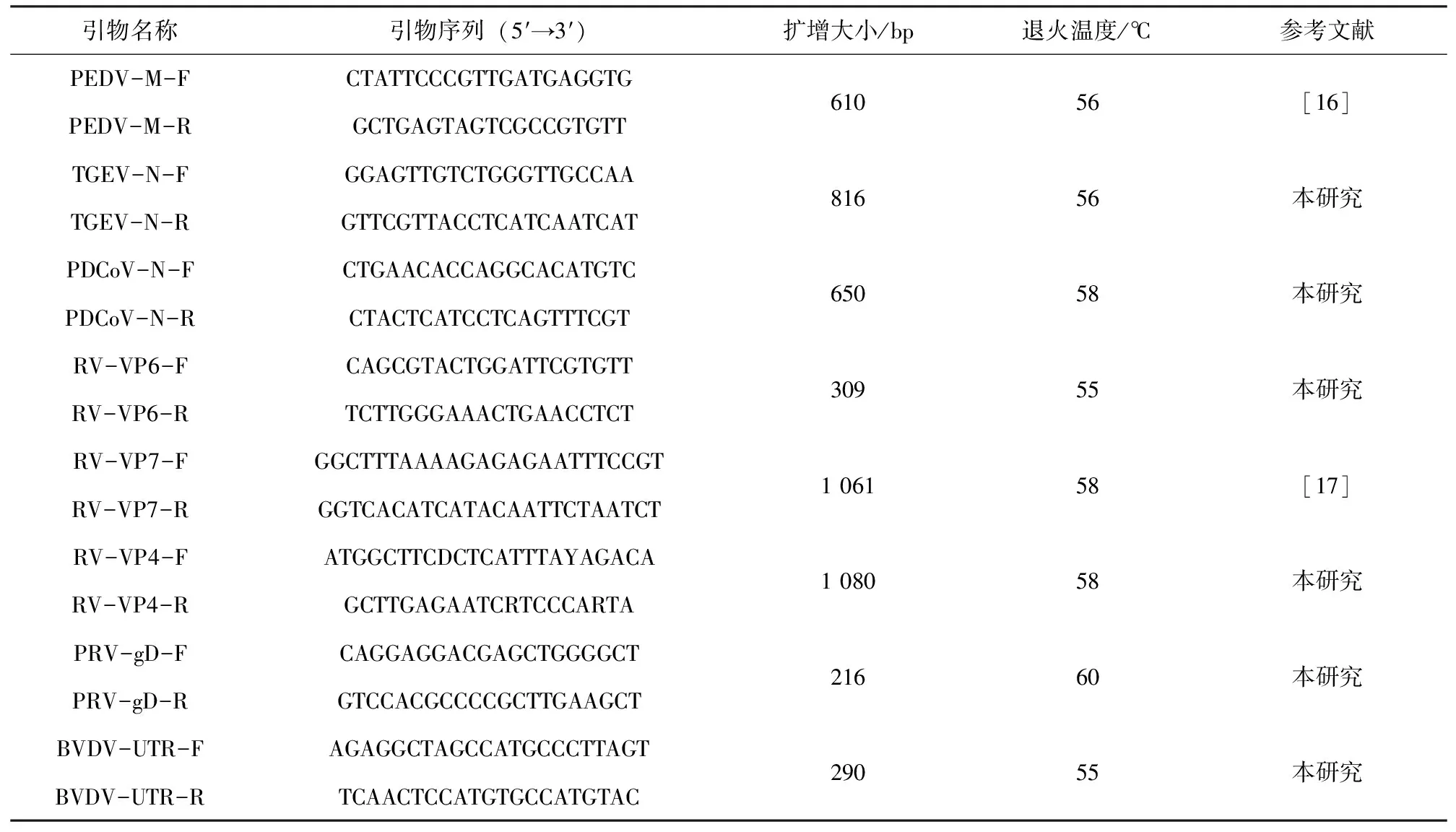
表1 引物信息
1.4 病原的RT-PCR檢測(cè)
吸取1.1中處理的上清液樣品100 μL,參照核酸提取試劑盒說(shuō)明書提取總RNA和DNA。按照反轉(zhuǎn)錄試劑盒HiScript Ⅲ 1st Strand cDNA Synthesis Kit(+gDNA wiper)說(shuō)明書進(jìn)行反轉(zhuǎn)錄獲得cDNA。利用2×GreenTaqMix和6種腹瀉相關(guān)病毒(PEDV、TGEV、PDCoV、RV、PRV和BVDV)的檢測(cè)引物進(jìn)行檢測(cè)。PEDV和TGEV:95 ℃預(yù)變性3 min;95 ℃ 30 s,56 ℃ 30 s,72 ℃ 1 min,30個(gè)循環(huán);72 ℃延伸10 min。PDCoV:95 ℃預(yù)變性3 min;95 ℃ 30 s,58 ℃ 30 s,72 ℃ 1 min,30個(gè)循環(huán);72 ℃延伸10 min。PRV:95 ℃預(yù)變性3 min;95 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s,30個(gè)循環(huán);72 ℃延伸10 min。BVDV和RV-VP6:95 ℃預(yù)變性3 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,30個(gè)循環(huán);72 ℃延伸10 min。
1.5 病毒分離和傳代
將長(zhǎng)滿單層的MA104細(xì)胞進(jìn)行消化和傳代,鋪1塊6孔板,待細(xì)胞長(zhǎng)滿單層后接毒。首先將1.1中過(guò)濾處理上清液預(yù)處理,加入終濃度為10 μg/mL胰酶,置于37 ℃激活30 min后備用。先用1×PBS洗滌6孔板單層細(xì)胞,將前述預(yù)處理過(guò)上清液加入至6孔板中,于5% CO2培養(yǎng)箱中使病毒與細(xì)胞孵育1 h。之后取出6孔板,棄掉上清液,并用1×PBS洗滌2次,加入無(wú)血清、胰酶終濃度為2 μg/mL的MEM維持液,連續(xù)觀察細(xì)胞病變(CPE)不少于3 d,待細(xì)胞有80% CPE時(shí)收獲病毒液,反復(fù)凍融2次,收獲病毒。相同的方法連續(xù)傳代。
1.6 間接免疫熒光(IFA)鑒定
將MA104細(xì)胞鋪于96孔板中,按照1.5方法進(jìn)行病毒感染,病毒感染后24 h左右即將出現(xiàn)細(xì)胞病變時(shí),棄掉上清液,用預(yù)冷的1×PBS洗滌細(xì)胞3次后加入200 μL預(yù)冷的冰乙醇,4 ℃固定30 min;再PBS洗滌3次后加入5%脫脂奶粉封閉1 h;棄掉封閉液,PBS洗滌3次之后向孔中加入50 μL稀釋的PoRV VP6蛋白單克隆抗體(1∶2 500),37 ℃孵育1 h;棄掉一抗,用PBS洗滌3次,加入50 μL稀釋的FITC-羊抗小鼠二抗(1∶2 000),37 ℃孵育45 min;棄掉二抗,用PBS洗滌3次,顯微鏡下觀察并拍照。
1.7 VP4和VP7基因擴(kuò)增和序列分析
RV-VP4-F和RV-VP4-R、RV-VP7-F和RV-VP7-R擴(kuò)增VP4和VP7基因片段,反應(yīng)程序?yàn)椋?5 ℃預(yù)變性3 min;95 ℃變性30 s,58 ℃退火30 s,72 ℃延伸1 min 10 s,共35個(gè)循環(huán);72 ℃延伸10 min。擴(kuò)增產(chǎn)物送南京擎科生物科技有限公司進(jìn)行測(cè)序。利用生物信息學(xué)軟件DNAStar對(duì)VP4(表2)和VP7(表3)基因與NCBI中的G基因型和P基因型代表毒株序列進(jìn)行核苷酸同源性分析和遺傳進(jìn)化分析。

表2 RV P[7](VP4基因)參考序列信息
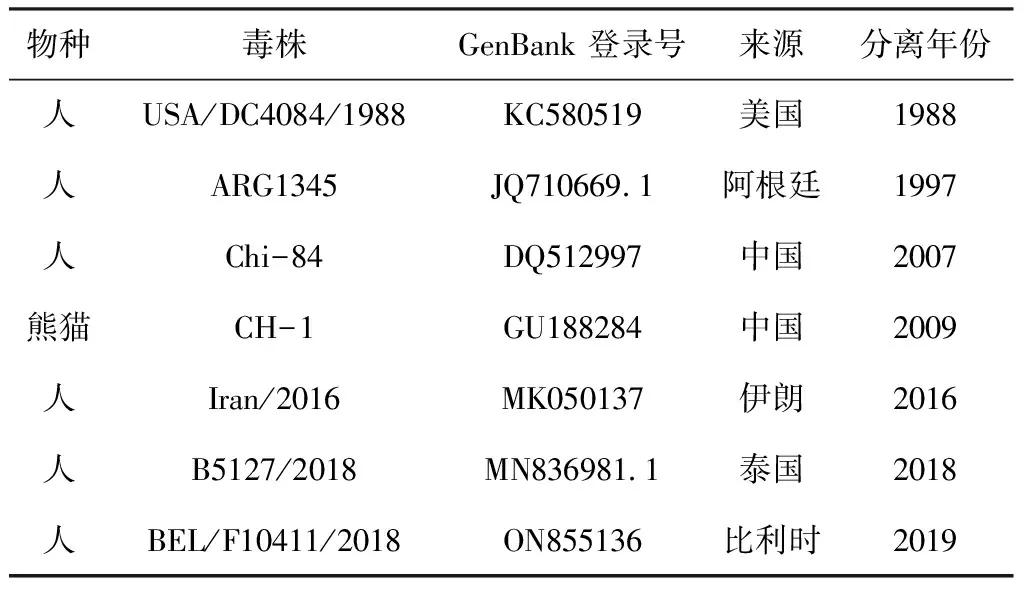
表3 RV G1(VP7基因)參考序列信息
1.8 兔源多克隆抗體制備和效價(jià)測(cè)定
將滅活劑β-丙內(nèi)酯按照0.05%比例加入到PoRV(TCID50=106/mL)病毒液中,充分混勻后4 ℃滅活24 h,經(jīng)滅活檢驗(yàn)合格后,與MONTANIDETM ISA 201VG佐劑按照1∶1混合乳化制備的免疫原,背部皮下注射新西蘭大白兔,1 mL/只。共免疫3次,首免和2次加強(qiáng)免疫之間分別間隔21 d,分別于免疫前和免疫后耳靜脈采血測(cè)定抗體效價(jià)。
采用中和抗體方法測(cè)定抗體效價(jià)。首先將200 TCID50的病毒液與10倍比稀釋的兔血清等體積混合,放37 ℃孵育1 h,然后加入到長(zhǎng)滿單層MA104的96孔板中,5% CO2培養(yǎng)箱中繼續(xù)孵育1 h。然后棄掉病毒血清混合液后,加入含有終濃度為2 μg/mL胰酶的病毒維持液(MEM),繼續(xù)培養(yǎng)。同時(shí)設(shè)置免疫前血清對(duì)照,所有試驗(yàn)孔和對(duì)照孔都設(shè)置3個(gè)重復(fù)。每日觀察并統(tǒng)計(jì)細(xì)胞病變情況,計(jì)算中和效價(jià)。
1.9 兔源多克隆抗體的反應(yīng)性鑒定
Western blot和IFA對(duì)多克隆抗體反應(yīng)性鑒定。IFA參照1.6方法操作,將PoRV VP6單克隆抗體替換為兔源多克隆抗體(1∶10 000),替換使用FITC-羊抗兔熒光二抗(1∶2 000),其他操作相同,顯微鏡下觀察并拍照。
Western blot檢測(cè)方法:吸取實(shí)驗(yàn)室保存的P7血清型重組蛋白His-sumo-VP4*P7,10 μL,按照1/5(體積比)加入5×上樣緩沖液,震蕩混勻加熱處理10 min后,上樣進(jìn)行SDS-PAGE分析,然后使用半干式轉(zhuǎn)印儀進(jìn)行免疫印跡操作。轉(zhuǎn)印結(jié)束,使用5%脫脂乳封閉醋酸纖維(NC)膜,洗滌后加入兔源多克隆抗體(1∶20 000),室溫孵育2 h,棄掉一抗,用PBS洗滌3次,加入稀釋的HRP-羊抗兔二抗(1∶10 000),室溫孵育1 h;棄掉二抗并洗滌后,ECL方法曝光顯色并拍照。
2 結(jié)果
2.1 腹瀉相關(guān)病原的檢測(cè)
將腹瀉樣品提取RNA,RT-PCR分別檢測(cè)RV、PEDV、TGEV、PDCoV、PRV和BVDV,只有RV-VP6引物成功擴(kuò)增出長(zhǎng)度為309 bp的條帶(圖1),而PEDV、TGEV、PDCoV和BVDV核酸全為陰性。提取腹瀉樣品DNA,PCR擴(kuò)增PRV,也呈現(xiàn)陰性擴(kuò)增。擴(kuò)增6種病毒陽(yáng)性對(duì)照全部成立。
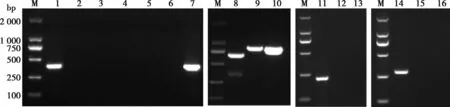
M.DL2000 分子量標(biāo)準(zhǔn);1.RV;2.PEDV;3.TGEV;4.PDCoV;5~6.RV陰性對(duì)照;7.RV陽(yáng)性對(duì)照;8~10.分別為PEDV、TGEV和PDCoV陽(yáng)性對(duì)照;11.PRV陽(yáng)性對(duì)照;12.PRV陰性對(duì)照;13.PRV;14.BVDV陽(yáng)性對(duì)照;15.BVDV陰性對(duì)照;16.BVDV。
2.2 病毒分離、鑒定和傳代培養(yǎng)
將RV陽(yáng)性濾液接種單層MA104細(xì)胞,初次接種48 h后,顯微鏡下即可觀察到融合、圓縮,最后瓦解脫落的CPE。按照相同方法傳至第5代,VP6基因檢測(cè)均為陽(yáng)性,且都可以產(chǎn)生明顯CPE。通過(guò)IFA檢測(cè),感染細(xì)胞呈現(xiàn)RV VP6蛋白特異性熒光(圖2),表明成功分離到RV,將其命名為JSNJ2019株。新分離病毒適應(yīng)細(xì)胞需傳代以提高病毒滴度。將JSNJ2019在MA104細(xì)胞上連續(xù)傳到第11代(P11),并分別測(cè)定半數(shù)組織細(xì)胞感染量(TCID50),病毒滴度從P8到P11逐步升高,介于105~106.5TCID50/mL(圖3)。
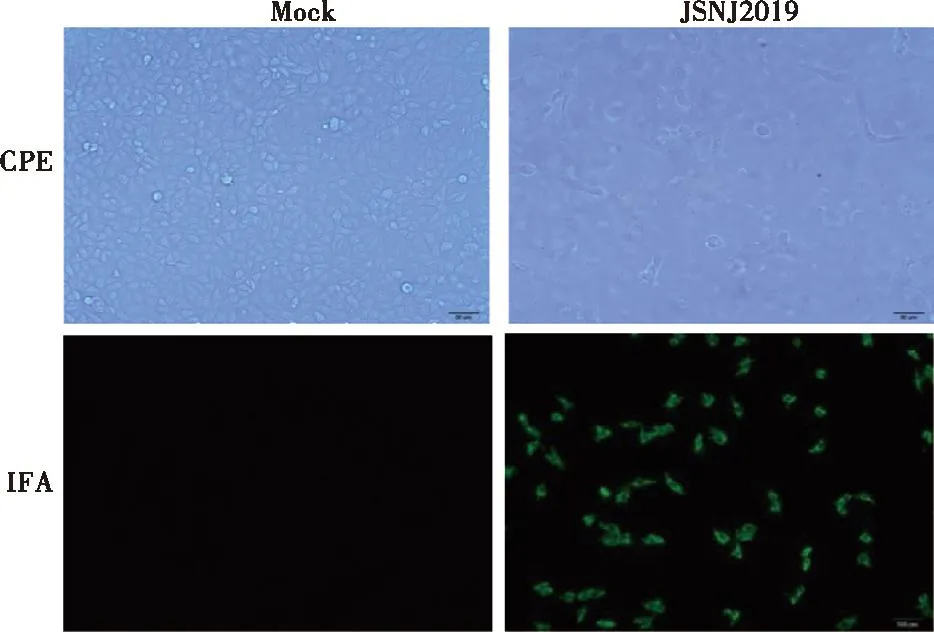
圖2 PoRV接種MA104細(xì)胞出現(xiàn)的病變
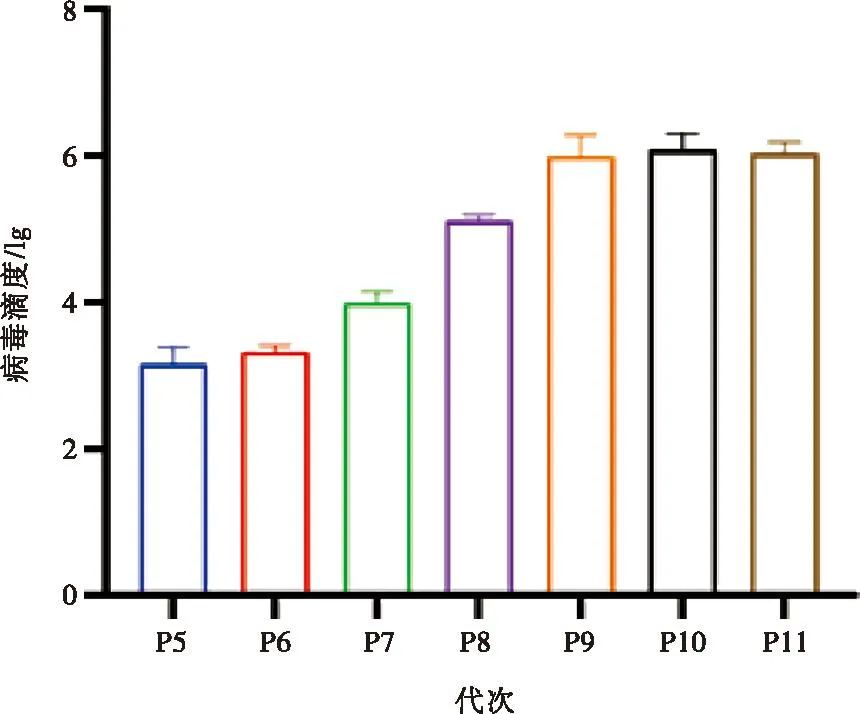
圖3 PoRV JSNJ2019不同代次滴度
2.3 VP4和VP7基因擴(kuò)增、序列同源性和遺傳進(jìn)化分析
VP4基因決定PoRV的P基因型,設(shè)計(jì)引物采用RT-PCR擴(kuò)增出特異性條帶,測(cè)序后序列比較確定為P[7]基因型。利用DNAStar軟件對(duì)7株P(guān)[7]基因型的PoRV的VP4基因進(jìn)行序列相似性比對(duì)分析,發(fā)現(xiàn)本試驗(yàn)分離毒株JSNJ2019株與參考毒株的PoRV VP4基因核苷酸相似性為91.8%~99.8%(圖4)。使用DNAStar軟件中的Clustal W方法對(duì)7個(gè)NCBI數(shù)據(jù)庫(kù)來(lái)源和JSNJ2019株的VP4基因序列進(jìn)行比對(duì)分析,并構(gòu)建遺傳進(jìn)化樹(shù)(圖5),發(fā)現(xiàn)JSNJ2019株單獨(dú)形成1個(gè)進(jìn)化分支,介于豬源與牛源之間的分支。
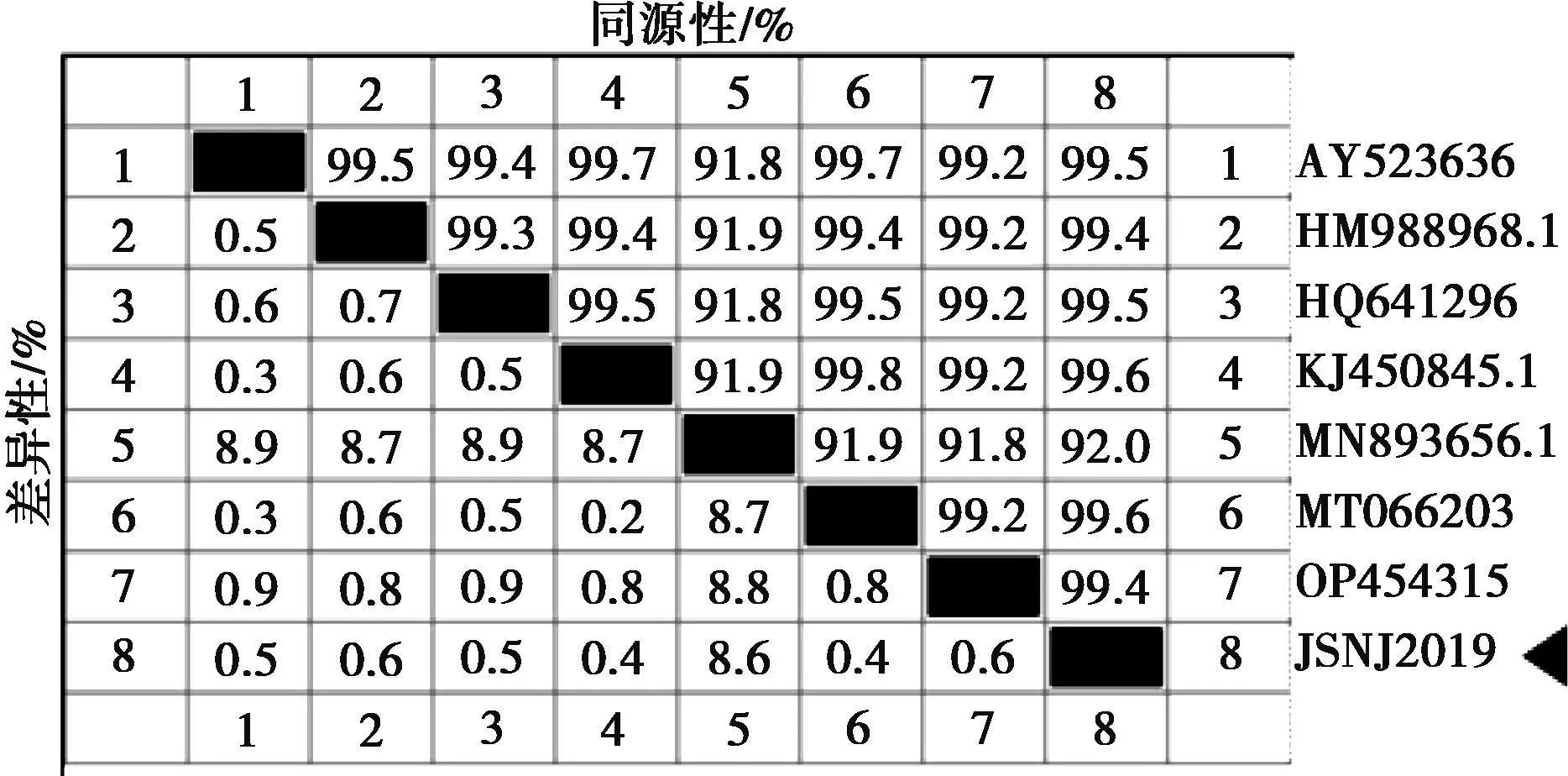
注:?為本研究分離株,下同。
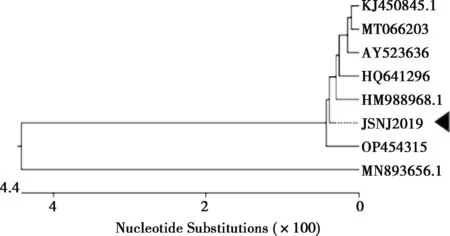
圖5 VP4基因遺傳進(jìn)化分析
VP7基因是決定PoRV的G基因型,設(shè)計(jì)引物采用RT-PCR擴(kuò)增出特異性條帶,測(cè)序后序列比較確定為G1基因型。利用DNAStar軟件對(duì)7株G1基因型PoRV的VP7基因進(jìn)行序列相似性比對(duì)分析,發(fā)現(xiàn)本試驗(yàn)分離毒株JSNJ2019株與參考毒株的PoRV VP7基因核苷酸相似性為96.3%~99.6%(圖6)。使用DNAStar軟件中的Clustal W方法對(duì)7個(gè)NCBI數(shù)據(jù)庫(kù)來(lái)源和JSNJ2019株的VP7基因序列進(jìn)行比對(duì)分析,并構(gòu)建遺傳進(jìn)化樹(shù)(圖7),發(fā)現(xiàn)JSNJ2019株與熊貓?jiān)吹脑谕贿M(jìn)化分支。
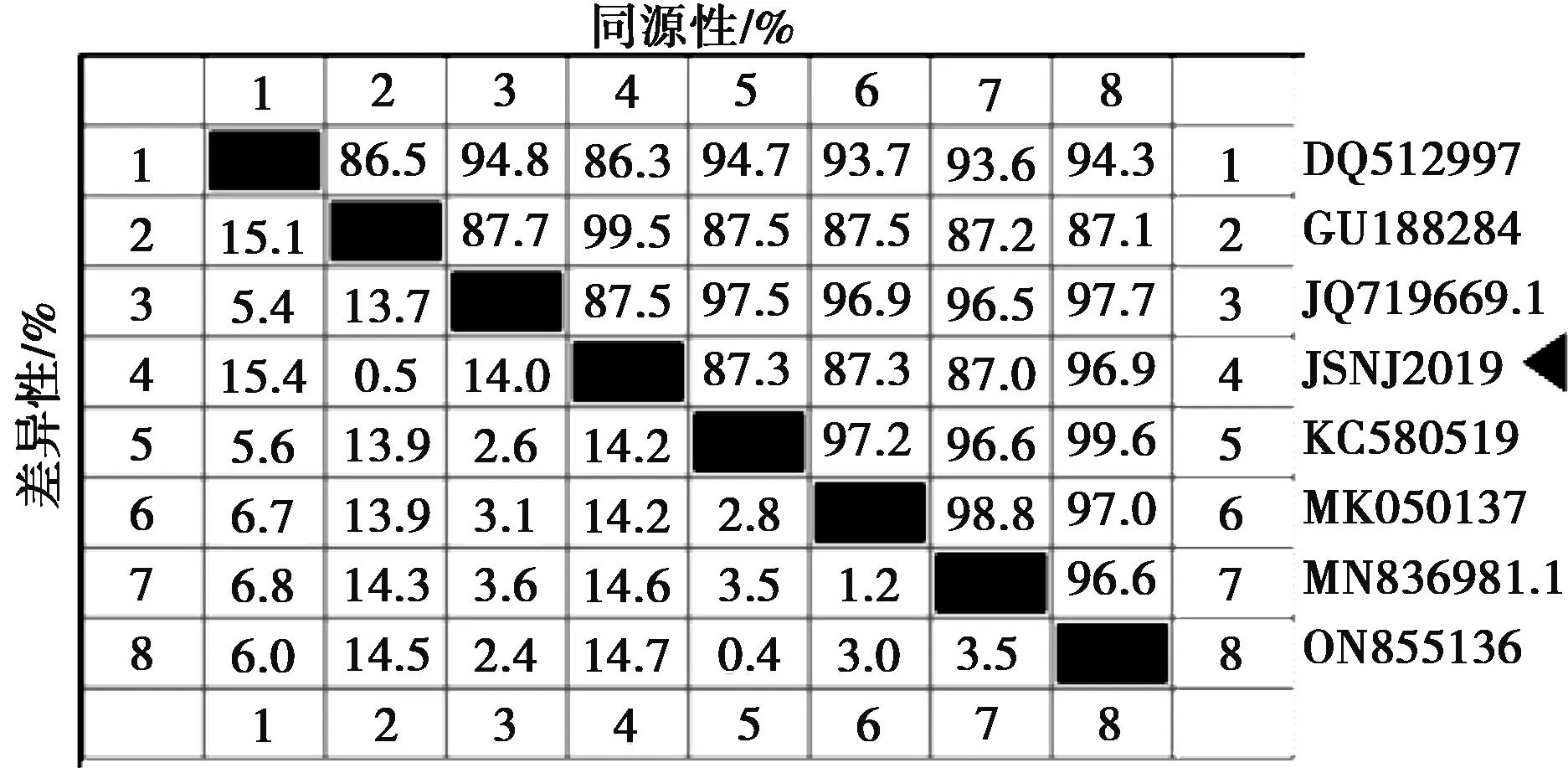
圖6 VP7基因同源性分析
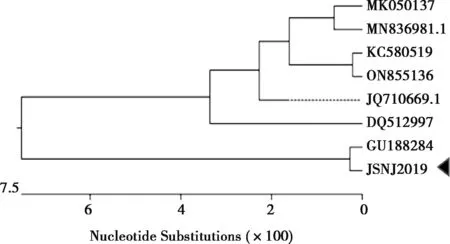
圖7 VP7基因遺傳進(jìn)化分析
綜上,確定JSNJ2019毒株基因型是G1P[7]。
2.4 高免血清制備和檢測(cè)
以JSNJ2019全病毒滅活苗為免疫原制備的抗血清,2倍比稀釋后測(cè)定中和JSNJ2019的能力,3次免疫后中和抗體效價(jià)最高可達(dá)211(圖8A)。同時(shí)取JSNJ2019第10代病毒接種單層MA104細(xì)胞,進(jìn)行IFA檢測(cè),鏡下可見(jiàn)明顯的綠色熒光(圖8B)。Western blot檢測(cè)顯示,只有PoRV重組蛋白His-sumo-VP8*P[7]出現(xiàn)特異性條帶,分子量38 kDa(圖8C)。

注:C圖中M為預(yù)染中分子量標(biāo)準(zhǔn),1為His-sumo標(biāo)簽蛋白,2~3為His-sumo-VP4*P[7]。
3 討論
PoRV是呼腸孤病毒科輪狀病毒屬的成員。到目前為止,國(guó)內(nèi)外流行病學(xué)數(shù)據(jù)顯示PoRV的感染率逐年升高,而且A群PoRV是豬場(chǎng)絕對(duì)優(yōu)勢(shì)的流行血清群[18-19]。PoRVA主要感染新生仔豬腸上皮細(xì)胞,多與PEDV等豬腹瀉病原混合感染而加重豬腹瀉,甚至死亡[20],糞便是腹瀉相關(guān)病原的主要傳播介質(zhì),通過(guò)糞-口和接觸傳播方式感染其他豬[21]。本研究采集腹瀉豬新鮮糞便,首先通過(guò)RT-PCR鑒別檢測(cè),只有PoRV為陽(yáng)性擴(kuò)增且條帶特異,經(jīng)測(cè)序分析進(jìn)一步明確VP6基因型為A群PoRV。
VP7和VP4蛋白決定PoRV的G型與P型以及它的毒力[2]。G/P型之間會(huì)產(chǎn)生不同的組合,不同組合型的PoRVA毒株誘導(dǎo)的交叉保護(hù)性低[22-23]。因此,加強(qiáng)不同G/P型的監(jiān)測(cè)和毒株分離對(duì)系統(tǒng)了解PoRVA的遺傳進(jìn)化和研發(fā)時(shí)效疫苗非常重要。本研究中從腹瀉豬糞便中檢測(cè)到G1和P[7]的組合基因型,成功分離到毒株,并可以穩(wěn)定傳代,國(guó)內(nèi)鮮見(jiàn)報(bào)道從豬體分離到豬源G1P[7]型,豐富了國(guó)內(nèi)PoRVA的資料,有利于對(duì)國(guó)內(nèi)RVA的監(jiān)測(cè)。VP4基因序列遺傳進(jìn)化分析發(fā)現(xiàn)JSNJ2019株單獨(dú)形成1個(gè)進(jìn)化分支,介于豬源與牛源之間的分支;VP7基因序列遺傳進(jìn)化進(jìn)一步發(fā)現(xiàn),JSNJ2019株與我國(guó)2009年報(bào)道分離的熊貓來(lái)源的CH-1毒株在同一個(gè)進(jìn)化分支。由此可見(jiàn),PoRVA不同G/P基因型組合具有明顯的多樣性和復(fù)雜性,對(duì)有效防控PoRVA的流行和感染具有很大的挑戰(zhàn)。
綜上,通過(guò)對(duì)南京附近豬腹瀉樣本主要流行腹瀉病原的檢測(cè),發(fā)現(xiàn)僅PoRVA為陽(yáng)性,通過(guò)系列分型檢測(cè)和序列分析,首次鑒定G1P[7]基因型組合在豬群的流行;進(jìn)一步采用敏感細(xì)胞系傳代培養(yǎng)和血清學(xué)鑒定成功分離得到G1P[7]型毒株JSNJ2019,并制備了高滴度的兔源多克隆抗體,表現(xiàn)出優(yōu)秀的免疫原性。因此,本研究豐富了PoRV流行病學(xué)數(shù)據(jù),并為時(shí)效疫苗的研制奠定了物質(zhì)基礎(chǔ)。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
