小麥黃花葉病抗性研究及育種應用進展
2024-04-17 01:00:40范德佳王汝琴何震天張容王建華韓燕陳士強
核農學報 2024年5期
范德佳 王汝琴 何震天 張容 王建華 韓燕 陳士強
(江蘇里下河地區農業科學研究所,江蘇 揚州 225007)
小麥黃花葉病是一種在世界各地廣泛發生的土傳病毒病,可導致小麥產量大幅度降低,其中中國有約200萬公頃冬小麥的生產受到小麥黃花葉病的威脅。該病害在我國最早發現于四川省雅安市,目前已擴展至長江中下游以及淮河流域,在山東、河南、江蘇、安徽、湖北、陜西等地相繼發生[1-3]。其癥狀與非生物脅迫(營養缺乏、霜凍或澇漬等)導致的老葉褪綠不同,黃化斑點在小麥幼苗的新生葉上即可表現出來。一般而言,受感染小麥植株在營養生長期新生葉呈現黃色,在拔節期生長發育遲緩或成熟延遲,最終導致產量下降,病害嚴重時小麥加工和烘焙品質也會大幅降低。我國的小麥黃花葉病主要由小麥黃花葉病毒(wheat yellow mosaic virus,WYMV)和中國小麥花葉病毒(Chinese wheat mosaic virus,CWMV)侵染導致,WYMV在中國和日本均有分布,而CWMV僅分布在中國的山東、河南、河北、江蘇等地[1]。為解決小麥黃花葉病造成的小麥減產、品質下降等問題,國內外研究人員從小麥黃花葉病的病原學、發病規律、抗源篩選、抗性遺傳和抗病育種等方面進行了大量研究。本文將重點介紹WYMV及其抗性遺傳相關研究進展,并對抗性基因在小麥抗病育種中的應用進行展望。
1 小麥黃花葉病毒(WYMV)
1.1 結構特征
WYMV屬于馬鈴薯Y病毒科(Potyviridae)大麥黃花葉病毒屬(Bymovirus),和同屬的小麥梭條花葉病毒(wheat spindle streak mosaic virus,WSSMV)在發病癥狀、傳毒特性、粒子形態、外殼蛋白、基因序列等方面非常相似。最開始我國的小麥黃花葉病被認為是由WSSMV引起的,但是RNA序列分析表明中國和日本的小麥黃花葉病是由WYMV引起,兩者是不同的物種。WYMV是一種由外殼蛋白包裹RNA組成的絲狀粒子,其基因組為2條正義單鏈RNA(圖1),其中RNA1長約7.3 kb,編碼8個非結構蛋白和1個外殼蛋白,即P3、PIPO(pretty interestingPotyviridaeORF)、7K、細胞質內含體(cytoplasmic inclusion,CI)、14K、基因組連接蛋白(viral protein genome-linked,VPg)、細胞核內含體A-蛋白酶(nuclear inclusion protein A-proteinase,NIa-Pro)、細胞核內含體B(nuclear inclusion protein B,NIb)和C端的外殼蛋白(coat protein,CP);RNA2約為3.5 kb,編碼病毒蛋白P1和P2[4]。病毒RNA在5'端與VPg蛋白相連,3'端有一個poly(A)尾,與真核生物中成熟mRNA類似,有助于RNA在宿主細胞中復制[5]。
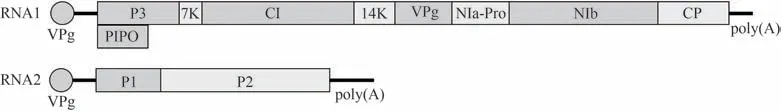
圖1 小麥黃花葉病毒(WYMV)的基因組結構Fig.1 Genomic organization of wheat yellow mosaic virus (WYMV)
1.2 宿主及其分布
WYMV的宿主主要是小麥屬(Triticum),也能夠侵染山羊草屬(Aegilops)和少數大麥屬(Hordeum)物種[6]。由于其復制依賴宿主真核翻譯起始因子4E(eukaryotic translation initiation factor 4E,eIF4E)和病毒VPg之間的相互作用,所以eIF4E-VPg交互模塊是宿主特異性的一個決定因素。例如,WYMV在大麥原生質體中不能復制,因此不侵染大麥,而在表達了小麥eIF4E后卻能夠復制;同樣,將大麥黃花葉病毒(barley yellow mosaic virus,BaYMV)的VPg替換成WYMV的VPg,也導致BaYMV病毒不能在大麥原生質體中復制[7]。此外,病毒RNA2編碼的P1蛋白是植株系統感染所必需的,WYMV與BaYMV的P1蛋白只有60%的相似性,可能也是宿主特異性的決定因素之一[8]。
WYMV主要分布于中國和日本。Sun等[2]分析了中國五個省的11個不同地點的小麥植株上的WYMV蛋白CP(297個序列)和VPg(87個序列)的核苷酸序列,發現所有序列之間存在密切的系統發育關系,不同族群的聚類與其來源地高度一致,表明WYMV種群遺傳結構與起源地密切相關。Ohki等[9]根據小麥品種是否被系統侵染,將來自日本各地的14種WYMV分離株劃分為致病型Ⅰ、Ⅱ或Ⅲ,進一步分析發現不同分離株的RNA1和RNA2編碼的氨基酸序列均存在差異,三種致病型分布于日本不同地區。上述對于該病毒致病型和分布差異的分析有利于宏觀掌握小麥黃花葉病的發生規律和流行特點,并針對性地采取有效防控措施。
1.3 傳播特性
大麥黃花葉病毒屬病毒可從禾谷多黏菌(Polymyxa graminis)的游動孢子轉移到幼苗根細胞,表明該類病毒以禾谷多黏菌為介體通過土壤傳播。禾谷多黏菌分布于世界各地,主要在谷類作物及其野生近緣種的根部繁殖,產生可移動的初級和次級游動孢子、休眠厚垣孢子。目前已知大麥黃花葉病毒屬(Bymovirus)、真菌傳桿狀病毒屬(Furovirus)和花生叢簇病毒屬(Pecluvirus)的14種土傳植物病毒以禾谷多黏菌為傳播載體[10]。禾谷多黏菌傳播病毒的能力受到土壤水分和溫度的影響,土壤水分對游動孢子到達宿主根部至關重要。而病毒從根部感染部位移動到葉片需要低于20 ℃的溫度。當平均溫度超過20 ℃時,病害癥狀會在受感染的葉子上停止發展;但在適宜條件下,在新葉上會重新出現癥狀。由此可見,該病毒一旦發生就會對受侵染的田地構成持久的威脅。另外,此類病毒經常與其他病毒同時感染宿主,如WYMV與中國小麥花葉病毒(Chinese wheat mosaic virus,CWMV)混合侵染且后具有協同作用[11]。目前通過特異性逆轉錄重組酶聚合酶擴增技術(reverse transcription-recombinase polymerase amplification,RT-RPA)或高通量測序的方法可有效鑒定病毒種類[12-13],為病害的精準防治提供參考依據。
1.4 病毒的復制與擴散
病毒在宿主中的完整生命周期涉及多個生物過程,由于RNA病毒只編碼少數蛋白質,成功的病毒感染取決于病毒和宿主之間復雜的相互作用網絡。首先,病毒需要招募一系列宿主蛋白來完成復雜感染周期中的所有步驟,包括病毒顆粒分解、病毒翻譯、病毒復制復合體(viral replication complex,VRC)的形成、病毒粒子組裝、細胞間移動和長距離運輸。
病毒復制與細胞質中病毒誘導的細胞內膜結構形成相關,植物細胞被感染后會產生由內質網(endoplasmic reticulum,ER)聚集形成的膜狀體(membranous body,MB),成為病毒復制復合體進行組裝的支架[14]。利用免疫電鏡觀察WYMV侵染的細胞發現,病毒蛋白P1、P2及P3是膜狀體的重要組成部分,而且P2能夠重新排列ER并形成大的聚合結構,與其他病毒蛋白互作并將它們招募到聚合體中[15]。由于病毒基因組較小且編碼能力有限,病毒需要借助宿主因子來合成自身成分。在真核生物中,宿主蛋白eIF4E與mRNA的5'-末端m7G帽子結構結合,形成蛋白質翻譯起始復合物[16]。而馬鈴薯Y病毒科病毒的VPg與病毒RNA的5'-末端連接,模擬m7G帽子結構與eIF4E互作結合,以促進病毒蛋白的翻譯起始。在體外試驗中,WYMV中RNA1的5'-UTR有一個獨特的內部核糖體進入位點(internal ribosome entry site,IRES)可以調節病毒翻譯水平,而3'-UTR的多聚腺苷酸化可抑制病毒基因組RNA互補鏈的合成[17-18]。此外,病毒外殼蛋白除了精確組裝病毒顆粒外,還協助WYMV的VPg獲得適當的構象以進行核質穿梭,促進病毒RNA的翻譯[19]。
一般情況下,病毒可以通過胞間連絲移動到相鄰細胞,導致系統性感染。馬鈴薯Y病毒科病毒的VPg除了參與復制外,還與宿主中一種富含半胱氨酸的蛋白質,即馬鈴薯Y病毒屬VPg互作蛋白(PotyvirusVPg-interacting protein,PVIP)相互作用,促進病毒在細胞間的移動[20]。WYMV的細胞質內含體CI蛋白也被證明介導了病毒的細胞間或長距離運動,而且CI導致了致病型Ⅰ和Ⅱ的致病性差異[21]。WYMV的P3NPIPO是一種致病性決定簇,通過轉錄滑移在保守的G2A6位點表達[22]。由于P3N-PIPO對馬鈴薯Y病毒的胞間移動非常重要,推測該蛋白可能參與WYMV的細胞間擴散[23]。WYMV的7K和14K蛋白都含有一個中心疏水跨膜結構域,可能參與病毒復制囊泡的形成、內質網輸出及細胞間移動[1]。
另外,WYMV的外殼蛋白與小麥熱休克蛋白家族蛋白TaHSP23.6發生相互作用,影響WYMV的感染[24]。病毒NIb蛋白與宿主光誘導蛋白(light induced protein,LIP)相互作用通過擾亂小麥中的脫落酸途徑促進其感染[25]。WYMV還可以通過m6A甲基化破壞免疫相關基因的表達,從而抑制宿主的抗病毒反應[26]。總之,WYMV和宿主相互作用的研究還很缺乏,需要繼續深入研究,以期為小麥黃花葉病抗性育種提供理論支撐。
2 小麥黃花葉病的抗性遺傳
2.1 抗病種質資源篩選
我國較早開展了小麥黃花葉病抗源的篩選,鑒定和創制出許多抗性資源。如何震天等[27]選育出的揚輻麥9311對小麥黃花葉病免疫。李鵬等[3]育成了高抗小麥黃花葉病的小麥品種魯原502。阮義理等[28]、周益軍等[29]采用田間自然誘發鑒定出免疫材料52份、高抗材料41份,包括儀寧小麥、西風等。孫炳劍等[30]在河南省主要推廣品種中篩選到對小麥黃花葉病免疫的品種新麥208。魏瑋等[31]從國內外收集了527個小麥品種(系),經WYMV病圃鑒定,發現有10個品種(系)的抗性相對比較穩定,并以抗病和感病品種為親本篩選出具有多態性的SSR標記。吳斌等[32]對山東80個小麥生產品種進行了WYMV抗性評價,發現濟麥22等15個品種達“抗”級。劉迪等[33]在河南省漯河市兩個重病田塊對146個小麥品種(系)進行了抗性鑒定,檢測到存麥16號等免疫品種(系)12個,周麥26等高抗品種(系)37個。Fouchard等[34]鑒定了158份法國品種在SBWMV和WYMV復合侵染條件下的抗性,三分之一的品種表現抗病性,其中僅1%表現WYMV感病癥狀。此外,研究發現高抗品種(系)多為廣譜抗性品種,如侯慶樹等[35]在9個病區毒源接種地發現寧豐小麥、西風、蘇麥3號等品種(系)高抗所有毒源;岳緒國等[36]研究表明西風、儀寧小麥、RF-1等品種對6種毒源均表現高抗。上述研究發現的抗病種質資源為小麥黃花葉病的抗性研究及抗病品種選育奠定了基礎。
2.2 抗性基因的挖掘
在小麥染色體2A、2DL、3BS、4D、5AL、6DS、7BS上共鑒定到14個小麥黃花葉病抗性基因或數量性狀位點(quantitative trait locus,QTL)(表1)。小麥2D染色體長臂上具有一個抗性基因熱點區域,包含來自中國小麥種質揚輻麥9311的YmYF[37]和儀寧小麥的QYm.nau-2D[38],來自歐洲種質Ibis的Ym1b[39],來自美國種質Madsen的Qym1[40]和YmMD[41]以及來自日本種質Yumechikara的Q.Ymym[42]等。其中,QYm.nau-2D在雜交育種中由于重組而以六種不同的狀態存在,序列分析表明該片段來源于山羊草屬(Aegilops)N基因組的滲入[43]。然而,染色體2DL上這些基因之間的等位性還不清楚,也沒有研究進行分離和歸類,以上有待后續研究。
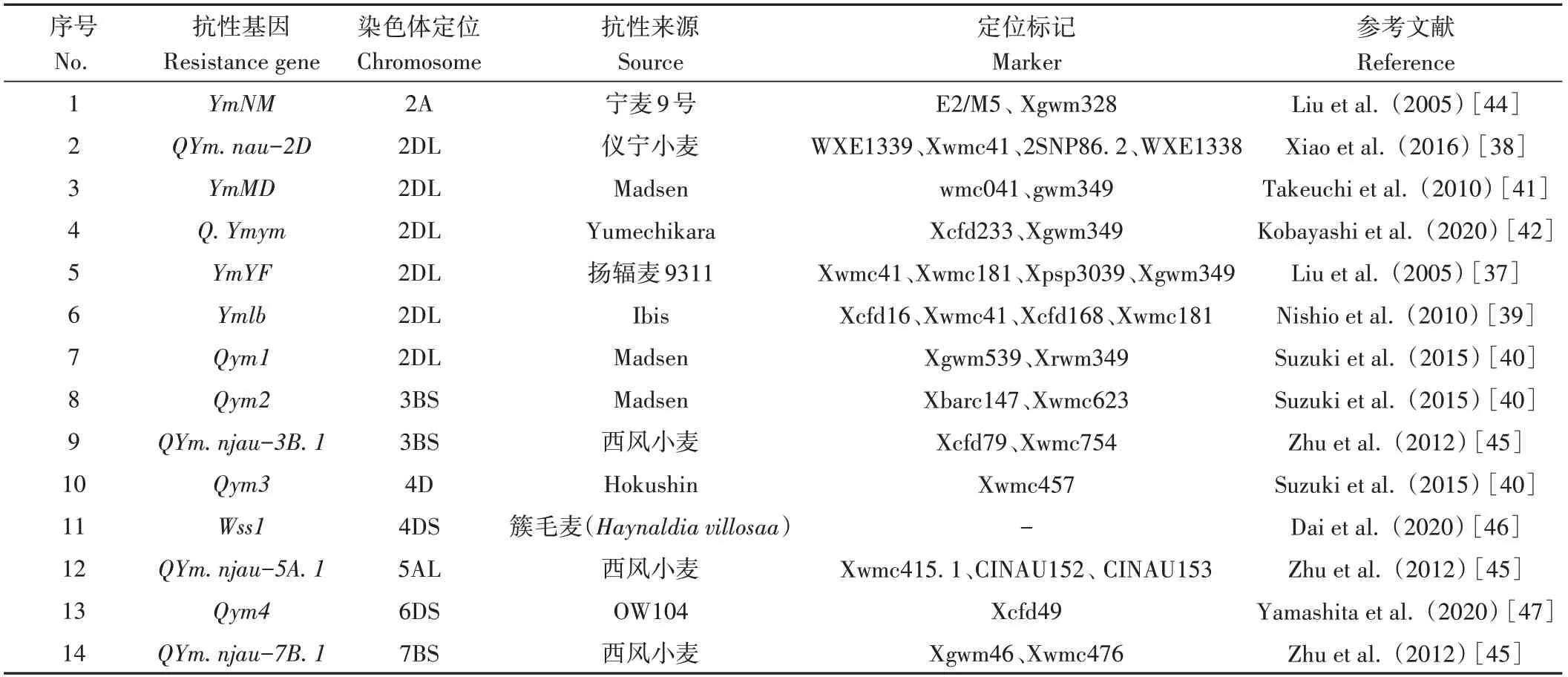
表1 已鑒定出的抗小麥黃花葉病基因及數量性狀位點Table 1 Identified resistance genes and QTLs against WYMV
小麥其他染色體上也有抗病位點分布,如在染色體2A上有YmNM[44],3BS上有Qym2[40]和QYm.njau-3B.1[45],4D上有Wss1(來源于簇毛麥)[46]和Qym3[40],5AL上有QYm.njau-5A.1[45],6DS上有Qym4[47],7BS上有QYm.njau-7B.1[45]。
2.3 抗性基因的克隆和驗證
Ym2是首個圖位克隆的顯性抗WYMV基因,編碼一種CC-NBS-LRR蛋白,位于小麥3BS染色體上主效QTL區域(Qym2),來源于小麥的近緣物種——沙融山羊草(Aegilopssharonensis)[48]。Ym2可以限制WYMV在根中的積累,通過回交方法將該基因轉入感病小麥品種可增強其抗病性,提高發病麥區小麥產量,具有重要的應用價值。TaPDIL5-1基因是小麥中首個被證實的抗病毒基因,該基因是大麥HvPDIL5-1的同源基因,編碼一種蛋白二硫鍵異構酶,利用CRISPR/Cas9同時敲除小麥A、B、D亞基因組上TaPDIL5-1的3個拷貝可顯著增強小麥黃花葉病抗性,而不影響株高、穗數、千粒重等農藝性狀,具有主要的育種利用價值[49]。同時敲除小麥eIF4E基因的3個拷貝,也可賦予小麥對黃花葉病的完全抗病性,表明TaeIF4E作為感病因子基因參與WYMV感染[50]。TaVTC2同樣對WYMV侵染具有正調控作用,NIb與TaVTC2的互作抑制了TaVTC2的酶活性從而觸發植物免疫反應,過表達TaVTC2顯著增加了WYMV在小麥植株中的積累;而利用RNAi技術敲除TaVTC2可抑制WYMV的侵染,此外,敲除植株對CWMV和大麥條紋花葉病毒(BSMV)以及白粉病菌也表現出較好的抗性[51]。上述基因中,Ym2已廣泛應用于抗WYMV小麥種質創制及品種選育,TaPDIL5-1、TaeIF4E、TaVTC2是基因編輯技術創制抗病種質的可用靶標,具有很大的潛在應用價值。
2.4 抗病毒機制研究
RNA沉默是植物中重要的先天抗病毒防御機制,具有高效性、快速性和特異性。外源基因整合到宿主細胞產生的雙鏈RNA(double-Standed RNA,dsRNA)會被核糖核酸內切酶Dicer切割生成小干擾RNA(small interfering RNA,siRNA),siRNA與宿主AGO蛋白結合形成RNA誘導沉默復合物(RNA induced silencing complex,RISC),RISC可以靶向病毒RNA導致其降解或翻譯停滯[52]。植物中病毒感染會導致細胞內病毒衍生小干擾RNA(virus-derived small interfering RNA,vsiRNA)積累,vsiRNA可靶向病毒RNA使其特異性降解。利用RNA干擾技術使病毒基因沉默可賦予植株對病毒的持久抗性,如WYMVNIb8基因反義序列的小麥轉基因株系N12-1對小麥黃花葉病的抗性增強[53];轉CP蛋白基因的轉基因株系P8-T2對WYMV表現高抗[54]。WYMVNIb基因衍生的vsiRNA不僅可以通過誘導RNA沉默來抑制植物中的病毒感染,還可以靶向小麥基因TaAAED1以降解其轉錄產物,從而抑制活性氧(reactive oxygen species,ROS)清除,激活植物免疫[55]。小麥植株不同部位WYMV vsiRNA譜顯示葉和根中vsiRNA在病毒基因組中的分布不同,而且來自根的vsiRNA在5'末端的A/U偏好性較低,表明針對WYMV的RNA沉默在根和葉之間的作用模式是不同的[56]。然而,病毒也已經進化出各種策略來對抗植物的病毒防御反應,WYMV可能調節DCL4的表達以減少vsiRNA的生物發生,從而促進病毒感染[57];WYMV P1蛋白可發揮RNA沉默抑制活性,促進病毒在小麥植株中的感染[58]。
在抗性小麥品種中,土傳病毒在根部和莖葉之間的運動被阻止或延緩[59]。Haufler等[60]利用免疫吸附電子顯微鏡觀察抗病品種的抗性反應發現病毒只存在植株根部,表明抗性品種可抑制病毒復制和擴散。目前在小麥中已鑒定出一些調節病毒轉運的抗性基因,如Wss1[61]可以阻止WSSMV從根到分蘗的移動。抗病品種Madsen(含有Qym1、Qym2)的抗性在根部起作用,可以阻止人工接種的病毒從葉部進入根部[48,62],已克隆的Ym2基因在小麥根中發揮功能,抑制病毒的積累[48]。但目前尚無分子證據表明宿主因子參與WYMV或WSSMV病毒的細胞間或長距離移動。根系土壤微生物群落可以刺激植物激素的合成,與病原菌競爭營養物質,從而對植物的抗病性產生顯著影響[63-64]。與感WYMV的品種相比,抗WYMV的小麥品種在土壤中可檢測到更高的微生物多樣性和更穩定的群落結構。WYMV抗性品種在其根際土壤中大量招募許多已知的有益細菌和真菌類群,包括黃單胞菌目、放線菌目、鞘氨醇單胞菌屬等,而敏感品種根際土壤中具有更多的潛在病原體[65]。由此可見,構建良好的根際微生物群落可提高植物對土傳病害的抗性。
此外,植物抗病機制與轉錄組和蛋白組的調控相關。Zhang等[26]通過m6A-seq發現抗性相關基因的轉錄后修飾(N6-甲基腺苷甲基化)在小麥對WYMV的抗性中發揮重要作用,沉默甲基轉移酶基因(Triticum aestivumm6A methyltransferase B,TaMTB)可以顯著降低宿主植物中的WYMV積累。小麥組蛋白乙酰轉移酶(histone acetyltransferase,HAT)和組蛋白去乙酰化酶(histone deacetylase,HDAC)的表達在受到大麥條紋花葉病毒(barley stripe mosaic virus,BSMV)、中國小麥花葉病毒(CWMV)和小麥黃花葉病毒(WYMV)等侵染后表達量顯著增加,表明兩者在抗性調控中發揮著一定作用[66-67]。WYMV感染后,小麥一些泛素特異性蛋白酶(ubiquitin-specific protease,UBP)家族基因表達發生顯著變化,這些基因可以響應水楊酸(salicylic acid,SA)信號,參與小麥對病毒的防御反應[68]。WYMV NIb通過干擾脫落酸(abscisic acid,ABA)信號通路促進病毒感染,表明ABA信號途徑在小麥抗WYMV反應中發揮重要作用[25]。miR168是植物中最豐富和高度保守的miRNA之一,它可以直接調節AGO1的表達[69]。Wu等[70]在未感病小麥和WYMV感染的小麥之間鑒定出了131個差異表達的miRNA,對應的85個靶基因涉及植物激素信號轉導和單環菌素生物合成。糖基化是一種重要的蛋白質翻譯后修飾,涉及蛋白質的折疊、穩定性和酶活性,在植物的抗病反應中也發揮重要作用,如小麥幾丁質觸發子受體激酶(Triticumaestivumchitin elicitor receptor kinase,TaCERK)的過度表達減少了WYMV的積累,TaCERK的N-糖基化可以調節小麥對WYMV的抗性[71]。目前利用多組學技術挖掘抗性基因、探索抗性調控機制已成為常規方法,隨著人們對病原體與植物互作機制的深入了解,小麥抗病育種進程將不斷加快。
3 小麥黃花葉病的防治
3.1 綜合防治策略
禾谷多黏菌是WYMV等多種病毒的傳播媒介,通過栽培管理抑制真菌繁殖可有效防治病毒侵染和傳播。目前主要的防治措施有:(1)輪作倒茬:通過與非寄主作物油菜、馬鈴薯等蔬菜輪作,能夠改變禾谷多黏菌的生活條件,達到減少病原菌、減輕病害的目的;(2)選用抗病品種:能經濟有效地預防病害的發生;(3)加強田間管理:前茬收獲后及時深耕滅茬,避免病菌通過帶病殘體、病土、農事機具等途徑傳播,適時遲播以避開禾谷多黏菌的最適侵染時期;(4)農藥處理:種子播前消毒、包衣,對于發病田塊用殺菌劑進行土壤消毒(圖2)。
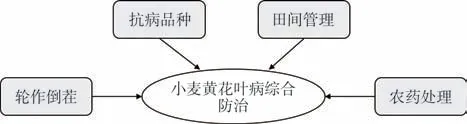
圖2 小麥黃花葉病的綜合防治策略Fig.2 Strategies for the control of wheat yellow mosaic disease
3.2 抗病品種的推廣與應用
目前,一批高抗小麥黃花葉病、豐產性好的小麥品種已成功選育并推廣應用。如江蘇省農業科學院以抗病品種西風為父本育成了寧麥9號,后續又育成寧麥13、寧麥17、寧麥26等抗病品種。江蘇太湖地區農業科學研究所以西風為母本,采用系譜法育成了抗病的蘇麥5號,后續又育成抗病品種蘇麥6號。江蘇省大華種業集團有限公司以蘇麥6號為親本選育出抗病品種華麥6號。江蘇丘陵地區鎮江農業科學研究所育成了抗WYMV的鎮麥5號、鎮麥9號、鎮麥12、鎮麥15等。江蘇里下河地區農業科學研究所利用抗病品種寧麥9號選育出超高產抗病品種揚輻麥4號,以其為親本育成揚輻麥6號、揚輻麥8號、揚輻麥9號等品種;以免疫種質揚輻麥9311育成抗病高產品種揚輻麥5號、揚輻麥7號、揚輻麥8167等一系列新品種(系)。其中,寧麥13連續多年為江蘇省淮南麥區第一大品種,揚輻麥4號連續多年為江蘇省主推品種、“江蘇好品種”。這些品種在江蘇省小麥黃花葉病發病區的推廣應用,對控制病害流行、挽回產量損失起到了十分重要的作用。
3.3 利用現代生物技術進行抗病種質創制及抗病品種選育
小麥對WYMV的遺傳抗性受多種因素的影響,開發和利用分子標記準確篩選WYMV抗性育種材料具有重要意義。如南京農業大學開發的11個QYm.nau-2D診斷標記為小麥WYMV抗性的改良提供了重要參考[43];最近日本學者利用與Madsen的Qym1和Qym2連鎖的分子標記育成了一個新的小麥育種系Kitami-94[72]。另外,近年來基因編輯技術在小麥的抗病育種中的應用備受關注,同時編輯小麥A、B、D亞基因組上TaPDIL5-1基因的3個拷貝和eIF4E的3個拷貝,都可賦予小麥對WYMV的抗性[49-50]。由此可見,通過基因編輯技術可以創制具有抗病性的小麥材料,為小麥的抗病毒病育種提供新思路。
4 總結與展望
近年來,小麥黃花葉病的抗性育種取得了許多進展,篩選出了一批高抗甚至免疫的種質資源,育成了一批抗病小麥品種(系),在小麥遺傳改良和生產應用方面發揮了重要作用。而且隨著小麥基因組測序完成、基因組功能的深入解析,小麥育種技術得到了快速發展。
目前,在小麥黃花葉病毒的抗病機理、抗性基因的挖掘與利用等方面需要進一步深入研究:
(1)病毒-宿主間互作機理研究。植物抗病性的可持續利用需要更深入地闡明病毒與宿主間的互作機制。雖然WYMV等大麥花葉病毒屬病毒的結構和基因組研究已較為充分,但病毒侵染小麥的分子基礎仍然不明,充分利用基因組學、蛋白組學等方法,可加快分子機制的解析。
(2)抗病資源篩選。目前育成品種的抗源遺傳基礎較狹窄,利用野生品種或近緣物種挖掘新的抗病基因資源已成為當務之急。因此,必須加強新抗源篩選,并結合現代分子育種技術高效培育小麥抗病新品種。
(3)數據庫建設和分子育種。一個整合小麥基因組功能和全基因組多態性標記的信息平臺,可以幫助人們更好地利用有育種價值的基因。通過表型篩選和標記輔助選擇相結合,育種家可以在更短的時間內培育出新的抗性小麥品種。
(4)基因編輯。基于CRISPR/Cas系統的基因編輯技術是目前最熱門的新興生物技術,許多研究團隊已利用CRISPR/Cas9成功編輯小麥的基因組,實現了小麥的抗病性改良。基因編輯技術可進行基因的敲除、表觀修飾、堿基編輯等多種基因修飾,在作物抗病性的遺傳改良中具有非常大的應用潛力。
