蚯蚓原位堆肥與不同比例生物炭對基質理化性質及番茄品質的作用研究
2024-04-17 00:57:36李文慧王繼濤安明遠徐廣亞尹翠游宏建曹云娥
核農學報 2024年5期
關鍵詞:生物
李文慧 王繼濤 安明遠 徐廣亞 尹翠 游宏建 曹云娥,*
(1寧夏大學葡萄酒與園藝學院,寧夏 銀川 750021;2寧夏回族自治區園藝技術推廣站,寧夏 銀川 750001;3寧夏共享人力集團有限公司,寧夏 銀川 750027)
番茄(SolanumlycopersicumL.)在我國種植廣泛且經濟利益較高,近年來,其生產方式基本由傳統的露地栽培轉變為設施栽培[1]。然而,設施農業高集約化和高復種指數導致土壤微生物群落失衡,土傳病害加重及土壤性質破壞等一系列土壤退化問題[2-3]。采用基質栽培是緩解此類問題的有效措施之一[4]。前人研究以菇渣、牛糞、蛭石為原料,進行了番茄基質配方的篩選,發現當牛糞∶菇渣∶蛭石體積比為6∶3∶1時,番茄的產量和品質顯著提高[5]。
然而,目前無土栽培相關的國標、行標等都較少,市面銷售的栽培基質物料配比、養分構成也不盡相同,難以開展標準化無土栽培體系研究[6]。蚯蚓(Pheretima)作為生態系統中常見的分解者,可以將有機固體廢棄物分解轉化為腐殖質,故蚯蚓堆肥作為一種富含有機質、養分和腐殖質的新型基質而備受關注[7-8]。相關研究表明,施用蚯蚓堆肥可使黑籽(NigellasativaL.)籽粒氮、磷、鉀含量提高13.50%、10.80%和14.10%,黑籽油中的亞油酸、油酸和亞麻酸含量提高53.53%、25.36%和0.60%[9]。為使蚯蚓堆肥長期穩定地向栽培體系輸送養分,保證番茄的穩產高質,本研究特引入“蚯蚓原位堆肥”(in-situ vermicomposting)——一種改良的蚯蚓堆肥技術。該技術將蚯蚓及共生菌群引入有機廢棄物(畜禽糞便、餐廚垃圾、農田果園殘留物等)中,其分解、轉化有機廢棄物的全過程不僅直接在待處理地塊自發完成,亦結合當地耕作制度實時補充有機廢棄物物料,以保證養分長期供應充足。此外,生物炭作為一種物理結構穩定、化學性質呈現惰性的富碳材料,被廣泛應用于農業研究[10]。生物炭可影響土壤微生物群落結構及多樣性,調控環境因子和微生物之間的互作關系,促進并改良微生物生態系統,從而對受微生物驅動的氮、磷、鉀營養元素循環轉化產生影響,最終改變土壤質量和功能[11]。Huang等[12]發現玉米芯生物炭既促進了細菌的數量和多樣性,又能進一步加速脫水污泥蚯蚓堆肥過程中有機物的礦化和腐殖化。同時,生物炭可有效減少NH3的揮發,進而將NH4+轉化為NO3-,達到氮肥的緩釋效果,為作物和根系生長提供充足的養分,提高氮素利用效率[13-14]。
目前針對蚯蚓(原位)堆肥和生物炭作為栽培基質的應用均較成熟,但將蚯蚓原位堆肥與生物炭配合施用,探究其對養分、菌群及農產品產量與品質構成的影響仍鮮有報道。因此,本研究通過向蚯蚓原位堆肥中添加不同比例的生物炭,探究比較不同處理的理化性質、菌群組成及番茄代謝發育特征,旨在為此后相關研究提供基礎數據。
1 材料與方法
1.1 試驗地概況
試驗于2021年2月24日—7月2日在寧夏賀蘭縣園藝產業園區的日光溫室(106.33°E,38.58°N)進行。該基地屬于國家級農業示范園區,年降雨量180~200 mm,80%的降雨量集中在6—8月,年平均氣溫8.8 ℃,無霜期160~170 d,屬典型的中溫帶干旱氣候區,具有大陸性氣候特征。
1.2 試驗材料
試驗番茄品種為粉得力(武漢楚為生物科技股份有限公司);試驗所需蚯蚓和牛糞購自寧夏萬輝生物環保科技有限公司;生物炭購自上海海諾碳工業有限公司。生物炭和蚯蚓原位堆肥基本特性見表1。

表1 基礎材料理化性質Table 1 Physicochemical properties of basic materials
1.3 試驗方法
采用單因素隨機區組設計。共設6個處理,分別為蚯蚓原位堆肥(CK)、蚯蚓原位堆肥+1%生物炭(C1)、蚯蚓原位堆肥+2%生物炭(C2)、蚯蚓原位堆肥+3%生物炭(C3)、蚯蚓原位堆肥+5%生物炭(C4)、蚯蚓原位堆肥+10%生物炭(C5),每個處理重復5次,隨機區組排列。
試驗將基質槽一分為二,種植壟和養殖壟尺寸均為4.1 m(長)×0.35 m(寬)×0.2 m(深),對照壟每壟鋪設蚯蚓原位堆肥226.73 kg,鋪設牛糞0.38 m3。根據等碳原則,種植壟添加生物炭替代蚯蚓原位堆肥中部分總碳,養殖壟投放一定密度蚯蚓處理牛糞產生蚯蚓堆肥。待種植壟一茬作物收獲,養殖壟中牛糞全部處理為蚯蚓堆肥后,養殖壟與種植壟模式調換,原養殖壟原位處理生成的蚯蚓堆肥中定植作物,原種植壟中重新鋪設牛糞進行蚯蚓原位處理,為下一茬作物定植做準備,最終實現作物原位輪作。番茄單行栽植于種植壟,株距15 cm,行距140 cm,所有處理采用水肥一體化滴管裝置進行水肥管理,番茄整個生育期灌水量約1 400 t·hm-2,肥料選用寧夏萬輝生物環保科技有限公司生產的茄果類專用滴灌配方肥(24.0% N+3.0% P2O5+21.0% K2O),全生育期共投入氮肥378 kg·hm-2、磷肥47.25 kg·hm-2、鉀肥330.75 kg·hm-2。
1.4 測定項目與方法
1.4.1 基質理化性質測定 于番茄盛果期(2021年5月18日),采用5點取樣法從栽培基質中部取基質500 g,每個處理5次重復,風干后過1 mm篩。根據馬全會等[15]的方法測定基質容重、孔隙度、含水量。使用pH儀和DDS-307A EC儀(上海精密科學儀器有限公司)測定基質pH值和電導率(electric conductivity,EC)(土∶水=1∶10)。有機質含量測定采用重鉻酸鉀氧化法;全氮(total N,TN)含量測定采用硫酸消煮-凱氏定氮-硼酸指示劑-0.01 mol·L-1硫酸滴定法;全磷(total P,TP)含量測定采用HCIO4-H2SO4消化-鉬銻抗比色法[16]。分別采用0.5 mol·L-1K2SO4、0.5 mol·L-1NaHCO3萃取-鉬銻法和NH4OAc萃取-火焰光度法測定速效氮(available N,AN)、速效磷(available P,AP)和速效鉀(available K,AK)含量[16]。
土壤脲酶活性采用苯酚鈉法測定;蔗糖酶活性采用3,5-二硝基水楊酸法測定;過氧化氫酶活性采用高錳酸鉀法測定;堿性磷酸酶活性采用磷酸苯二鈉法測定[16]。
1.4.2 微生物豐度指標測定 于番茄盛果期(2021年5月18日)采集新鮮基質樣品進行微生物群落多樣性分析。根據QIAamp DNA MiniKit試劑盒(Qiagenn,德國)說明書進行微生物群落總DNA抽提,使用1%的瓊脂糖凝膠電泳檢測DNA的提取質量,使用NanoDrop 2000(美國Thermo Fisher Scientific公司)測定其DNA純度和濃度。采用引物338F(5'-ACTCCTACGGGAG GCAGCAG-3')和806R(5'-GGACTACHVGGGTWTCT AAT-3')對細菌16S rRNA基因V3-V4的可變區進行PCR擴增及高通量測序;引物ITS2R(5'-GCTGCGTTC TTCATCGATGC-3')和ITS1F(5'-CTTGGTCATTTAGA GGAAGTAA-3')對真菌進行PCR擴增及高通量測序。細菌PCR反應體系:5×FastPfu Buffer緩沖液4 μL,2.5 mmol·L-1dNTPs 2 μL,5 μmol·L-1上、下游引物各0.8 μL,FastPfu Polymerase DNA聚合酶0.4 μL,模板DNA 10 ng,ddH2O補足至20 μL。真菌PCR反應體系:10×Buffer緩沖液2 μL,2.5 mmol·L-1dNTPs 2 μL,5 μmol·L-1上、下游引物各0.8 μL,rTAP Polymerase DNA聚合酶0.2 μL,模板DNA 10 ng,ddH2O補足至20 μL。程序為:95 ℃預變性3 min;95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸30 s,細菌27個循環,真菌35個循環;72 ℃終延伸10 min。測序數據優化處理、操作分類單元(operational taxonomic unit,OTU)聚類分析及注釋、多樣性及群落結構分析等均在上海美吉生物科技有限公司的I-Sanger平臺進行,用QIIME 1.9.1軟件進行稀疏分析,以區分OTU豐富度及α多樣性指數[覆蓋度(Coverage)、Shannon、Simpson、Chao1、Ace指數][17];使用CANOCO 5.0軟件對基質微生物門、屬水平群落結構、相對豐度等與基質理化性質進行冗余分析(redundancy analysis,RDA)。
1.4.3 番茄生長量、品質和產量測定 于盛果期,每個處理隨機選取植株6株,取下大小均勻果實10個,測定果實質量。可溶性固形物含量采用TD-45數字折光儀(中國金科利達公司)進行測定。有機酸和維生素C(vitamin C,Vc)含量分別通過NaOH滴定法和2,6 -二氯苯酚直接滴定法進行測定。此外,分別通過二硝基水楊酸和蒽酮比色法測定硝酸鹽和可溶性糖含量[18]。全生育期記錄每個處理產量,按照小區面積產量折合成番茄產量。
測定果實品質的同時,在每個處理中隨機選取植株6株。用天平測定地上部與地下部鮮重后,每個植株裝進信封并標記,將樣品置于烘箱105 ℃下殺青30 min,80 ℃烘干至恒重。用電子天平稱量地上部與地下部干重。
1.4.4 高效液相色譜-質譜聯用(chighperfomanceliquid chromatography-tandem mass spectrometry, LC-MS)檢測 在1 000 mg番茄鮮樣中加入1.3 mL提取液(甲醇∶水=4∶1,V∶V),利用Wonbio-96c型冷凍組織研磨儀(上海萬柏生物科技有限公司)破碎,低溫渦旋混勻處理30 min。4 ℃、13 000 r·min-1離心15 min,取上清后重復上述萃取操作3次,將上清液在氮氣下混合濃縮。最終,樣品用乙腈∶水=1∶100 μL(V∶V)復溶后用于液相色譜-質譜分析。上機完成之后,在美吉生物云平臺(https://cloud.majorbio.com)進行差異代謝物分析,篩選出差異代謝物,并通過KEGG(kyoto encyclopedia of genes and genomes)數據庫進行代謝通路注釋,獲得差異代謝物參與的通路[19]。
1.5 數據統計與分析
采用Excel 2021對數據進行整理與統計,采用SPSS 26.0軟件對數據進行單因素方差分析(ANOVA)及相關性分析,采用Origin 2021軟件作圖。
2 結果與分析
2.1 不同比例生物炭對蚯蚓原位堆肥理化性質的影響
由表2可知,添加生物炭增大了蚯蚓原位堆肥的總孔隙度,減小了容重,但各處理間無顯著差異(P>0.05)。
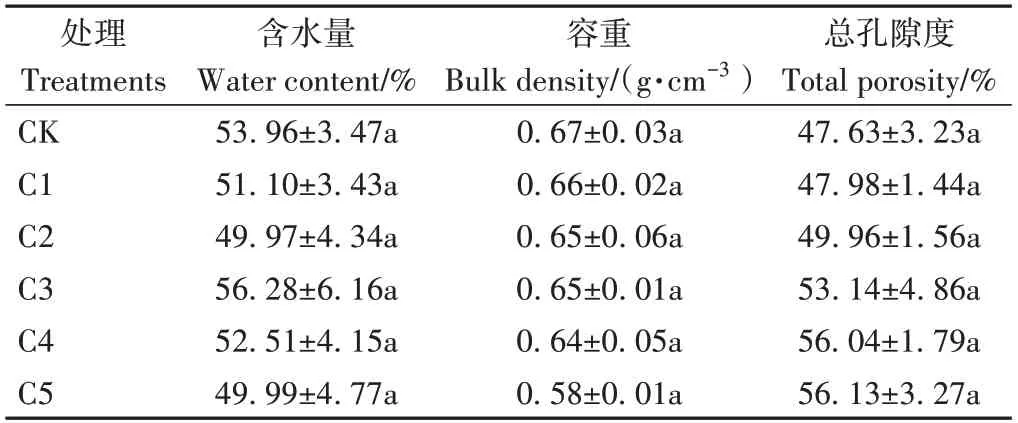
表2 生物炭對蚯蚓原位堆肥物理性質的影響Table 2 Effect of biochar on the physical properties of in-situ vermicomposting
由表3可知,與CK相比,添加生物炭能顯著提高蚯蚓原位堆肥的pH和EC值(P<0.05),并且隨著生物炭添加比例的增大,pH和EC值呈持續上升趨勢。有機質含量也表現出相同趨勢,從121.50 g·kg-1增加到310.50 g·kg-1。添加生物炭提高了蚯蚓原位堆肥的養分含量,并且隨著生物炭添加比例的增大,蚯蚓原位堆肥中全氮、速效氮、速效磷、全磷和速效鉀含量均呈現先增加后降低趨勢。其中,全磷、速效磷和速效鉀含量以C3處理最高,分別較CK顯著提高了46.27%、10.10%和68.43%;全氮和速效氮含量以C4處理最高,分別較CK顯著提高了15.96%和92.20%。
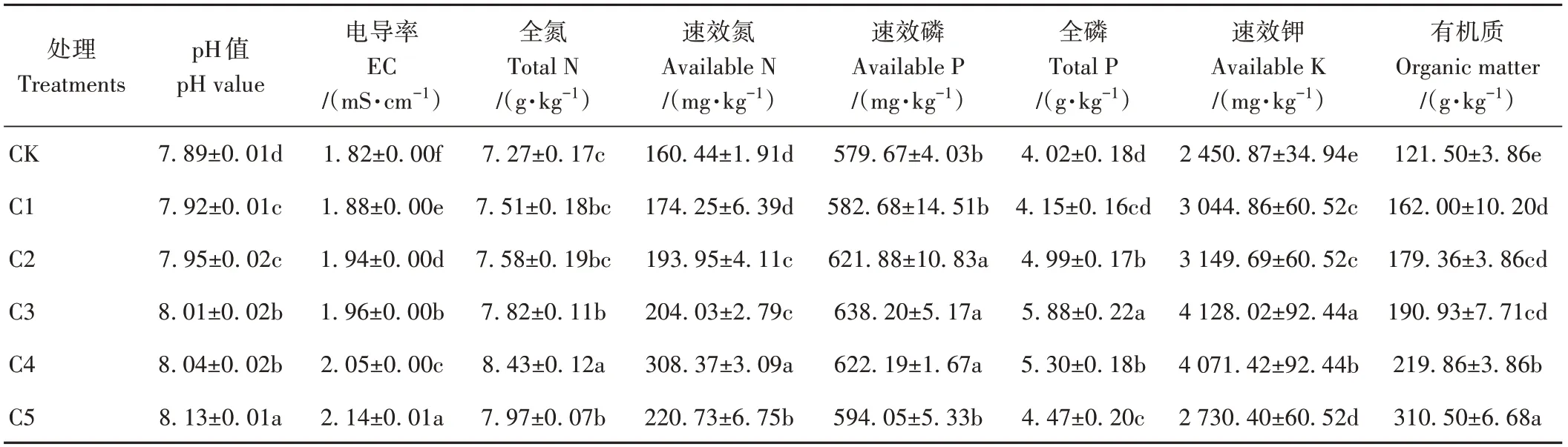
表3 生物炭對蚯蚓原位堆肥化學性質的影響Table 3 Effect of biochar on the chemical properties of in-situ vermicomposting
2.2 不同比例生物炭對蚯蚓原位堆肥酶活性的影響
由圖1可知,添加生物炭能提高蚯蚓原位堆肥中的酶活性,并且隨著生物炭添加比例的增大,過氧化氫酶活性呈持續增大趨勢;蔗糖酶、脲酶、堿性磷酸酶活性均呈先增大后減小趨勢。其中,蔗糖酶和堿性磷酸酶活性以C3處理最高,分別較CK顯著提高了52.09%和18.66%;脲酶活性以C4處理最高,較CK顯著提高了86.49%。
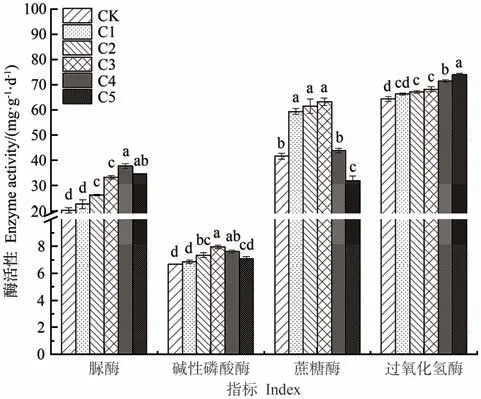
圖1 生物炭對蚯蚓原位堆肥酶活性的影響Fig.1 Effect of biochar on enzyme activity of in-situ vermicomposting
2.3 不同比例生物炭對蚯蚓原位堆肥微生物多樣性的影響
由圖2可知,細菌群落中各處理共產生5 567個OTU,其中共有OTU數量為2 422,占總數的43.51%。CK~C5處理特有OTU數分別為41、46、44、65、51、68。相較CK,C1~C5處理特有OTU數量均有不同程度增加,以C5處理特有OTU數量最多。這可能是因為生物炭多孔的結構有利于微生物的生長發育。由此表明,生物炭能增加蚯蚓原位堆肥中的特有細菌數量,對細菌的產生可能具有一定的促進作用。
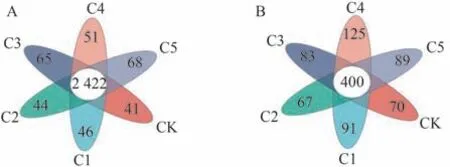
圖2 生物炭對蚯蚓原位堆肥細菌(A)和真菌(B)OTU水平的影響Fig.2 Effect of biochar on the OTU levels bacteria (A) and fungi (B) of in-situ vermicomposting
真菌群落中各處理共產生1 671個OTU,其中共有OTU數量為400,占總數的23.94%。CK~C5處理特有OTU數分別為70、91、67、83、125、89,除C2處理外,其他處理的特有OTU數量均高于CK,變化趨勢與細菌基本一致,以C4處理特有OTU最多。C5處理特有OTU小于C4處理的原因可能是過量生物炭對真菌的產生具有抑制作用。
為了進一步分析生物炭對微生物群落多樣性的影響,采用五種指標(覆蓋度、Ace指數、Chao1指數、Simpson指數以及Shannon指數)計算蚯蚓原位堆肥中的細菌和真菌豐度。由表4可知,細菌和真菌覆蓋度(Coverage)分別達96%和99%以上,說明蚯蚓原位堆肥中所測出的細菌、真菌序列數足以反映其真實情況。不同處理間細菌Chao1指數和Ace指數無顯著差異;隨著生物炭添加比例的增大,各處理細菌Shannon指數呈先降低后升高趨勢,Simpson指數呈先升高后降低趨勢,其中,C3處理的Shannon指數和Simpson指數相較CK差異顯著,細菌多樣性最低。真菌Chao1指數、Ace指數、Shannon指數和Simpson指數在不同處理間均無顯著差異。
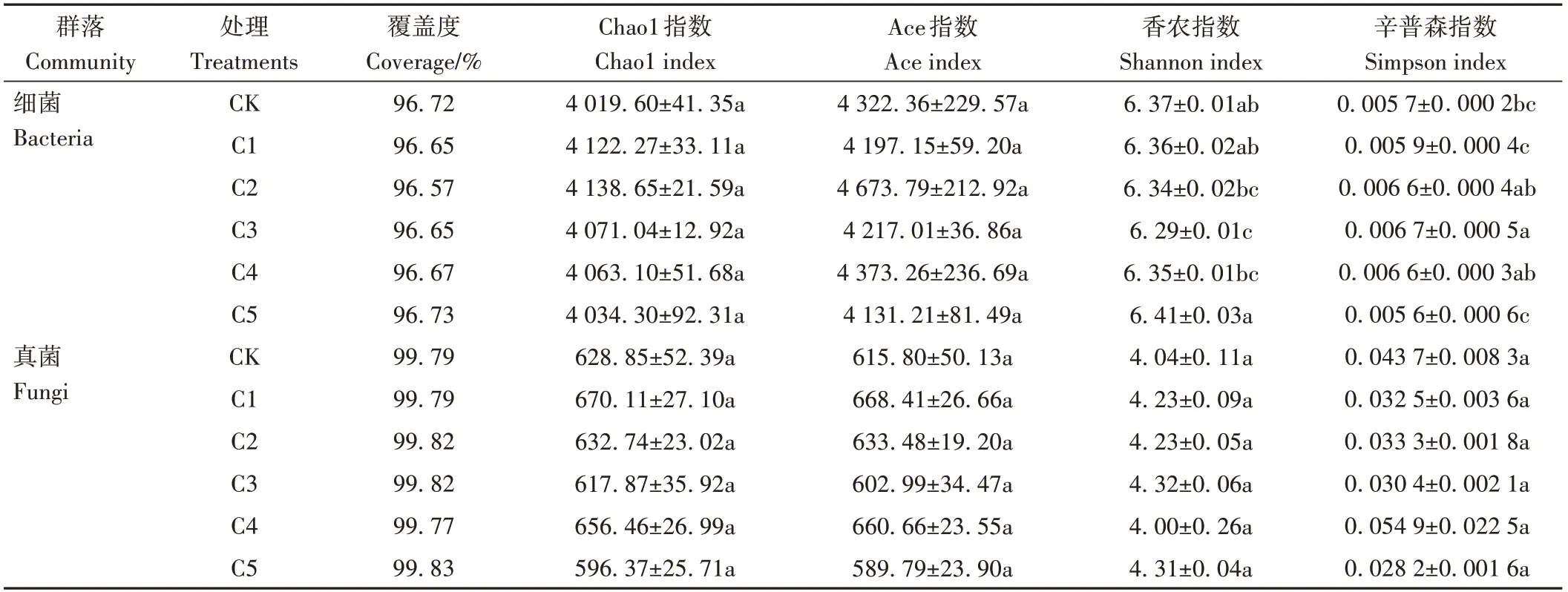
表4 生物炭對蚯蚓原位堆肥細菌和真菌群落Alpha多樣性指數的影響Table 4 Effect of biochar on Alpha diversity index of biochar on Alpha diversity index of in-situ vermicomposting with bacterial and fungal communities
2.4 不同比例生物炭對蚯蚓原位堆肥微生物群落組成的影響
由圖3可知,在細菌門水平上,不同處理得到12個相對豐度>1%的細菌菌門,其中優勢菌門為放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、綠彎菌門(Chloroflexi)和厚壁菌門(Firmicutes),共占到全部菌門相對豐度的87.44%~89.51%。CK~C5處理的綠彎菌門(Chloroflexi)相對豐度分別為12.69%、15.87%、13.93%、17.85%、16.39%和26.87%,變形菌門(Proteobacteria)和厚壁菌門(Firmicutes)相對豐度較CK均有所降低,以C3處理最低,分別為17.56%和11.76%。(圖3-A)。在屬水平上,共有11個優勢細菌屬(相對豐度>2%),其中相對豐度排名前三的細菌屬分別為norank_f__norank_o__Actinomarinales、unclassified_f__Intrasporangiaceae和沉積巖桿菌屬(Ilumatobacte)。相較于CK,生物炭處理中的norank_f__norank_o__Actinomarinale相對豐度下降了0.03~0.58個百分點,unclassified_f__Intrasporangiaceae相對豐度增加了0.29~0.59個百分點,以C4處理相對豐度最大(圖3-B)。
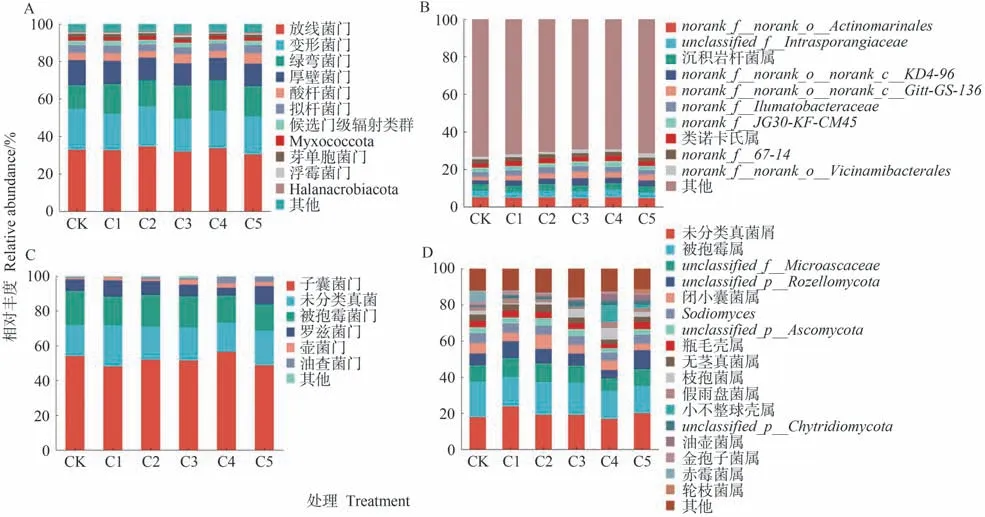
圖3 生物炭對蚯蚓原位堆肥細菌和真菌群落組成的影響Fig.3 Effect of biochar on the relative abundance of microorganisms of in-situ vermicomposting at the phylum and genus level
在真菌菌落中,不同處理檢測出7個相對豐度>1%的真菌菌門,其中優勢菌門為子囊菌門(Ascomycota)、被孢霉菌門(Mortierellomycota)和羅茲菌門(Rozellomycota)。各處理中子囊菌門(Ascomycota)相對豐度為48.05%~56.43%。相較CK,生物炭的添加使被孢霉菌門(Mortierellomycota)相對豐度減少。除C4處理外,其他處理的羅茲菌門(Rozellomycota)相對豐度均高于CK。此外,隨著生物炭添加比例的增加,油壺菌門(Olpidiomycota)作為潛在的優勢菌門,其相對豐度在C4和C5處理中均高于CK(圖3-C)。在屬水平上,共有18個優勢真菌屬(相對豐度>2%),其中相對豐度排名前三的真菌屬為未分類菌屬(unclassified_k__Fung)、被孢霉屬(Mortierella)和羅茲菌屬(Rozellomycota)。相較CK,生物炭處理降低了被孢霉屬(Mortierella)相對豐度,降幅為1.4~4.09個百分點。除C4處理外,其他處理的未分類真菌屬(unclassified_k__Fung)和羅茲菌屬(Rozellomycota)相對豐度較均高于CK(圖3-D)。
2.5 相關指標的冗余分析
將細菌和真菌門、屬水平相對豐度前10的種群與基質理化性質進行RDA分析。結果表明(圖4),基質理化性質對細菌群落豐度解釋度為40.23%(門)和32.58%(屬),真菌群落豐度解釋度為60.54%(門)和40.39%(屬)。
在門水平上,影響細菌群落相對豐度的主要環境因子是全磷(TP)、速效鉀(AK)和速效磷(AP)。其中,放線菌門(Actinobacteria)、綠彎菌門(Chloroflexi)相對豐度與TP、AP、AK含量呈正相關,變形菌門(Proteobacteria)相對豐度與TP、AP、AK含量呈負相關(圖4-A)。影響屬水平細菌群落相對豐度的主要環境因子是全氮(TN)、速效鉀(AK)和全磷(TP)。沉積巖桿菌屬(Ilumatobacte)、unclassified_f__Intrasporangiace相對豐度與AK含量呈正相關,norank_f__norank_o__Vicinamibacterales相對豐度與AK含量呈負相關(圖4-B)。
對門水平的真菌群落變化產生關鍵作用的環境因子為全氮(TN)、速效氮(AN)和速效鉀(AK)。其中,子囊菌門(Ascomycota)相對豐度與TN、AN和AK含量呈正相關,被孢霉菌門(Mortierellomycota)、羅茲菌門(Rozellomycota)相對豐度與AK、AP含量呈負相關(圖4-C)。影響屬水平真菌群落相對豐度的主要環境因子是全氮(TN)、速效氮(AN)和速效鉀(AK)。枝孢菌屬(Cladosporiu)相對豐度與TN含量呈正相關,瓶毛殼屬(Lophotrichus)相對豐度與TN含量呈負相關,unclassified_p__Rozellomycota相對豐度與AK含量呈負相關(圖4-D)。
2.6 蚯蚓原位堆肥與不同比例生物炭對番茄品質和產量的影響
由表5可知,生物炭處理整體顯著增加了番茄植株地下部生物量(P<0.05)。與CK相比,C1~C5處理植株地下部生物量整體顯著增加16.84%~112.19%。隨著生物炭添加比例的增大,番茄植株地下部生物量呈先增大后減小趨勢。其中,C2處理番茄植株地下部生物量達到最大值。同時,生物炭處理整體顯著提高了番茄的產量(P<0.05)。與CK相比,C1~C5處理番茄產量增加11.89%~36.36%,以C3處理產量最高(9 411.72 kg·667m-2)。
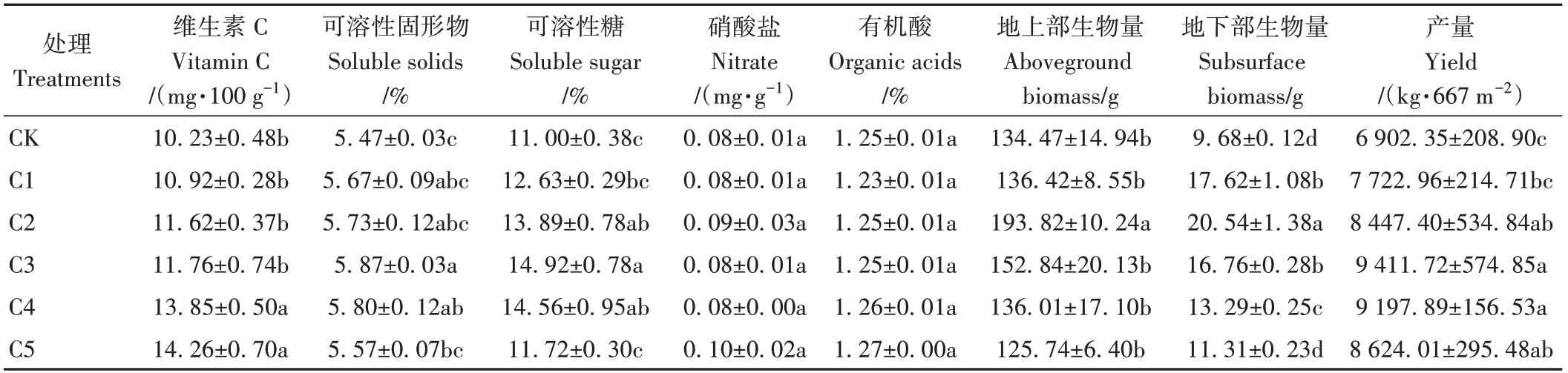
表5 蚯蚓原位堆肥與不同比例生物炭對番茄品質及產量的影響Table 5 Effect of in-situ vermicomposting with different ratio of biochar on quality and yield of tomatoes
添加生物炭均可促進果實品質相關參數的增加。與CK相比,C1~C5處理Vc、可溶性固形物和可溶性糖含量分別增加6.74%~39.39%、0.10~0.40個百分點和0.72~3.92個百分點。隨著生物炭添加比例的增大,番茄Vc含量呈持續上升趨勢;可溶性固形物和可溶性糖含量呈先增大后減小趨勢,二者含量均以C3處理最高,分別為5.87%和14.92%。
2.7 蚯蚓原位堆肥與不同比例生物炭對番茄果實代謝的影響
為了更好地了解蚯蚓原位堆肥與生物炭對番茄果實代謝物的影響,在CK和C3處理之間進行了代謝物分析,通過偏最小二乘法判別分析(partial least squares discriminant analysis,PLS-DA)圖來評估植物組織樣品之間的差異,結果顯示(圖5),PC1的貢獻率為30.70%,PC2的貢獻率為29.50%。從圖中易看出C3與CK間分離趨勢明顯,且因變量擬合指數(R2Y)>0.9,模型預測指數(Q2)>0.5,表明處理之間存在顯著差異,模型具備較好的穩定性,獲得的數據可繼續進行后續分析。此外,潛在結構判別分析(orthogonal partial least squares discriminant analysis,OPLS-DA)建模的監督正交投影也揭示了兩樣本的顯著差異。
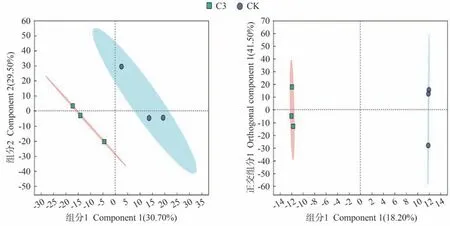
圖5 番茄果實的LC-MS評價(n=3)Fig.5 Evaluation of LC-MS of tomato fruits (n=3)
由圖6可知,通過變量重要性投影(variable importance in projection,VIP)≥1、P值<0.05、差異倍數(fold change,FC)>1或<1的篩選標準,在C3 vs CK中顯著鑒定了16種差異代謝物,其中有14種差異代謝物上調(fold change>1),2種差異代謝物下調(fold change≤1)。由表6可知,相較于CK,C3處理中上調物質包括1-己醇阿拉伯糖基葡糖苷、L-4-羥基谷氨酸半醛、4-羥基雙苯喹啉、5-甲氧基水楊酸等,下調物質包括反式祖馬林和L-谷氨酰胺。
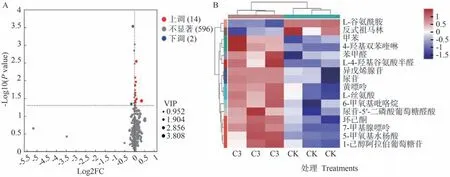
圖6 番茄果實差異代謝物火山圖(A)和熱圖(B)代謝途徑分析Fig.6 Volcano map (A) and heat map (B) of tomato fruit differential metabolites
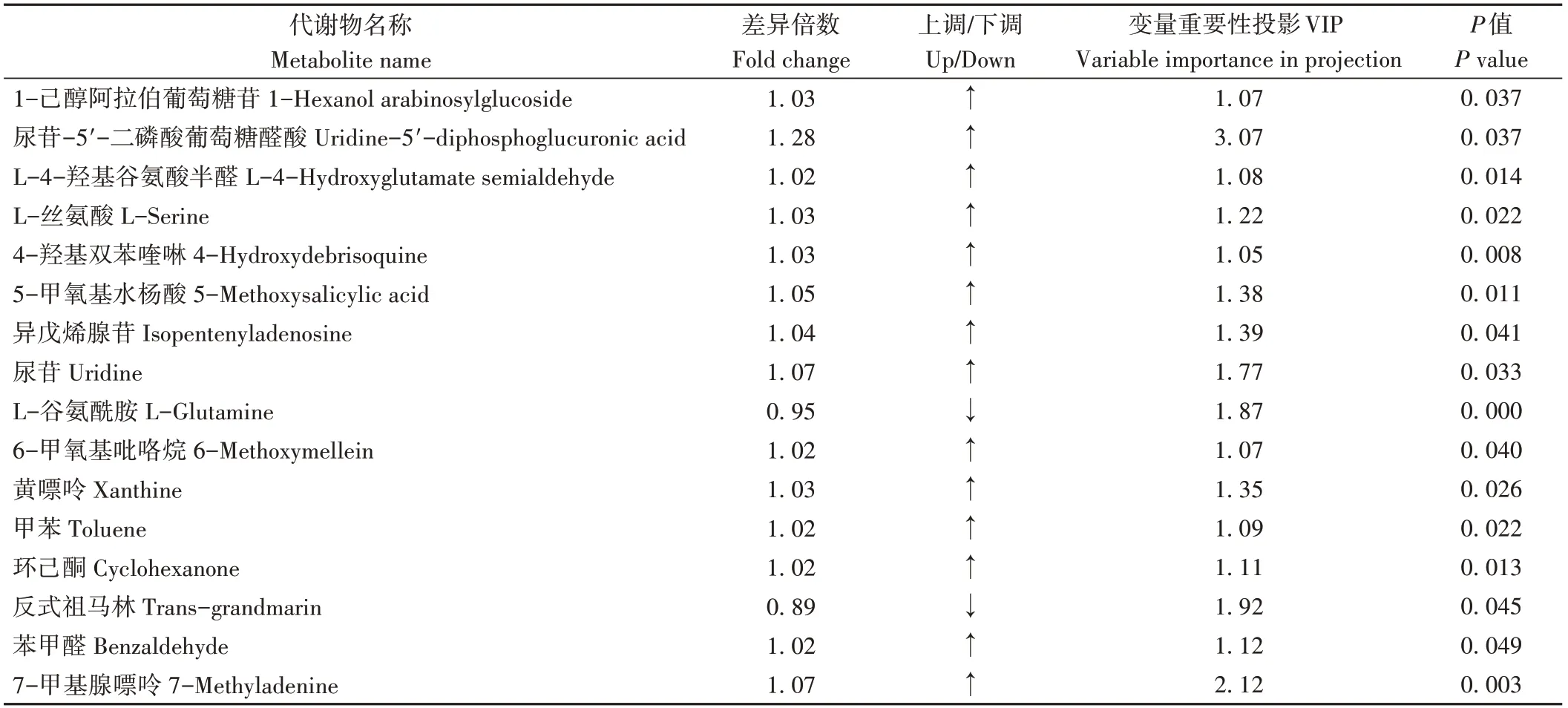
表6 番茄果實差異代謝物Table 6 Differential metabolites of tomato fruit
為進一步了解代謝途徑中差異代謝物的變化,將篩選出的差異代謝物映射到KEGG通路上進行注釋和通路富集分析。由圖7可知,生物炭的施用對氨酰tRNA生物合成、ABC轉運體蛋白(ATP-binding cassette)、乙醛酸和二羧酸代謝、嘌呤代謝和嘧啶代謝等代謝途徑有極顯著影響(P<0.01)。由表7可知,參與乙醛酸和二羧酸代謝、氨酰tRNA生物合成途徑的差異代謝物為L-絲氨酸和L-谷氨酰胺;參與ABC轉運蛋白途徑的差異代謝物為L-絲氨酸、L-谷氨酰胺和尿苷;參與嘌呤代謝途徑的差異代謝物為L-谷氨酰胺和黃嘌呤;參與嘧啶代謝途徑的差異代謝物為尿苷。
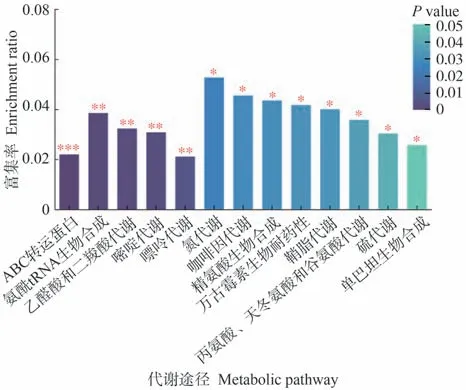
圖7 番茄果實差異代謝物途徑Fig.7 Differential metabolite pathways in tomato fruits
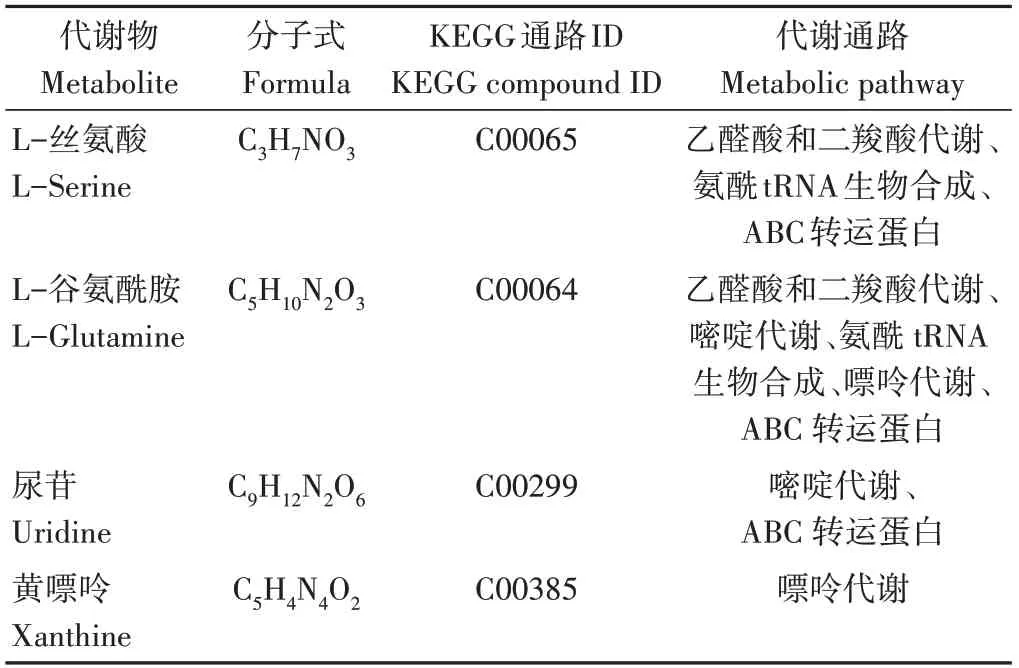
表7 差異代謝物的代謝途徑歸屬Table 7 Metabolic pathway attribution of differential metabolites
3 討論
研究表明,容重在0.1~0.8 g·cm-3、總孔隙度在54%~96%、氣水比在0.25~0.50范圍內的基質最適宜植物生長[20]。本研究所有處理的容重和總孔隙度均滿足上述要求。生物炭的施入整體顯著提高了蚯蚓原位堆肥中的有機質和養分含量,原因可能是生物炭較高的比表面積和吸附能力能固持蚯蚓原位堆肥中的氮、磷、鉀離子,減少養分的流失,增加其可利用養分的總量,且生物炭能夠延緩養分的釋放,使肥效變緩,補充作物后期生長發育所需的養分[21-23]。但隨著生物炭施用量的增加,蚯蚓原位堆肥養分含量呈先上升后下降的趨勢,并且酶活性的變化趨勢與之相似,其潛在原因是過量生物炭會吸附更多酶分子,抑制酶促反應進行,導致酶活性降低[24-25]。
適量生物炭對蚯蚓原位堆肥中的細菌豐富度有顯著影響,且細菌α多樣性指數大于真菌,這歸因于生物炭結構疏松多孔,自身呈弱堿性,配施蚯蚓原位堆肥能夠刺激微生物的代謝,促進細菌群落多樣性的提升,這與程揚等[26]的研究結果相吻合。赤子愛勝蚓腸道內的細菌類群主要為放線菌門、綠彎菌門和厚壁菌門等,上述菌群也被認為是蚯蚓堆肥中常見的優勢微生物[27-28]。這與本研究的結論相似,但各處理間菌群的相對豐度存在差異。一方面,綠彎菌門是一種兼性厭氧菌,可通過反向三羧酸循環來固定碳、氧化硫化物和Fe2+等物質,促進土壤養分循環和碳固定[29],添加生物炭提高了綠彎菌門的相對豐度,這表明生物炭能增加蚯蚓原位堆肥的碳匯,提升有機碳的固持能力。另一方面,生物炭的添加降低了變形菌門和厚壁菌門的相對豐度。變形菌門作為細菌中最大的門類,包括如T.Escherich 1885、沙門氏菌等在內的諸多病原菌,施加生物炭會抑制變形菌門活性,進而降低相關病原菌的相對豐度[30]。此外,Cleveland等[31]發現土壤穩定碳含量的增加會導致γ-變形菌門和厚壁菌門相對豐度的減弱。真菌類群中的優勢菌門為子囊菌門,其次是被孢霉菌門和羅茲菌門。子囊菌門的主要功能是參與土壤有機質分解,尤其具有分解木質纖維素的能力,因此子囊菌門相對豐度的提高有利于有機質的礦化和養分釋放[32]。被孢霉菌門中含有一些具有溶磷作用的真菌種群,可以溶解土壤中難以利用的磷元素[33]。此外,隨著生物炭施用量的增加,油壺菌門的相對豐度提高,這可能是由于生物炭中某種微量物質的積累刺激了油壺菌門的繁殖,但具體原因有待進一步研究。
蚯蚓原位堆肥中添加生物炭提高了番茄的產量和品質。生物炭可加速蚯蚓原位堆肥的腐殖化過程,生物炭的表面也可以被堆肥中的微生物群落和腐殖質氧化,兩者的協同作用增加了基質中可用營養物質的數量,促進了番茄植株的生長,改善了番茄的產量和品質[34]。然而,隨著生物炭添加比例的增大,番茄產量、可溶性固形物和可溶性糖含量呈先增大后減小趨勢,以C3處理最高。因此,生物炭對作物增產效應存在峰值[35]。此外。本研究通過LC-MS測定番茄果實代謝物,結果表明,當生物炭添加比例為3%時,KEGG通路富集產生了5條差異極顯著代謝途徑(P<0.01),以氨酰tRNA生物合成代謝富集率最高。L-絲氨酸和L-谷氨酰胺參與了此過程,絲氨酸作為生糖氨基酸可參與糖異生途徑轉變為葡萄糖,而L-絲氨酸又是由糖酵解中間體3-磷酸甘油酸酯通過三個催化步驟從頭合成的;谷氨酰胺可經過代謝轉化為三羧酸循環的中間產物α-酮戊二酸,進而實現氨基酸與糖類之間的轉化。此外,本研究還確定了尿苷-5'-二磷酸葡萄糖醛酸、1-己醇阿拉伯葡萄糖苷等差異代謝物。尿苷-5'-二磷酸葡萄糖醛酸可在UGlc AE酶的作用下轉化生成尿苷二磷酸半乳糖醛酸,進入高爾基體內參與果膠的生物合成。同時,它還可生成尿苷二磷酸木糖,間接形成木酮糖[36]。這些物質的轉化可能是C3處理糖類物質含量顯著高于CK的主要原因(表5)。綜上,生物炭的施用可能促進了果實中氨基酸與糖類之間的轉化,從而提高了果實品質。
4 結論
本研究結果表明,生物炭可提高蚯蚓原位堆肥中的養分含量和酶活性,優化蚯蚓堆肥菌群結構,進而達到改善番茄品質、提高番茄產量的目標。其中,當生物炭替代比例為3%時,蚯蚓原位堆肥中的全磷、速效磷、速效鉀含量及蔗糖酶和堿性磷酸酶活性最高。上述營養狀況的改變增加了細菌群落的豐富度和多樣性,促使綠彎菌門(Chloroflexi)相對豐度提高,變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)和被孢霉門(Mortierellomycota)相對豐度降低。此外,LC-MS分析發現,生物炭通過改變番茄果實尿苷-5'-二磷酸葡萄糖醛酸、L-絲氨酸、L-谷氨酰胺等的代謝水平而影響果實中糖類物質含量,從而保證其品質與產量保持在高水平。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
