豬CD1d蛋白多克隆抗體的制備及應用
2024-04-24 02:21:18劉傳霞陳欣王曉李雪雯李婷婷翁長江鄭君
中國農業科學 2024年8期
關鍵詞:檢測
劉傳霞,陳欣,王曉,李雪雯,李婷婷,翁長江,鄭君
豬CD1d蛋白多克隆抗體的制備及應用
劉傳霞,陳欣,王曉,李雪雯,李婷婷,翁長江,鄭君
中國農業科學院哈爾濱獸醫研究所/動物疫病預防控制國家重點實驗室/國家非洲豬瘟專業實驗室/基礎免疫創新團隊,哈爾濱 150069
【目的】制備豬源CD1d的多克隆抗體,為探究豬CD1d蛋白在非洲豬瘟病毒(African swine fever virus,ASFV)感染過程中的功能奠定基礎。【方法】利用PCR方法擴增了豬CD1d基因,并將其同源重組至pGEX-6p1載體中,構建了原核重組表達質粒pGEX-6p1-CD1d。將重組質粒轉化BL21(DE3)并用IPTG進行誘導表達,表達的GST-CD1d重組蛋白經SDS-PAGE和Western blot(WB)方法鑒定,SDS-PAGE結果顯示約50 ku處有一條明顯的條帶,該蛋白以包涵體形式表達。然后利用谷胱甘肽瓊脂糖親和層析方法進行蛋白純化,將純化的GST-CD1d蛋白與等體積的弗氏完全佐劑混合乳化后,將純化的蛋白免疫新西蘭大白兔,采用頸背部多點皮下注射,免疫劑量為200 μg/只,首免后第3周和第5周分別進行二免和三免,均采用弗氏不完全佐劑乳化,方法和劑量與首免相同。三免后第7天,通過耳緣靜脈采血分離血清。首免后第7周進行第四次免疫,一周后心臟采血。該抗體經Protein G親和層析純化后凍存于-80 ℃冰箱。通過WB和間接免疫熒光(IFA)鑒定瞬時轉染表達的外源CD1d蛋白和豬原代巨噬細胞(PAMs)表達的內源CD1d蛋白的表達與細胞定位情況。同樣,制備的CD1d抗體可以將外源瞬時轉染表達的CD1d通過IP拉下。為了探究CD1d在ASFV感染早期的情況,將ASFV接種于PAMs,制備ASFV感染0、15、30、60 min的樣品,以CD1d為一抗,通過WB檢測了CD1d蛋白的表達情況。在HEK293T細胞共轉pCAGGS-HA-CD1d和pCAGGS-Flag-CD2v質粒,24 h后收取細胞裂解,加入Flag beads過夜結合蛋白,通過WB檢測互作情況染,同時,質粒共轉染于共聚焦小皿中的HEK293T細胞,用標簽抗體對其進行孵育,選擇相應的熒光二抗,通過激光共聚焦顯微鏡觀察CD1d與CD2v在細胞中共定位情況。采用Co-IP驗證CD1d與ASFV外囊膜蛋白CD2v的相互作用。【結果】原核表達的GST-CD1d蛋白以包涵體形式表達在感受態細胞中,分子質量約為35 ku;實驗兔4次免疫CD1d重組蛋白后采血并分離血清,純化的抗體經SDS-PAGE檢測在45和25 ku處各出現一條特異性條帶,分別為CD1d抗體的重鏈與輕鏈。以純化的CD1d蛋白作為免疫原制備的兔抗CD1d抗體包含重鏈和輕鏈,且具有較好純度;該抗體能夠通過WB和IFA鑒定瞬時轉染的外源以及PAMs內源CD1d蛋白的表達和細胞定位。進一步檢測結果顯示,ASFV感染PAMs后,CD1d蛋白表達水平明顯增加,并且WB和IFA結果顯示CD1d與ASFV編碼的外囊膜蛋白CD2v存在相互作用和共定位。【結論】通過原核表達技術制備了CD1d的抗體,為進一步探究CD1d蛋白在ASFV感染過程中的生物學功能打下了基礎。
CD1d蛋白;原核表達;多克隆抗體;非洲豬瘟病毒;CD2v蛋白
0 引言
【研究意義】非洲豬瘟病毒(African swine fever virus,ASFV)是危害全球生豬產業的“頭號殺手”之一,目前還沒有商品化的藥物或疫苗能夠控制該病毒的傳播,其中一個主要原因是人們對其感染與免疫逃逸機制認知不足[1]。【前人研究進展】ASFV病毒顆粒由核酸、核殼、內膜和衣殼4個同心結構組成[2],成熟的病毒粒子在通過細胞質膜出芽時獲得病毒的外囊膜,由ASFV EP402R基因編碼的CD2v蛋白是已知的病毒外囊膜上的唯一標記分子[2-4]。CD2v基因全長為1 083 bp,預測的蛋白質大小為41 kDa,由402個氨基酸構成,是一個高度糖基化的囊膜蛋白[4-5]。因編碼蛋白的胞外區氨基酸序列與宿主細胞的CD2非常相似,所以命名為CD2v,該蛋白在促進病毒復制與傳播、病毒免疫逃逸等方面發揮著重要作用[2]。CD2v也具有良好的免疫原性,可應用在減毒活疫苗、亞單位疫苗等研究中。有研究顯示泛素化的CD2v蛋白很可能參與了MHC-I類分子抗原遞呈途徑,從而激活特異性T細胞免疫[6]。MINMA等[7]的研究使用計算機模擬方法鑒定CD2v 蛋白胞外區包含4個B細胞表位和5個T細胞表位。BURMAKINA等[8]的研究鑒定結果顯示了4個T細胞表位能夠出現T細胞應答。CD1d分子是CD1家族的成員之一,是一種與MHC-I類分子類似的蛋白[9-10],主要來源于單核細胞、樹突狀細胞及巨噬細胞等。CD1d的分子結構包含胞外功能區(a1、a2、a3區)、跨膜區和胞內區。其中a1和a2區構成抗原結合域,它是由疏水性氨基酸組成的利于糖脂類分子與之結合的空穴樣結構[11]。CD1d分子的組成包括非糖基化和糖基化兩種,后者是由β2微球蛋白(β2m)與CD1d分子以非共價鍵形成的二聚體。CD1d能夠識別并遞呈糖脂類抗原給T細胞,通過特異性激活自然殺傷T(NKT)細胞,發揮抗病毒感染的免疫功能[12]。CD1d分子與β2-微球蛋白結合后,通過內質網-高爾基體的分泌途徑到達質膜,進而特異性地將抗原提呈給NKT細胞,使其活化并分泌多種細胞因子,直接或間接參與機體免疫應答[13]。CD1d分子提呈糖脂抗原[14],在抗原裝載、胞內運輸和其加工處理等方面,具有獨特特征,從而在抗腫瘤、抗感染和抑制自身免疫中發揮重要作用[15-17]。CD1d主要表達在抗原提呈細胞等細胞表面,髓系細胞通常低表達CD1d,這有助于避免激活自身反應性NKT細胞。自身免疫病患者外周血中CD1d的表達呈上調狀態,促進了對機體自身糖脂類抗原的提呈,從而導致了病理性的免疫應答。類風濕性關節炎患者體內檢測出低水平的可溶性CD1d[18],而正常人體的CD1d分子是以跨膜蛋白的形式存在,可溶性CD1d在自身免疫性疾病中作用尚不清楚,有待進一步揭示。【本研究切入點】柯薩奇病毒感染心肌細胞后,CD1d出現明顯表達升高的現象[19]。HBV或HCV感染者的肝臟組織中CD1d表達出現上調的現象[20]。這些都提示了CD1d能夠介導產生抗病毒感染的作用。【擬解決的關鍵問題】由于商品化的其他物種CD1d抗體不能很好識別豬CD1d蛋白,本研究利用原核表達系統制備了豬源CD1d重組蛋白,通過免疫新西蘭大白兔制備了CD1d多克隆抗體,進而將其應用于WB、IFA和Co-IP等試驗,鑒定了ASFV感染后CD1d蛋白的表達變化,驗證了CD1d與CD2v蛋白的相互作用,為研究CD1d在ASFV感染過程中的生物學功能以及抗原提呈機制奠定了基礎。
1 材料與方法
1.1 試驗材料及實驗動物
HEK293T細胞由哈爾濱獸醫研究所基礎免疫團隊保存,豬肺泡巨噬細胞(PAMs)分離自30日齡ASFV陰性豬;DH5ɑ菌株、BL21(DE3)感受態細胞購自TaKaRa公司;真核表達質粒pCAGGS- HA-CD1d、pCAGGS-Flag-CD1d由哈爾濱獸醫研究所基礎免疫團隊制備并保存[21];SPF級2月齡雌性新西蘭大白兔購自遼寧長生生物技術股份有限公司。
1.2 主要試劑
Prime STAR Max DNA Polymerase 購自寶生物工程有限公司;限制性內切酶HⅠ、I購自NEB公司;同源重組試劑盒購自Vazyme公司;DMEM培養基、胎牛血清購自Gibco公司;質粒小提試劑盒、膠回收試劑盒均購自QIAGEN公司;弗氏完全佐劑和弗氏不完全劑購自Sigma公司;兔抗GST單克隆抗體(MAb)由哈爾濱獸醫研究所基礎免疫團隊制備并制備;兔源HA標簽MAb購自Zen Bioscience公司;兔源Flag標簽MAb購自Sigma- Aldrich公司;紅外熒光標記的山羊抗兔和抗鼠IgG購自LICOR公司。
1.3 重組質粒的構建
利用TMHMM軟件分析GenBank中登錄的豬CD1d蛋白的氨基酸序列,預測其跨膜區(267aa— 289aa),針對該區對應cDNA序列設計引物CD1d-F:5'-TCCAGGGGCCCCTATGCTGCATTCCCTTCCA GC-3'(下劃線為HⅠ酶切位點)CD1d-R: 5'-GTCAGTCACGATTCAGATGTCCTGGTCTCCTA-3'(下劃線為Ⅰ酶切位點),由庫美生物科技有限公司(吉林)合成,預期PCR產物大小為834 bp。由吉林省庫美生物科技有限公司合成。
1.4 重組蛋白的表達與純化
以pCAGGS-HA-CD1d為模板,CD1d-F/CD1d-R為引物,利用PCR方法擴增CD1d對應的cDNA片段,PCR擴增體系50 μL:PrimeSTARMax Premix(2×)25 μL、上下游引物各1 μL、DNA模板50 ng,ddH2O補至50 μL。PCR反應條件:98 ℃預變性2 min;98 ℃變性10 s,55 ℃退火15 s,72 ℃延伸20 s,共35個循環;72 ℃延伸5 min;同時,pEGX-6p1空載體經H I和I雙酶切,將PCR產物和酶切產物經瓊脂糖凝膠電泳鑒定后,用膠回收試劑盒純化回收,然后二者產物經同源重組酶連接。連接產物轉化感受態DH5α,并涂布于氨芐抗性LB平板,37 ℃倒置過夜培養12 h。菌液PCR鑒定正確后,送至吉林省庫美生物科技有限公司測序。測序正確的重組質粒按照小提質粒試劑盒說明書提取質粒,命名為pGEX-6p1-CD1d,-20 ℃保存。正確的重組質粒命名為 pGEX-6p1- CD1d。將重組質粒pGEX-6p1-CD1d轉化至大腸桿菌BL21(DE3)感受態細胞中,37 ℃ 220 r/min培養至OD600nm值為0.6—0.8,加入終濃度為1mol·L-1的IPTG,16℃220 r/min誘導20 h。收集細菌,超聲破碎后離心,收集上清和沉淀后分別經SDS-PAGE檢測重組蛋白的表達。獲得的重組GST-CD1d蛋白可以與谷胱甘肽S轉移酶融合,因此可以通過谷胱甘肽-瓊脂糖親和層析進行純化,經BCA法測定純化蛋白濃度后置-80℃保存。
1.5 多克隆抗體的制備
將純化的GST-CD1d蛋白與等體積的弗氏完全佐劑混合乳化后,采用頸背部多點皮下注射,免疫劑量為200 μg/只,首免后第3周和第5周分別進行二免和三免,均采用弗氏不完全佐劑乳化,方法和劑量與首免相同。三免后第7天,通過耳緣靜脈采血分離血清。首免后第7周進行第四次免疫,一周后心臟采血,分離血清利用 Protein G在4℃過夜結合后經1 mol·L-1Tris-HCl(pH8.5)洗脫液洗脫純化并經SDS-PAGE鑒定后,進一步以兔抗 GST MAb(1﹕2 000)作為一抗,以IRDye 800CW標記的山羊抗兔IgG(1﹕10 000)作為二抗進行WB鑒定。
1.6 外源CD1d表達和定位的鑒定
將pCAGGS-HA-CD1d轉染至HEK293T細胞中,24 h后收集細胞,用含1% NP-40的細胞裂解液裂解,離心后取上清,經SDS-PAGE電泳后,轉印到PVDF膜,用TBST配制的5%脫脂乳封閉2 h。以制備的CD1d抗體(1﹕2 000)作為一抗,以IRDye 800CW標記的山羊抗兔IgG(1﹕10 000)作為二抗;以轉染空質粒的細胞裂解液作為陰性對照,經WB鑒定CD1d抗體與真核表達CD1d蛋白的反應性,評估CD1d抗體在檢測外源CD1d蛋白表達中的應用效果。然后利用多聚賴氨酸包被的共聚焦培養皿,進而接種HEK293T細胞,將pCAGGS-HA-CD1d質粒轉染HEK293T細胞24 h后,棄上清,固定、透膜后,10% FBS室溫封閉1 h;分別以HA標簽抗體(1﹕1 000)或CD1d抗體(1﹕100)作為一抗;以Alexa Fluor-594山羊抗兔IgG(1﹕2 000)作為二抗,室溫避光孵育1 h;經DAPI染色20 min,IFA檢測CD1d抗體與真核表達CD1d蛋白的反應性,鑒定豬CD1d外源蛋白在HEK293T細胞中的定位。
1.7 內源CD1d表達和定位的鑒定
將PAMs鋪于6孔板中,24 h收集細胞,采用WB方法檢測CD1d抗體與內源表達CD1d蛋白的反應性;以HEK293T細胞裂液作為陰性對照。評估CD1d抗體在檢測內源表達CD1d蛋白中的應用。利用多聚賴氨酸包被的共聚焦培養皿,進而接種PAMs,24 h后棄上清,采用IFA方法檢測CD1d抗體與內源表達CD1d蛋白的特異性反應,鑒定內源性CD1d蛋白的定位情況。
1.8 豬CD1d的抗體在IP試驗中的應用
將pCAGGS-Flag-CD1d真核表達質粒轉染至HEK293T細胞中,同時轉染空質粒至HEK293T細胞作為對照。24 h后收集細胞樣品,裂解細胞后進行IP試驗:在樣品中加入FLAG-Agarose beads,4℃旋轉孵育至少6 h;用預冷的含1% NP-40 細胞裂解液洗滌beads 5次后煮沸,將制備的樣品經SDS-PAGE電泳,轉印到PVDF膜,以制備的CD1d抗體(1﹕1 000)作為一抗,以IRDye 800CW標記的山羊抗兔IgG(1﹕10 000)作為二抗,檢測免疫沉淀中的CD1d蛋白,評估CD1d抗體在IP試驗中的應用效果。
1.9 ASFV感染對豬CD1d表達水平的影響
將PAMs鋪于6孔板中,24 h后接種ASFV(1 MOI),分別于0、15、30、60 min收集細胞并裂解,以不接種ASFV的細胞裂解液作為對照。以制備的CD1d抗體(1﹕1 000)作為一抗,以IRDye 800CW標記的山羊抗兔IgG(1﹕10 000)作為二抗,經WB檢測PAMs中CD1d蛋白的表達水平變化。
1.10 CD1d與CD2v相互作用的鑒定
將pCAGGS-HA-CD1d和pCAGGS-Flag-CD2v各2 μg共轉染共聚焦培養皿和六孔板中的HEK293T細胞,對照細胞不作任何處理。24 h后棄去培養基,4%多聚甲醛室溫固定共聚焦培養皿中的細胞30 min,分別以CD1d抗體(1﹕500)、HA和Flag標簽抗體(1﹕500)為一抗,以Alexa Fluor-594 山羊抗鼠IgG(1﹕2 000)和Alexa Fluor-633山羊抗兔IgG(1﹕2 000)為二抗,通過激光共聚焦顯微鏡觀察CD1d與CD2v的共定位情況;六孔板中的HEK293T細胞,轉染質粒24 h后使用FLAG beads進行Co-IP試驗,以驗證CD1d與CD2v的相互作用。
2 結果
2.1 CD1d蛋白的表達及純化
以pCAGGS-HA-CD1d質粒為模板,PCR擴增出的CD1d片段與預測大小相符(834 bp)。將獲得的豬CD1d片段克隆至pGEX-6p1原核表達載體,獲得的重組表達質粒經限制性內切酶HⅠ和Ⅰ雙酶切鑒定及測序鑒定,結果表明重組質粒pGEX-6p1-CD1d構建正確。將重組質粒轉化BL21(DE3)并誘導表達,SDS-PAGE結果顯示約50 ku處有一條明顯的條帶,該蛋白以包涵體形式表達(圖1-A)。WB結果顯示,此條帶為CD1d蛋白特異性條帶,其大小與預期結果一致,且經過純化的蛋白較為純凈(圖1-B)。以上結果說明得到了純度較高的CD1d重組蛋白,經BCA法測定后的蛋白濃度為58 mg·mL-1。
2.2 豬CD1d多克隆抗體的純化
試驗兔4次免疫CD1d重組蛋白后采血并分離血清,通過Protein G純化的抗體經SDS-PAGE檢測,結果顯示,在45和25 ku處各出現一條特異性條帶,分別為CD1d抗體的重鏈與輕鏈(圖2)。結果表明,豬CD1d多克隆抗體被成功制備和純化。
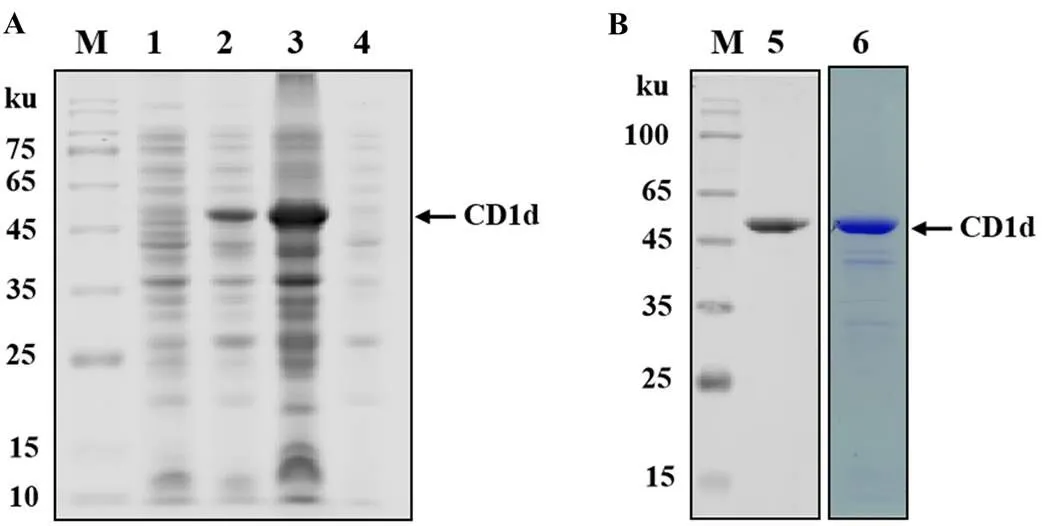
A:SDS-PAGE的鑒定結果;B:WB的鑒定結果。M:蛋白分子質量標準;1:未誘導的菌液;2-4:IPTG誘導的重組菌菌液、超聲后沉淀、超聲后上清;5:WB鑒定純化的CD1d蛋白; 6: SDS-PAGE鑒定純化的CD1d蛋白
2.3 外源CD1d蛋白的表達與定位
將pCAGGS-Flag-CD1d轉染HEK293T細胞,24 h后收集細胞,分別經WB和IFA檢測。WB結果顯示,純化的CD1d抗體可以識別外源表達的Flag-CD1d蛋白,其大小近35 ku,結果中顯示出了兩條特異性條帶,推測可能是由于豬源的Flag-CD1d蛋白在HEK293T細胞中表達存在一定的修飾造成的(圖3-A)。質粒pCAGGS-HA-CD1d轉染的細胞分別用HA標簽抗體及CD1d抗體作為一抗,通過IFA檢測,結果顯示,均可檢測到特異性的熒光,其位于細胞的胞質內(圖3-B)。這些結果說明制備的CD1d抗體可以用于WB及IFA檢測外源表達的豬CD1d蛋白。

M:蛋白質分子質量標準;1-4:純化的不同濃度CD1d抗體

A:WB分析外源CD1d蛋白在HEK293T細胞中的表達;B:IFA鑒定外源CD1d蛋白在HEK293T細胞中的定位。M:蛋白質分子質量標準;1:轉染pCAGGS-HA-CD1d的 HEK293T細胞裂解液;2:轉染pCAGGS-HA的HEK293T細胞裂解液
2.4 內源CD1d蛋白的表達與定位
鋪于6孔板中的PAMs裂解后分別經WB和IFA鑒定。WB結果顯示,制備的CD1d抗體能夠與PAMs中的內源CD1d蛋白反應,產生一條近55 ku的特異性條帶(圖4-A);以CD1d抗體作為一抗,IFA結果顯示,PAMs中出現特異性紅色熒光,位于細胞膜和細胞質,而對照無熒光(圖4-B),表明CD1d定位于PAMs的胞質內。這些結果說明制備的抗體可以用于檢測內源表達的豬CD1d蛋白。
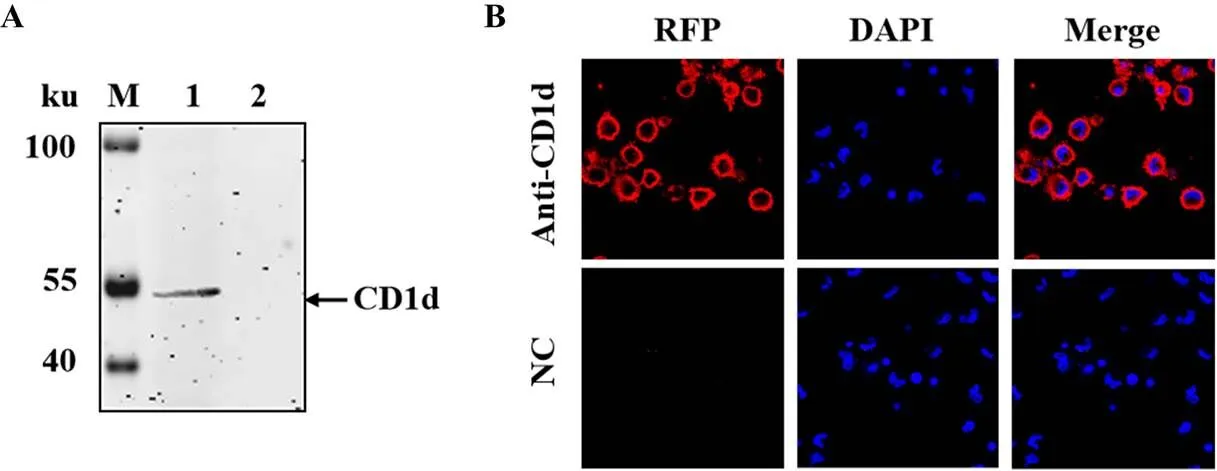
A:WB分析PAMs中CD1d的表達;B:IFA鑒定CD1d在PAMs中的定位。M:蛋白質分子質量標準;1:PAMs裂解液;2:MA104細胞裂解液
2.5 豬CD1d抗體在IP試驗中的應用
將pCAGGS-Flag-CD1d轉染至HEK293T細胞中,24 h后收集細胞樣品,經IP試驗檢測,結果顯示,CD1d抗體可以將Flag-CD1d蛋白沉淀下來(圖5)。
2.6 ASFV感染對豬CD1d表達水平的影響
將ASFV(1 MOI)接種6孔板中的PAMs,分別在不同時間收集并裂解細胞,通過WB檢測病毒感染不同時間PAMs中CD1d的表達水平。結果顯示,隨著ASFV感染時間的增加,CD1d蛋白的表達水平逐漸升高(圖6-A),通過Image J軟件對圖6-A的結果進行灰度值分析,結果顯示,在ASFV感染60 min時CD1d的表達水平增加了近3倍(圖6-B)。結果表明,ASFV感染促進了PAMs中CD1d蛋白的表達水平。
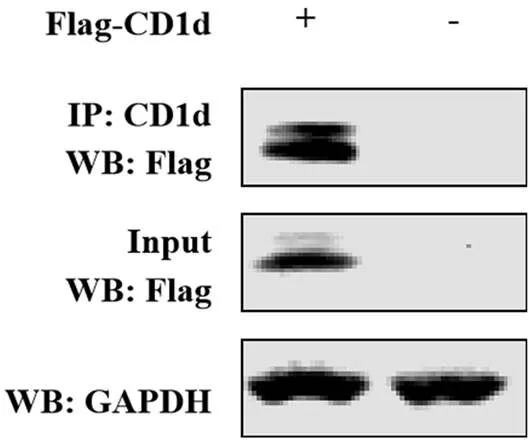
圖5 CD1d抗體用于IP試驗
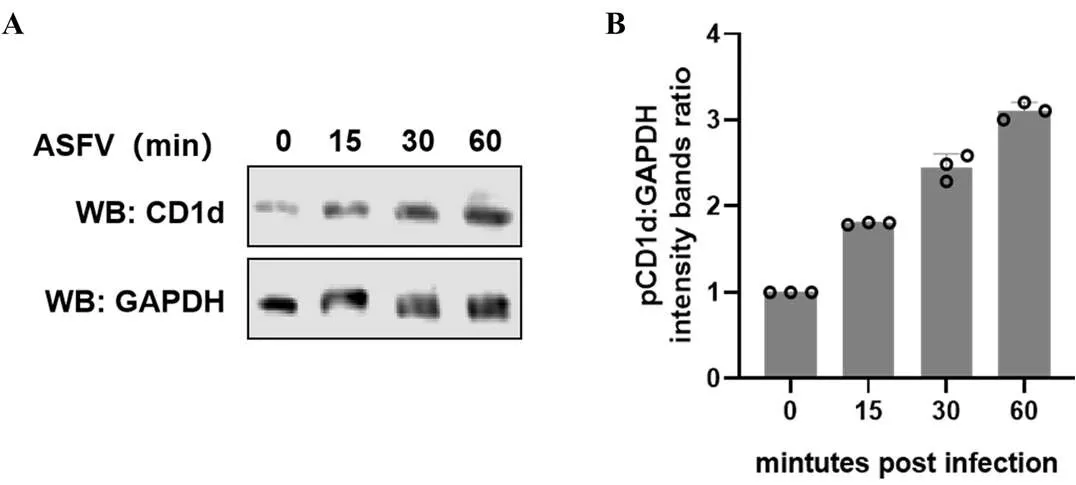
A:WB分析ASFV感染不同時間CD1d的表達情況;B:對A圖的灰度值進行比較分析。縱坐標顯示的是CD1d與GAPDH的灰度比值
2.7 CD1d與CD2v相互作用的鑒定
將pCAGGS-Flag-pCD2v與pCAGGS-HA-CD1d轉染HEK293T細胞,24 h后分別利用CD1d和HA標簽抗體作為一抗,相應熒光二抗孵育后經激光共聚焦顯微鏡下觀察(圖7-A);將CD1d和CD2v質粒轉染HEK293T細胞中,24 h后收集細胞,樣品經IP試驗檢測與相互作用(圖7-B)。結果顯示,宿主蛋白CD1d與非洲豬瘟外囊膜蛋白CD2v存在相互作用。

A:IFA鑒定CD1d與CD2v的共定位;B:Co-IP鑒定CD1d與CD2v的相互作用
3 討論
3.1 非洲豬瘟的危害
非洲豬瘟(African swine fever virus,ASF)是由ASFV感染家豬和野豬而引起的急性、烈性、高度接觸性傳染病。對中國甚至全世界造成嚴重的經濟損失。非洲豬瘟的傳播方式眾多, 發病豬及感染豬的排泄物、分泌物、豬肉及其制品以及污染的運輸車輛、飼料、人員、衣物、鞋子等均為重要的傳染源, 特別是感染豬的調運會加速疫情的傳播[22]。在ASFV感染的巨噬細胞或其他細胞系中表達了超過150種病毒蛋白,這些蛋白不僅參與病毒進入、復制、病毒粒子組裝和排出,還調節宿主抗病毒先天免疫應答[23-28]。由于ASFV病毒粒子的結構和逃避先天免疫應答的復雜機制,目前還沒有現有的治療藥物或有效的疫苗。
3.2 CD1d蛋白的結構與表達形式
CD1家族蛋白為非經典的MHC-I類分子,主要與脂類或糖脂類抗原形成復合物,從而將抗原提呈給NKT細胞。CD1d與抗原結合的結構域主要位于其胞外功能區頂端的a1和a2區,它是由疏水性氨基酸組成的利于脂類分子與之結合的空穴樣結構,這便導致CD1d的疏水性較強,因此在通過原核表達CD1d蛋白時呈現出不可溶的包涵體形式。然而其抗原表位仍有很大幾率暴露在外側,通過免疫兔子制備的抗體也呈現出較好的反應原性,說明通過包涵體來制備豬CD1d的特異性抗體切實可行。
3.3 CD1d參與多種病毒感染過程
CD1d 的表達對于激活NKT 細胞是必要的,這一表達在病原感染時被調節。HCV在感染時,肝臟的CD1d表達明顯增多,進而被NKT細胞識別其遞呈的脂質抗原,從而正向或負向調節炎癥應答反應[15]。巨細胞病毒(MCMV)或淋巴細胞脈絡叢腦膜炎病毒(LCMV)感染小鼠時,控制NKT細胞的激活和CD1d抗原提呈,結果增強了對病毒的清除作用[16]。牛痘病毒(VV)或水皰性口炎病毒(VSV)感染后,CD1d分子被分離到細胞的一邊,這也伴隨著細胞信號通路的改變[17]。單純皰疹病毒-1(HSV-1)改變了人類CD1d的黏附力,致使CD1d介導的抗原提呈受到損傷,進而抑制了NKT細胞的活化[18]。
3.4 CD1d與ASFV感染相關聯
早期對CD1d的研究主要集中認為CD1d只能識別脂類抗原。但隨著對α-半乳糖基神經酰胺研究的深入,發現CD1d分子還能夠特異性結合外源性糖脂類抗原[29]。但糖脂類抗原在未經CD1d 加工處理不能被NKT 細胞所識別[30]。本研究結果顯示,CD1d蛋白存在于細胞膜和細胞質中,在ASFV感染初期,CD1d的表達水平呈現逐漸增高的趨勢。此外,豬CD1d能夠與ASFV編碼的外囊膜蛋白CD2v相互作用,這些結果說明CD1d可能與介導病毒入侵相關,也可能是細胞的免疫調節反應,從而促進CD1d的病毒抗原提呈作用。這種結合是否符合CD1d的抗原裝載過程,又是否能夠提呈CD2v抗原還有待進一步鑒定。
4 結論
本研究利用原核表達的豬CD1d蛋白成功制備了特異性多克隆抗體,該抗體能夠有效識別內源性的豬CD1d抗原。通過使用該抗體,發現ASFV感染能夠促進CD1d的表達水平,且CD1d與CD2v存在相互作用,這為進一步鑒定CD1d在ASFV感染過程中的功能研究打下了基礎。
[1] DIXON L K, SUN H, ROBERTS H. African swine fever. Antiviral Research, 2019, 165: 34-41.
[2] ALEJO A, MATAMOROS T, GUERRA M, ANDRéS G. A proteomic atlas of the African swine fever virus particle. Journal of Virology, 2018, 92(23): e01293-18.
[3] WANG G G, XIE M J, WU W, CHEN Z Z. Structures and functional diversities of ASFV proteins. Viruses, 2021, 13(11): 2124.
[4] WANG N, ZHAO D M, WANG J L, ZHANG Y L, WANG M, GAO Y, LI F, WANG J F, BU Z G, RAO Z H, WANG X X. Architecture of African swine fever virus and implications for viral assembly. Science, 2019, 366(6465): 640-644.
[5] DIXON L K, CHAPMAN D A G, NETHERTON C L, UPTON C. African swine fever virus replication and genomics. Virus Research, 2013, 173(1): 3-14.
[6] 王曼, 沈宇清. 非洲豬瘟病毒結構蛋白CD2v的功能研究進展. 中國免疫學雜志, 2021, 37(22): 2734-2737, 2744.
WANG M, SHEN Y Q. Research progress in function of ASFV structural protein CD2v. Chinese Journal of Immunology, 2021, 37(22): 2734-2737, 2744. (in Chinese)
[7] MINMA K A, KATORKINA E I, KATORKIN S A, TSYBANOV S Z, MALOGOLOVKIN A S. In silico prediction of B- and T-cell epitopes in the CD2v protein of African swine fever virus (African Swine Fever Virus, Asfivirus, Asfarviridae). Problems of Virology, 2020, 65(2): 103-112.
[8] BURMAKINA G, MALOGOLOVKIN A, TULMAN E R, XU W D, DELHON G, KOLBASOV D, ROCK D L. Identification of T-cell epitopes in African swine fever virus CD2v and C-type lectin proteins. Journal of General Virology, 2019, 100(2): 259-265.
[9] FOWLKES B J, KRUISBEEK A M, TON-THAT H, WESTON M A, COLIGAN J E, SCHWARTZ R H, PARDOLL D M. A novel population of T-cell receptor αβ-bearing thymocytes which predominantly expresses a single Vβ gene family. Nature, 1987, 329(6136): 251-254.
[10] MAKINO Y, KANNO R, ITO T, HIGASHINO K, TANIGUCHI M. Predominant expression of invariant Vα14+TCR α chain in NK1.1+T cell populations. International Immunology, 1995, 7(7): 1157-1161.
[11] GODFREY D I, MACDONALD H R, KRONENBERG M, SMYTH M J, VAN KAER L. NKT cells: what’s in a Name? Nature Reviews Immunology, 2004, 4: 231-237.
[12] 陸田田, 黃震, 陳章權. CD1d分子的結構與功能. 生命的化學, 2008, 28(2): 159-161.
LU T T, HUANG Z, CHEN Z Q. Structure and Function of CD1d Molecule. Chemistry of Life, 2008, 28(2): 159-161. (in Chinese)
[13] KANG S J, CRESSWELL P. Calnexin, calreticulin, and ERp57 cooperate in disulfide bond formation in human CD1d heavy chain. Journal of Biological Chemistry, 2002, 277(47): 44838-44844.
[14] 師義, 王昆華, 劉為軍, 徐玉. CD1d分子研究進展. 廣東醫學, 2012, 33(11): 1678-1680.
SHI Y, WANG K H, LIU W J, XU Y. Research Progress of CD1d Molecular. Guangdong Medical Journal, 2012, 33(11): 1678-1680. (in Chinese)
[15] ARGILAGUET J M, PéREZ-MARTíN E, NOFRARíAS M, GALLARDO C, ACCENSI F, LACASTA A, MORA M, BALLESTER M, GALINDO-CARDIEL I, LóPEZ-SORIA S, ESCRIBANO J M, RECHE P A, RODRíGUEZ F. DNA vaccination partially protects against African swine fever virus lethal challenge in the absence of antibodies. PLoS ONE, 2012, 7(9): e40942.
[16] DURANTE-MANGONI E, WANG R J, SHAULOV A, HE Q, NASSER I, AFDHAL N, KOZIEL M J, EXLEY M A. Hepatic CD1d expression in hepatitis C virus infection and recognition by resident proinflammatory CD1d-reactive T cells. The Journal of Immunology, 2004, 173(3): 2159-2166.
[17] RENUKARADHYA G J, WEBB T J R, KHAN M A, LIN Y L, DU W J, GERVAY-HAGUE J, BRUTKIEWICZ R R. Virus-induced inhibition of CD1d1-mediated antigen presentation: reciprocal regulation by p38 and ERK. The Journal of Immunology, 2005, 175(7): 4301-4308.
[18] WEBB T J, CAREY G B, EAST J E, SUN W J, BOLLINO D R, KIMBALL A S, BRUTKIEWICZ R R. Alterations in cellular metabolism modulate CD1d-mediated NKT-cell responses. Pathogens and Disease, 2016, 74(6): ftw055.
[19] YANG J Q, CHUN T, LIU H Z, HONG S, BUI H, VAN KAER L, WANG C R, SINGH R. CD1d deficiency exacerbates inflammatory dermatitis in MRL-mice. European Journal of Immunology, 2004, 34(6): 1723-1732.
[20] 陳建勇, 沈學文, 張吉翔. CD1d/NKT在抗HBV和HCV中的作用. 生命的化學, 2007, 27(3): 246-248.
CHEN J Y, SHEN X W, ZHANG J X. Protection of CD1d/NKT against HBV and HCV. Chemistry of Life, 2007, 27(3): 246-248. (in Chinese)
[21] CHEN X, ZHENG J, LIU C X, LI T T, WANG X, LI X W, BAO M F, LI J N, HUANG L, ZHANG Z X, BU Z G, WENG C J. CD1d facilitates African swine fever virus entry into the host cells via clathrin-mediated endocytosis. Emerging Microbes & Infections, 2023, 12(2): 2220575.
[22] 羅玉子, 孫元, 王濤, 仇華吉. 非洲豬瘟: 我國養豬業的重大威脅. 中國農業科學, 2018, 51(21): 4177-4187.
LUO Y Z, SUN Y, WANG T, QIU H J. African swine fever: a major threat to the Chinese swine industry. Scientia Agricultura Sinica, 2018, 51(21): 4177-4187. (in Chinese)
[23] MATAMOROS T, ALEJO A, RODRíGUEZ J M, HERNáEZ B, GUERRA M, FRAILE-RAMOS A, ANDRéS G. African swine fever virus protein pE199L mediates virus entry by enabling membrane fusion and core penetration. mBio, 2020, 11(4): e00789-20.
[24] ANDRE?S G, GARC??A-ESCUDERO R, VIN?UELA E, SALAS M L, RODR??GUEZ J M. African swine fever virus structural protein pE120R is essential for virus transport from assembly sites to plasma membrane but not for infectivity. Journal of Virology, 2001, 75(15): 6758-6768.
[25] SáNCHEZ E G, PéREZ-Nú?EZ D, REVILLA Y. Mechanisms of entry and endosomal pathway of African swine fever virus. Vaccines, 2017, 5(4): 42.
[26] CUESTA-GEIJO M á, GARCíA-DORIVAL I, DEL PUERTO A, URQUIZA J, GALINDO I, BARRADO-GIL L, LASALA F, CAYUELA A, SORZANO C O S, GIL C, DELGADO R, ALONSO C. New insights into the role of endosomal proteins for African swine fever virus infection. PLoS Pathogens, 2022, 18(1): e1009784.
[27] HERNAEZ B, ALONSO C. Dynamin- and clathrin-dependent endocytosis in African swine fever virus entry. Journal of Virology, 2010, 84(4): 2100-2109.
[28] LIU J Y, GALLO R M, DUFFY C, BRUTKIEWICZ R R. A VP22-null HSV-1 is impaired in inhibiting CD1d-mediated antigen presentation. Viral Immunology, 2016, 29(7): 409-416.
[29] LEE A, FARRAND K J, DICKGREBER N, HAYMAN C M, JüRS S, HERMANS I F, PAINTER G F. Novel synthesis of α-galactosyl- ceramides and confirmation of their powerful NKT cell agonist activity. Carbohydrate Research, 2006, 341(17): 2785-2798.
[30] PRIGOZY T I, NAIDENKO O, QASBA P, ELEWAUT D, BROSSAY L, KHURANA A, NATORI T, KOEZUKA Y, KULKARNI A, KRONENBERG M. Glycolipid antigen processing for presentation by CD1d molecules. Science, 2001, 291(5504): 664-667.
Preparation and Application of Polyclonal Antibodies Against Pig CD1d Protein
LIU ChuanXia, CHEN Xin, WANG Xiao, LI XueWen, LI TingTing, WENG ChangJiang, ZHENG Jun
Division of Fundamental Immunology, National African Swine Fever Para-Reference Laboratory, State Key Laboratory for Animal Disease Control and Prevention/Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, Harbin 150069
【Objective】 The aim of this study was to prepare polyclonal antibodies against porcine CD1d protein, so as to lay the foundation for exploring the function of porcine CD1d protein in the process of African swine fever virus (ASFV) infection. 【Method】 In this study, the pig CD1d gene was amplified using PCR and homologously recombined into the pGEX-6p1 vector, constructing a prokaryotic recombinant expression plasmid pGEX-6p1-CD1d. The recombinant plasmidBL21 (DE3) was transformed and induced for expression using IPTG. The expressed GST CD1d recombinant protein was identified by SDS-PAGE and Western blot (WB) methods. The SDS-PAGE results showed a clear band at approximately 50 ku, which was expressed in the form of an inclusion body. Then, protein purification was performed using glutathione agarose affinity chromatography. The purified GST-CD1d protein was mixed and emulsified with an equal volume of Freund's complete adjuvant. The purified protein was immunized in New Zealand white rabbits and administered subcutaneously at multiple points on the neck and back, with an immune dose of 200 μG/piece, and then second and third immunizations were performed at the 3rd and 5th weeks after the first immunization, respectively, using Freund's incomplete adjuvant emulsification, with the same method and dosage as the first immunization. On the 7th day after the third immunization, the blood was collected from the ear vein to isolate the serum. The fourth immunization was conducted at the 7 weeks after the first immunization, and the blood was collected from the heart one week later. The antibody was purified by Protein G affinity chromatography and frozen at -80 ℃. The expression and cellular localization of endogenous CD1d protein expressed by transient transfection of exogenous and porcine primary macrophages (PAMs) were indentified by using WB and indirect immunofluorescence (IFA). Similarly, the prepared CD1d antibody could pull down CD1d expressed by transient exogenous transfection through IP. In order to investigate the early stage of ASFV infection, ASFV was inoculated into PAMs and samples of ASFV infection for 0, 15, 30, and 60 minutes were prepared, respectively. CD1d was used as the primary antibody and the expression of CD1d protein was detected by WB. Plasmids pCAGGS-HA-CD1d and pCAGGS-Flag-CD2v were co transfected into HEK293T cells. After 24 hours, the cells were collected for lysis, and Flag beads overnight binding protein was added. The interaction was detected by WB staining. At the same time, the plasmids were cotransfected into HEK293T cells in a confocal dish, incubated with labeled antibodies, and corresponding fluorescent secondary antibodies were selected. The co localization of CD1d and CD2v was observed under a laser confocal microscope. Verification of Co-IP interaction between CD1d and ASFV outer capsule protein CD2v was verified.【Result】 The GST-CD1d protein expressed in prokaryotic cells was expressed in the form of inclusion bodies, with a molecular weight of approximately 35 ku; After four rounds of immunization with CD1d recombinant protein in experimental rabbits, blood was collected and serum was separated. The purified antibody was detected by SDS-PAGE and showed a specific band at 45 and 25 ku, respectively, representing the heavy and light chains of the CD1d antibody. The rabbit anti CD1d antibody prepared using purified CD1d protein as immunogen contained both heavy and light chains, and had good purity; This antibody could identify the expression and cellular localization of transient transfected exogenous and PAMs endogenous CD1d proteins through WB and IFA. Further testing results showed that after ASFV infection with PAMs, the expression level of CD1d protein significantly increased, and WB and IFA results showed that CD1d interacted and co localized with the outer capsule protein CD2v encoded by ASFV. 【Conclusion】 This study prepared antibodies against CD1d through prokaryotic expression technology, laying the foundation for further exploration of the biological function of CD1d protein in ASFV infection process.
CD1d protein; prokaryotic expression; polyclonal antibody; African swine fever virus; CD2v protein

10.3864/j.issn.0578-1752.2024.08.015
2022-11-17;
2024-03-01
十四五國家重點研發計劃(2021YFD1800100)、國家自然科學基金(32172874)
劉傳霞,E-mail:18264110178@163.com。通信作者鄭君,E-mail:zhengjun01@caas.cn
(責任編輯 林鑒非)
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
