富硅或富磷育秧降低水稻砷含量的效果及其分子機制
2024-05-09 03:35:06賈夢凡李燕麗王興祥周志高丁昌峰
生態與農村環境學報 2024年4期
關鍵詞:水稻
賈夢凡,李燕麗,王興祥,周志高,丁昌峰②
〔1.濕地生態與農業利用教育部工程研究中心/ 長江大學農學院,湖北 荊州 434025;2.土壤與農業可持續發展國家重點實驗室(中國科學院南京土壤研究所),江蘇 南京 210008;3.中國科學院大學,北京 100049〕
砷(As)是我國農田主要的重(類)金屬污染物之一,2014年《全國土壤污染狀況調查公報》表明,我國2.7%的土壤樣點As含量超標。由于水稻自身的特性及淹水栽培模式,其富集As的能力遠遠大于其他旱作物[1],稻米是人體攝入As的主要食物來源[2]。硅(Si)是水稻的有益元素,磷(P)是水稻的必需元素,對水稻的高產穩產十分關鍵。由于硅酸和亞砷酸具有相似的解離常數和分子大小,三價砷〔As(Ⅲ)〕可以通過硅酸的轉運通道蛋白OsLsi1進入水稻根系,進而通過OsLsi2在根中進行橫向運輸進入木質部。因此,施Si可調控水稻對As的吸收以及轉運[3];由于五價砷〔As(Ⅴ)〕與P有非常相似的化學性質和結構,水稻根系可通過P吸收通道吸收As(Ⅴ),再通過P的轉運子(OsPT1、OsPT4、OsPT8)進入細胞[4]。
前人對外源添加Si或P調控As在水稻體內的積累進行了較多研究。例如,在自然土壤中(As含量≤15 mg·kg-1),添加10~20 g·kg-1硅酸形態的Si肥可降低水稻秸稈和糙米中As含量,降幅可達65%~78%和11%~23%[5-6]。但Si肥如果用量不當,反而會通過促進土壤固相對As(Ⅲ)、As(Ⅴ)以及二甲基砷(DMA)的釋放,增加土壤溶液中As濃度,提高土壤中As的生物有效性[7-8]。此外,在營養生長階段,葉面噴施Si可顯著降低水稻糙米中總As、無機砷及二甲基砷的積累[9]。但由于作物葉片對于養分的吸收效果受到自身營養狀況、生育時期、環境條件以及葉面肥種類等多種因素的影響,葉面噴施的效果不太穩定[10]。雷鳴等[11]通過盆栽試驗證明,向As污染土壤中施用磷酸氫二鈉或羥基磷灰石能使糙米總As含量顯著降低8%~22%。但由于P能夠通過離子交換作用置換出土壤中的As[12-13],在水稻生育期內施用大量的磷肥反而會增加土壤As的生物有效性[14]。
除此之外,也有研究證實內源Si或P對于苗期水稻降As有顯著作用。通過水培試驗研究發現,營養液中提前1~2周加入0.5~2 mmol·L-1的Si進行水稻幼苗培養,再進行As(Ⅴ)暴露,可以使地上部和根系As(Ⅴ)含量分別降低37%和22%,表明內源Si可顯著調控水稻對As(Ⅴ)的吸收轉運[15]。WANG等[16]研究表明,水稻幼苗體內P含量較低的情況下對As更敏感。因此,為避免出現土壤施Si或P提高土壤溶液中As有效性的情況以及葉面噴施效果不穩定的問題,在水稻育秧階段進行Si或P元素富集,增加水稻內源Si或P含量,可能是調控水稻吸收和累積As的一個有效措施。但目前關于秧苗富集Si或P對移栽后水稻糙米As含量的影響鮮見報道。
因此,該研究擬通過育秧過程中添加不同濃度硅酸和磷酸鹽以獲取富Si或P秧苗,以期篩選出能夠有效降低糙米As含量的最優濃度,研究富Si或P秧苗對 Si和 P 的積累特征及移栽后水稻植株對 As 的吸收轉運特征,明確富Si或P秧苗對于糙米的降As效果及機制,為As超標農田安全利用提供技術支撐。
1 材料與方法
1.1 供試水稻品種
供試水稻品種為“天優華占”秈型三系雜交稻,全生育期123 d左右。
1.2 植物培養
秧苗培育在中國科學院南京土壤研究所溫室進行,水稻種子用φ=30.0%的H2O2消毒15 min,去掉癟粒,用去離子水洗凈后置于25 ℃黑暗環境中浸種24 h,再轉移至濕潤紗布中催芽48 h,挑選出芽狀況良好且一致的種子置于浮板上進行育苗。育苗期間保證每個處理水稻種子數大致相同。育苗初期采用1/2水稻營養液培育3 d,再用正常濃度營養液進行培養。營養液培養實驗為每3 d進行1次營養液的更換育苗過程,水稻營養液采用木村B營養液(pH值5.6),具體成分包括0.36 mmol·L-1(NH4)2SO4、0.55 mmol·L-1MgSO4·7H2O、0.18 mmol·L-1KNO3、0.37 mmol·L-1Ca(NO3)2·4H2O、0.18 mmol·L-1KH2PO4、20 μmol·L-1FeSO4·7H2O、20 μmol·L-1EDTA-Na2、0.50 μmol·L-1MnCl2·4H2O、3.0 μmol·L-1H3BO3、1.0 μmol·L-1(NH4)6Mo7O24·4H2O、0.40 μmol·L-1ZnSO4·7H2O、0.20 μmol·L-1CuSO4·4H2O。
盆栽試驗供試土壤為采自江西贛州的As污染水稻土,土壤pH值為5.19,有機質含量為34 g·kg-1,總As含量為49.1 mg·kg-1。
1.3 試驗設計
秧苗培育在中國科學院南京土壤研究所溫室進行,水稻種子消毒、浸種、催芽后進行育苗。育苗初期采用1/2水稻營養液培育3 d,再進行Si、P處理,以硅酸(由K2SiO3溶液過氫型陽離子交換樹脂Amberlite IR 120制備而得)以及磷酸二氫鉀的形式加入。共包括5個處理,分別如下:(1)全水稻營養液(CK);(2)全水稻營養液+ 5 mmol·L-1Si (Si1);(3)全水稻營養液+ 8 mmol·L-1Si(Si2);(4)全水稻營養液+ 2 mmol·L-1P(P1);(5)全水稻營養液+ 4 mmol·L-1P (P2)。
每3 d更換1次營養液,培育25 d后獲得富Si或富P水稻幼苗,采集部分水稻幼苗分析地上部及地下部生物量和Si、P積累量,篩選出較優富集濃度下的富集秧苗用于土壤盆栽試驗。水稻幼苗移栽前一周,進行盆栽試驗裝土和添加基肥等前處理。每盆裝5 kg風干土,裝土過程中拌入基肥:0.20 g·kg-1N(尿素)、0.15 g·kg-1P2O5〔Ca(H2PO4)2·H2O〕、0.20 g·kg-1K2O(KCl),加水保持土壤濕潤,平衡1周左右。所有處理均設置3次重復。水稻幼苗培育25 d后移栽到盆栽土壤中,每盆種植2株,移栽后保持土壤淹水,水稻種植至成熟期收獲。
1.4 樣品采集
采集水培25 d后各處理水稻幼苗,幼苗根系在0.50 mmol CaCl2溶液中浸泡20 min,去除根表吸附的元素,清洗干凈后分成根系和莖葉,75 ℃烘干并測定各部位干物質質量以及Si、P含量。另取部分富集秧苗,擦干根系表面營養液后迅速做好標記并放入液氮中,轉移至-80 ℃冰箱中保存,待提取 RNA,測定砷相關轉運基因(OsLsi1、OsLsi2、OsPT1、OsPT4、OsPT8、OsABCC1)的相對表達量。水稻成熟后采集根系、莖葉、稻穗,根系及莖葉經去離子水清洗干凈后75 ℃烘干并測定總As含量,稻谷經礱谷機去殼后獲得糙米,冷凍干燥后消解測定各形態As含量。
1.5 樣品處理與測定
1.5.1樣品中元素含量的測定
樣品中Si含量采用w=50%的NaOH高壓滅菌法消解并用分光光度計測定[17];樣品中P含量采用濕式消解鉬銻抗比色法測定[18];總As含量采用硝酸-過氧化氫消解[19],電感耦合等離子質譜法(ICP-MS)測定。
1.5.2樣品中As形態的提取與測定
采用φ=15%的硝酸熱浸提法提取,液相色譜-電感耦合等離子質譜法(LC-ICP/MS)測定,色譜柱為反相柱[19]。
1.5.3As相關轉運基因相對表達量的測定
在液氮中將待測水稻根系研磨成粉末狀,采用UNIQ-10柱式Trizol總RNA提取試劑盒(上海生工生物工程股份有限公司)提取總RNA。采用核酸儀(美國賽默飛NanoDrop ND-1000)測定各樣品總RNA濃度,采用反轉錄試劑盒〔南京諾唯贊HIScript Ⅲ RT SuperMix for qPCR(+gDNA wiper)〕和PCR儀(日本Takara)對RNA進行反轉錄過程合成cDNA。OsLsi1、OsLsi2、OsPT1、OsPT4、OsPT8、OsABCC1等基因的熒光定量過程采用熒光定量試劑盒(南京諾唯贊Taq Pro Universal SYBR qPCR Master Mix)、RT-PCR儀(美國伯樂CFX96)完成。
1.6 數據處理
以根-莖為例,轉運系數(TF,FT)的計算方法為莖As含量與根As含量的比值,表示As由水稻根系向莖轉運的能力。
采用Excel 2021和SPSS 19.0軟件對試驗數據進行單因素方差分析(ANOVA),采用Duncan多重比較進行顯著性檢驗,GraphPad Prism 8軟件作圖,采用Origin 2021軟件作相關性熱圖。
2 結果與分析
2.1 富Si或富P育秧對水稻幼苗吸收Si或P的影響
不同濃度處理下富Si秧苗中Si含量如表1所示。Si1和Si2處理下秧苗的生長沒有受到抑制,地上部與根系Si含量與對照相比均顯著增加,且Si1處理根系Si含量顯著高于Si2。Si1和Si2處理下地上部Si含量分別增至對照的1.9 和1.8倍,根系Si含量分別增至對照的2.3 和 2.0倍,秧苗整株Si吸收量與對照相比也呈顯著增加趨勢,分別增至對照的19.6和18.3倍。
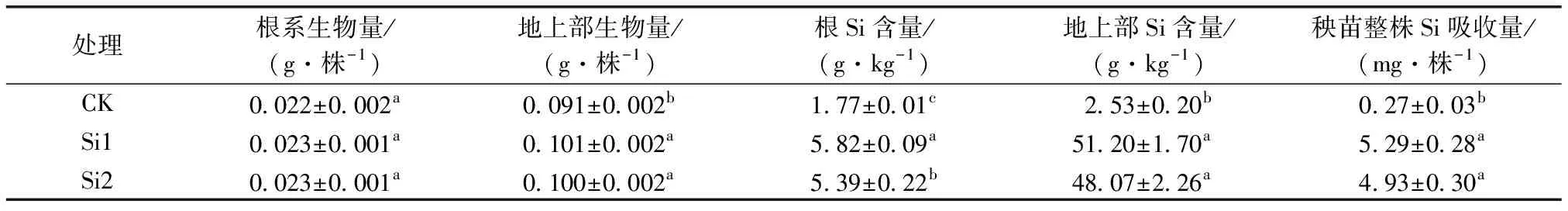
表1 不同濃度處理富Si秧苗生物量及Si含量
不同濃度處理下富P秧苗中P含量如表2所示。P1處理下秧苗生長沒有受到抑制,但P2處理地上部生物量較對照顯著降低7.7%;地上部與根系P含量與對照相比均顯著增加,P1和P2處理根系P含量分別增至對照的2.1和2.6倍,地上部P含量分別增至對照的2.2和 2.3倍,秧苗整株P吸收量與對照相比也呈顯著增加趨勢,分別增至對照的2.3和2.1倍。
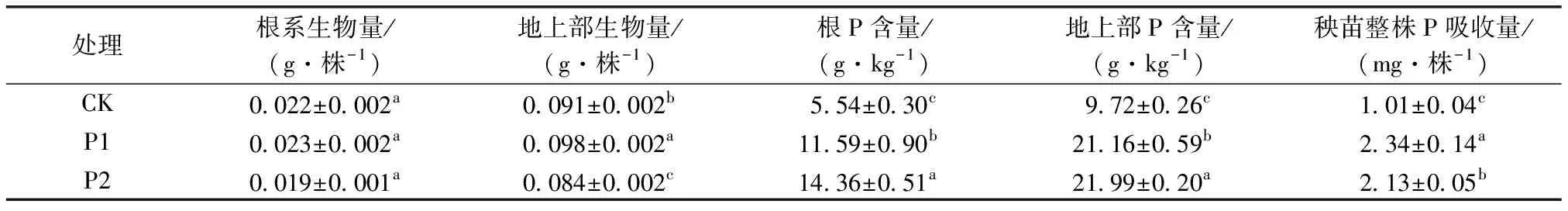
表2 不同濃度處理富P秧苗生物量及P含量
由于Si1處理下秧苗整株Si吸收量略高于Si2處理,而與P1相比,P2處理秧苗地上部的生物量顯著降低,影響了水稻正常生長。據此,筆者選擇Si1和P1這2個濃度處理的富集秧苗,并將其移栽至As污染土壤中進行盆栽試驗。
2.2 富Si或富P育秧對水稻產量及各部位As含量的影響
表3為富Si或富P秧苗移栽至As污染土壤后的產量以及各部位總As含量。富Si或富P處理對于水稻產量沒有顯著影響。從表3可知,與對照相比,Si1處理糙米和葉總As含量分別降低31.1%和16.8%,水稻根總As含量顯著增加22.7%,莖總As含量沒有顯著變化;與對照相比,P1處理水稻根系和莖總As含量分別顯著增加13.4%和16.0%,糙米、水稻葉的總As含量均沒有顯著變化。

表3 富Si或富P秧苗移栽后的水稻產量及各部位總As含量
2.3 富Si或富P育秧對水稻體內As轉運的影響
富Si或富P育秧可不同程度影響水稻各部位之間As的轉運系數。由圖1可知, Si1處理使得水稻各部位間As轉運系數均顯著降低;P1處理水稻根-莖、根-葉轉運系數與對照相比無顯著差異,但莖-葉、莖-糙米的轉運系數顯著降低。
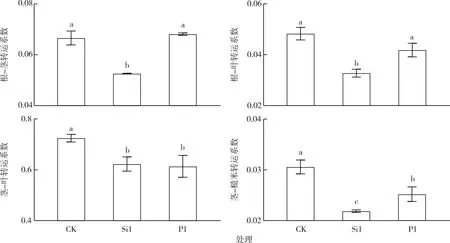
CK—全水稻營養液;Si1—全水稻營養液+5 mmol·L-1 Si;P1—全水稻營養液+2 mmol·L-1 P。同一幅圖中直方柱上方小寫字母不同表示不同處理間某指標差異顯著(P<0.05)。
2.4 富Si或富P育秧對糙米As形態的影響
糙米As形態測定的試驗過程中并未檢測出一甲基砷的存在。圖2為最優Si或P濃度水稻秧苗培育至成熟期后所獲糙米的As(Ⅲ)、As(Ⅴ)以及DMA含量。從圖2可以看出,與常規育秧相比,富Si育秧使得糙米中3種形態As含量均顯著降低,下降率分別達32.1%、58.3%、33.5%,其中無機砷含量〔As(Ⅲ)與As(Ⅴ)總含量〕顯著降低37.3%;富P育秧使得As(Ⅴ)含量降低59.2%,但對于糙米中As(Ⅲ)和DMA含量沒有顯著影響,其中無機砷含量有降低趨勢,但與對照相比無顯著差異。
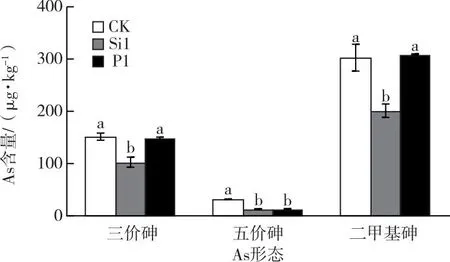
CK—全水稻營養液;Si1—全水稻營養液+5 mmol·L-1 Si;P1—全水稻營養液+2 mmol·L-1 P。同一組直方柱上方小寫字母不同表示不同處理間某指標差異顯著(P<0.05)。
2.5 富Si或富P育秧對As轉運基因相對表達量的調控
富Si育秧會顯著影響As相關轉運基因的相對表達量。圖3表明,與對照相比,富Si育秧對OsLsi1的相對表達量沒有顯著影響,使OsLsi2的相對表達量下降26%,OsABCC1的相對表達量上調203%。
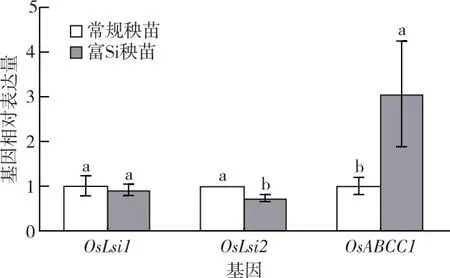
同一組直方柱上方小寫字母不同表示不同處理間某指標差異顯著(P<0.05)。
富P育秧也會顯著影響As相關轉運基因的相對表達量。由圖4可見,與常規育秧相比,富P育秧使OsPT1、OsPT4及OsPT8的相對表達量分別下降26%、51%和71%,OsABCC1的相對表達量上調22%。
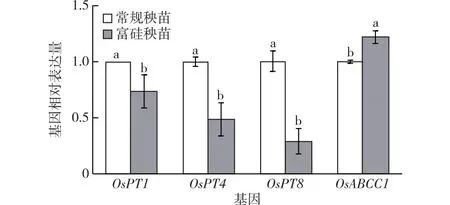
同一組直方柱上方小寫字母不同表示不同處理間某指標差異顯著(P<0.05)。
2.6 水稻不同組織As含量與秧苗Si或P吸收量及轉運基因表達量的相關性分析
相關性分析結果(圖5)表明,水稻糙米無機砷含量與秧苗Si吸收量、成熟期水稻根As含量、OsABCC1的相對表達量呈顯著負相關關系,與OsLsi1、OsLsi2的相對表達量以及成熟期水稻葉總As含量呈顯著正相關關系,與其他因素相關性不顯著。根系總As含量與秧苗Si吸收量、OsABCC1的相對表達量呈顯著正相關關系,與OsLsi2的相對表達量呈顯著負相關關系。OsLsi1和OsLsi2的相對表達量與秧苗Si吸收量呈顯著負相關關系,OsABCC1的相對表達量與秧苗Si吸收量呈顯著正相關關系,這表明富Si抑制了OsLsi1和OsLsi2基因的表達,且誘導了OsABCC1的過量表達。OsLsi1與根系吸收Si、As有關,OsLsi2與木質部轉運Si、As有關,OsABCC1與根系液泡固定重金屬螯合物有關,表明富Si秧苗可能通過調控OsLsi1、OsLsi2以及OsABCC1的表達,從而抑制水稻植株As含量,進一步影響糙米As含量。
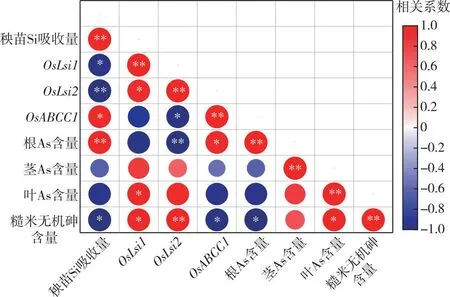
*表示P<0.05,**表示P<0.01。
圖6顯示,糙米無機砷含量與OsABCC1的相對表達量呈顯著負相關關系,與秧苗P吸收量呈負相關關系,與As相關轉運基因的相對表達量呈正相關關系。水稻成熟期根、莖總As含量與秧苗P吸收量呈顯著正相關關系,與OsPT1、OsPT4和OsPT8的相對表達量呈負相關關系,與OsABCC1的相對表達量呈顯著正相關關系,這說明秧苗富P在一定程度上可以影響水稻體內As的轉運。OsPT1、OsPT4和OsPT8的相對表達量與秧苗P吸收量呈負相關關系,OsABCC1的相對表達量與秧苗P吸收量呈顯著正相關關系,這說明秧苗富P抑制了OsPT1和OsPT4的表達,誘導了OsABCC1的過量表達。OsPT1和OsPT4與根系吸收P、As有關,OsABCC1與根系液泡固定重金屬螯合物有關,表明富P秧苗可能通過調控OsPT1、OsPT4以及OsABCC1的表達,從而抑制水稻植株對As的吸收并增強根系對于As的滯留,進一步影響糙米As含量。
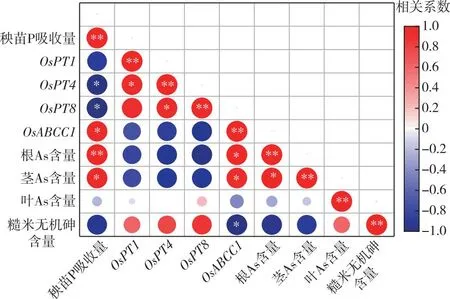
*表示P<0.05,**表示P<0.01。
3 討論
筆者試驗結果表明,適宜Si或P濃度下培育出的秧苗對于成熟期糙米中As含量有降低作用,富Si秧苗顯著降低了糙米As(Ⅲ)含量,富P秧苗顯著降低了糙米As(Ⅴ)含量,且富Si或P秧苗根系中As相關轉運基因的相對表達量均呈現顯著下降趨勢。一般來說,在稻田土壤溶液中As主要以As(Ⅲ)和As(Ⅴ)的形態存在[20],而在水稻根系中,單硅酸〔Si(OH)4〕和亞砷酸〔As(OH)3〕共用同一吸收通道蛋白OsLsi1,水稻在吸收單硅酸時會吸收亞砷酸進入體內[21-22],單硅酸進入水稻體內后由OsLsi2向木質部運輸,進一步向地上部轉運。As(Ⅴ)由于化學性質與磷酸鹽相似,主要通過磷酸鹽轉運蛋白OsPT4、OsPT8進入水稻根系細胞[23-24]。由于水稻為典型的Si高積累植物,P是植物生長所必需的大量元素,而As為非必需元素,轉運蛋白可能會優先轉運必需的營養元素以滿足水稻生長需求。當水稻組織中Si或P元素充足時,相關轉運基因的表達會受到抑制,從而影響水稻根系對As的轉運吸收。CHEN等[25]也發現,施用鐵肥抑制了根系OsIRT1、OsNramp1和OsNramp5等基因的表達,進而降低了水稻對As的吸收。
試驗發現,5 mmol·L-1Si富集處理顯著降低糙米As(Ⅴ)含量,這是由于As(Ⅴ)通過磷酸鹽通道進入水稻根系后,一部分通過磷酸鹽轉運蛋白進入木質部和韌皮部向上運輸,另一部分會被還原成As(Ⅲ)[26]。被還原的As(Ⅲ)部分被隔離在液泡中,部分通過轉運蛋白OsLsi2等由木質部和韌皮部向上運輸[27],富Si處理通過影響這一部分As(Ⅲ)的轉運從而影響糙米中As(Ⅴ)含量。有研究證實,水稻水通道蛋白OsLsi1會介導甲基砷的吸收[28],這可能是Si1處理能夠顯著降低糙米中DMA含量的原因。
相較于對照,Si或P富集秧苗顯著增加了成熟期水稻根系中總As含量,一方面是富Si或富P處理增加了水稻根系中轉運基因OsABCC1的相對表達量,而OsABCC1負責將As(Ⅲ)轉運并固定在液泡中,抑制了As(Ⅲ)在水稻體內的向上轉運[29],增強了As在根系中的滯留;另一方面,富Si或富P處理降低了As相關轉運基因OsLsi2、OsPT4、OsPT8的相對表達量,阻礙了As的向上轉運,也在一定程度上增強了As在根系中的滯留[23-24,27]。
試驗結果顯示,相較于對照,5 mmol·L-1Si富集處理顯著降低了糙米中As(Ⅲ)、As(Ⅴ)、DMA含量及水稻糙米總As含量,而2 mmol·L-1P富集處理僅顯著降低As(Ⅴ)含量,對糙米總As含量沒有顯著影響。這可能是因為稻田淹水條件會促進土壤微生物對As的還原和甲基化[30],使得淹水條件下土壤環境中As形態以As(Ⅲ)和DMA為主,導致糙米中的As主要以DMA和As(Ⅲ)為主。WANG等[31]對于湖南As污染水稻土(總As含量為38.2 mg·kg-1)的盆栽試驗同樣發現,稻米中有機砷占比(77%)顯著高于無機砷(23%)。
綜上所述,適宜濃度的Si肥和P肥培育后的秧苗均可降低糙米中As含量,且與2 mmol·L-1富P育秧相比,5 mmol·L-1富Si育秧降As效果更好。富Si或富P育秧是一項低成本、輕簡化的As污染稻田安全利用技術,能夠突破大田全生育期繁瑣的控As技術弊端,不會對大田土壤產生次生障礙風險,且此方法降低了過度施用Si或P肥造成農業面源污染的隱患。
需要指出的是,筆者雖然研究了富Si或富P秧苗對水稻植株的降 As 效果,但并未探討苗期富集水稻對于水稻整個生育期的持續影響,且僅為初步研究結果,需進一步深入研究其生物學機制。此外,該試驗僅探討了單一水稻品種富Si或富P育秧的降As效果,對于不同品種的普適性還需要進一步研究。
4 結論
(1)研究篩選出的適宜富Si或富P育秧濃度均顯著增加了秧苗體內Si或P含量,且對水稻生長沒有顯著影響。
(2)5 mmol·L-1Si育秧處理能夠調控As相關轉運基因OsLsi1、OsLsi2以及OsABCC1的表達,增強As在水稻根系中的滯留,顯著降低糙米中As(Ⅲ)、As(Ⅴ)、DMA含量。
(3)2 mmol·L-1P育秧處理能夠調控As相關轉運基因OsPT1、OsPT4、OsPT8以及OsABCC1的表達,增強了As在水稻根系中的滯留,顯著降低糙米中As(Ⅴ)含量。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
