梅山豬和杜洛克豬卵泡中差異表達lncRNA克隆鑒定及與miRNAs相關性分析
2024-05-14 03:23:08張化鵬張慶澤何凡祁夢凡符彬彬李清春李夢尋馬力鵬劉乙黃濤
中國農業科學 2024年9期
關鍵詞:梅山
張化鵬,張慶澤,何凡,祁夢凡,符彬彬,李清春,李夢尋,馬力鵬,劉乙,黃濤,2
梅山豬和杜洛克豬卵泡中差異表達lncRNA克隆鑒定及與miRNAs相關性分析
張化鵬1,張慶澤1,何凡1,祁夢凡1,符彬彬1,李清春1,李夢尋1,馬力鵬1,劉乙1,黃濤1,2
1石河子大學動物科技學院,新疆石河子 832000;2新疆生豬種業工程技術研究中心,新疆昌吉 831100
【目的】通過克隆鑒定梅山豬和杜洛克豬卵泡期第4天M2卵泡中差異表達的lncRNA-ALDBSSCT0000005583,分析其在豬顆粒細胞與miRNAs表達量的相關性,為探究lncRNA調控miRNA在母豬卵泡發育過程中的作用提供理論依據。【方法】對課題組前期在梅山和杜洛克M2卵泡中篩選出的差異表達lncRNA-ALDBSSCT0000005583,進行RT-qPCR驗證,并利用RACE克隆其全長序列;根據編碼潛力評估工具CPAT、CPC對該lncRNA的編碼能力進行預測,原核表達試驗進一步鑒定其編碼能力;核質分離試驗對lncRNA-ALDBSSCT0000005583進行亞細胞定位,RT-qPCR檢測其在各組織中的表達水平;利用miRBase網站查找豬的miRNA數據庫,結合RNAhybrid、miRanda在線軟件預測與lncRNA-ALDBSSCT0000005583有相互作用的物種間保守miRNA,使用TargetScan和miRanda預測與lncRNA-ALDBSSCT0000005583具有相互作用miRNA的靶基因,并對其靶基因進行GO富集和KEGG信號通路分析;通過過表達以及干擾lncRNA驗證其對miRNA表達量的影響。【結果】lncRNA- ALDBSSCT0000005583在杜洛克豬M2卵泡中的表達量顯著高于梅山豬,lncRNA 5'RACE和3'RACE片段大小為569 bp和546 bp,測序分析表明lncRNA-ALDBSSCT0000005583大小為588 bp。生物信息學預測其編碼潛能較低,同時原核表達試驗結果進一步證明其不編碼蛋白質。組織表達譜分析表明,lncRNA-ALDBSSCT0000005583在腎上腺、脾肝和卵巢中表達量較高,在下丘腦和心臟中的表達量較低,亞細胞定位結果顯示該lncRNA主要存在于細胞質中。生物信息學分析后共篩選出9個物種間保守的miRNA與lncRNA-ALDBSSCT0000005583具有潛在的相互作用,其中有兩個與卵巢發育相關的miRNA:miR-193a-5p、miR-361-3p;KEGG和GO富集分析顯示miR-193a-5p、miR-361-3p的靶基因與系統進程和細胞與細胞信號傳導等生物學過程有關,并顯著參與到催產素、Ras、NF-κB、促性腺激素釋放激素分泌等通路。隨后在顆粒細胞中過表達lncRNA-ALDBSSCT0000005583,經RT-qPCR發現miR-193a-5p、miR-361-3p的表達量均顯著下調(<0.05),但干擾lncRNA-ALDBSSCT0000005583后無顯著影響。【結論】lncRNA-ALDBSSCT0000005583是一個不具備編碼蛋白質能力的lncRNA,在梅山與杜洛克豬中等卵泡間表達量有極顯著差異,在腎上腺、脾肝和卵巢中表達量較高,主要存在于顆粒細胞的細胞質中,可能與miR-193a-5p、miR-361-3p相互作用,從而參與豬卵巢顆粒細胞的發育過程。
lncRNA-ALDBSSCT0000005583;豬;卵泡;顆粒細胞;miR-193a-5p;miR-361-3p
0 引言
【研究意義】長鏈非編碼RNA(long non-coding RNA lncRNA)是長度超過200個堿基的轉錄本,沒有蛋白質編碼潛能[1]。越來越多的研究表明,lncRNA參與各種重要生物過程的調控,包括染色質修飾、選擇性剪接、表觀遺傳修飾、充當分子骨架、轉錄和轉錄后調控[2- 6]。lncRNA既可以充當競爭性內源性RNA(ceRNA),也可以“吸附”miRNA或蛋白質,調節mRNA的翻譯[7]。母豬的繁殖力是養豬業的一個重要特征,梅山豬以其高繁殖力和較大的窩產仔豬數而聞名[8]。與杜洛克母豬相比,梅山母豬在卵泡期中后期可以維持較多的中型卵泡庫,產生更多的排卵前卵泡,有可能排出更多的成熟卵子并生下更多的胎兒[9-10]。因此研究梅山豬和杜洛克豬M2卵泡中差異表達的lncRNA的具體功能及調控機制,對提高母豬的繁殖力具有重要意義。【前人研究進展】近年來隨著高通量測序技術的發展,許多研究表明lncRNA可以調節卵泡發育,進而影響母豬的繁殖力。HU等[11]鑒定了大白母豬在發情周期的卵泡期和黃體期的繁殖力相關的卵巢lncRNA,并發現卵巢中的lncRNA顯著影響豬的生育能力。LI等[10]通過轉錄組測序在梅山豬和杜洛克豬M2卵泡中共鑒定出3 554個lncRNA,其中與繁殖性狀有關的差異表達lncRNA有127個。LIU等[12]在杜洛克豬卵泡期的第0、2和4天鑒定了豬卵巢的lncRNA和mRNA表達譜,發現lncRNA- ENSSSCT00000034907可能在卵泡發育中發揮重要作用。【本研究切入點】前人研究表明,lncRNA在豬的卵巢中發揮著重要作用,但是關于豬卵巢lncRNA的研究只集中于篩選出卵巢中差異表達的lncRNA,并未進行下一步驗證。【擬解決的關鍵問題】本研究利用本課題組前期出在梅山和杜洛克豬M2卵泡篩選的差異表達lncRNA-ALDBSSCT0000005583進行RACE克隆,并利用生物信息學對其靶向miRNA進行預測,結合組織表達譜分析其表達量,并驗證其在顆粒細胞中的亞細胞定位,從而為豬lncRNA通過miRNA發揮功能的調控機制研究提供試驗依據。
1 材料與方法
試驗于2022—2023年在石河子大學動物科技學院遺傳育種與繁殖實驗室完成。
1.1 試驗材料
1.1.1 試驗樣品來源 2023年8月,在石河子大學附屬獸醫站,選取體重接近、健康大白豬三頭,屠宰后采集心、胃、大腦、脾、腎上腺、肝、小腸、卵巢、子宮等組織,DEPC處理后,錫紙包裹于液氮中凍存,帶回實驗室備用;梅山和杜洛克豬M2卵泡樣品為石河子大學動物科技學院遺傳育種與繁殖實驗室保存。細胞在石河子市綠源達康屠宰場中采集的豬卵巢,經處理后凍存于液氮中的原代顆粒細胞。
1.1.2 主要試劑與儀器 主要試劑:pCDNA3.1(+)載體;pET-28a(+)載體;DH5α感受態細胞為實驗室保留;siRNA oligos由上海吉瑪公司合成;lipofectamine? 3000(賽默飛);Trizol up(全式金);SMARTer?RACE cDNA Amplification Kit(TaKaRa);DAPI染色劑(碧云天);一抗,卵泡刺激素受體(FSHR),二抗,CY3-羊抗兔IgG;NE-PERTM細胞核和細胞質提取試劑盒(賽默飛)等。
主要儀器:超低溫冰箱(-80℃)、CO2培養箱、高壓滅菌鍋、LightCycle 96實時熒光定量PCR儀、高速離心機、Nano-Drop ND 2000c Spectrophotometer、熒光倒置顯微鏡等。
1.2 總RNA提取及cDNA合成
根據TRIzolip up試劑盒說明書提取樣本總RNA,用1%的瓊脂糖凝膠電泳和NanoDrop 2000進行總RNA的完整性和純度檢驗合格后備用。
按照TaKaRa反轉錄試劑盒PrimeScriptTM RT ReagentKit with gDNA Eraser(PerfectReal Time)說明書進行cDNA合成。
1.3 熒光定量PCR和lncRNA-ALDBSSCT0000005583克隆及分析
根據實驗室之前二代測序結果中lncRNA- ALDBSSCT0000005583的部分已知核苷酸序列,使用Primer Primer 5.0軟件設計實時熒光定量PCR引物、5′RACE和3′RACE特異性引物。實時熒光定量PCR內參基因選擇GAPDH,在用于RACE克隆的特異性引物(GSP)在5′端添加15 bp“GATTACGCCAA GCTT”的序列用于后續實驗中的連接測序,所有引物見表1。所有引物均由生工生物工程(上海)股份有限公司合成。
1.4 lncRNA-ALDBSSCT0000005583編碼能力預測
通過編碼潛力評估工具CPAT(coding-potential assessment tool)、CPC(http://lilab.research.bcm.edu/)在線預測工具對lncRNA-ALDBSSCT0000005583進行編碼潛能預測。
1.5 過表達載體以及原核表達載體構建
以杜洛克豬M2卵泡cDNA為模板,擴增lncRNA全長,經2 %的瓊脂糖凝膠電泳(120 V,40 min)檢測后,切取目的條帶,膠回收,經雙酶切后與pcDNA3.1(+)載體進行連接,轉化,測序。
將lncRNA-ALDBSSCT0000005583全長和YY1的CDs區亞克隆到Pet-28a(+)載體的RⅠ和Ⅰ酶切位點上,獲得lncRNA-ALDBSSCT0000005583和YY1原核表達質粒,轉化至BL21(DE3)感受態細胞,獲得重組質粒后進行酶切和測序鑒定。YY1為實驗室保存的pcDNA3.1過表達載體,經RⅠ和Ⅰ雙酶切后連接至Pet-28a(+)載體上,作為陽性對照,空載體作為陰性對照。
經測序鑒定成功的陽性菌液,37 ℃搖床200 r/min培養至OD 0.6—0.8,以Pet-YY1為陽性對照,空載體為陰性對照,各加入終濃度為1.0 mmol·L-1IPTG溶液,誘導表達4 h。取1 mL誘導后的菌液,12 000 r/min離心5 min,棄去上清,根據菌液沉淀的量,用50—100 μL PBS重懸沉淀,加入5×loading buffer上樣緩沖液,金屬浴中100 ℃10 min,保證菌體完全裂解,12 000 r/min離心5 min后,取10 μL用于SDS-PAGE電泳分析。SDS凝膠用考馬斯亮藍染色2 h,脫色液脫色后,在凝膠成像系統上觀察結果。
1.6 豬顆粒細胞培養與鑒定
常規的細胞復蘇,向復蘇的細胞中加入10 mL完全培養基(DMEM,FBS,雙抗)輕柔吹勻,根據細胞密度合理鋪入6孔板中。將鋪好的細胞于37 ℃、5% CO2培養箱中,培養細胞,48 h后使用PBS清洗顆粒細胞。運用免疫熒光的方法檢測細胞中卵泡刺激素受體(FSHR),以鑒定顆粒細胞。
1.7 lncRNA亞細胞定位
試驗采用豬顆粒細胞,按照NE-PER? 細胞核和細胞質提取試劑盒說明書進行細胞核細胞質RNA分離,通過qPCR檢測lncRNA-ALDBSSCT0000005583在豬顆粒細胞細胞核細胞質中的表達量。
1.8 顆粒細胞轉染及收集
6孔板培養顆粒細胞待融合度達到70%—90%時,使用Lipofectamine? 3000試劑盒將過表達載體pCDNA3.1-lncRNA-ALDBSSCT0000005583以及siRNA- ALDBSSCT0000005583轉染至豬顆粒細胞,以空質粒pCDNA3.1及siRNA-NC作為對照。利用RT-qPCR檢測轉染處理48 h后的過表達/抑制的效率。
1.9 lncRNA-ALDBSSCT0000005583-miRNA靶基因預測
從miRBase(https://www.mirbase.org)獲得了豬的成熟miRNA序列,利用RNAhybrid(https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid)和miRanda(https://www.miranda-ng.org/en/)分別預測與lncRNA- ALDBSSCT0000005583存在相互作用的miRNA。對兩款軟件預測出的miRNA分別按照每款軟件的評分標準進行篩選,取兩款軟件的交集作為最終的miRNA。使用TargetScan(v5.0)、miRanda(v3.3a)對篩選出的miRNA分別進行靶基因預測。利用KEGG和GO數據庫進行富集分析,用Cytoscape 3.6.0軟件構建候選miRNA-mRNA相互作用的結果圖。

表1 RT-qPCR及載體構建引物序列
F:上游引物;R:下游引物。表中加粗部分為保護堿基;下劃線部分為酶切位點;酶切位點為R Ⅰ和Ⅰ;波浪線部分為添加的序列,用于后續試驗中的無縫克隆。lncRNA-5583為lncRNA-ALDBSSCT0000005583縮寫,下同
F: Upstream primers; R: Downstream primers. The bold part in the table is the protective base; The underlined part is the enzyme cleavage site; The digestion sites wereR I. andI. The wavy portion is the added sequence for seamless cloning in subsequent experiments. lncRNA-5583 is an abbreviation for lncRNA-ALDBSSCT0000005583, The same as below
1.10 數據分析
實時熒光定量PCR以GAPDH作為內參,RT-qPCR結果按使用2-ΔΔct法計算,用SPSS17.0軟件進行顯著性分析。以GAPDH基因作為內參基因對檢測的目的基因進行歸一化,U6作為細胞核基因內參。數據使用 Graph Pad Prism 8 進行作圖,其中“*”(<0.05)為差異顯著,“**”(<0.01)為差異極顯著,“ns”(>0.05)表示差異不顯著。所有引物見表1。
2 結果
2.1 lncRNA-ALDBSSCT0000005583在M2卵泡中的表達分析
通過qRT-PCR技術對lncRNA-ALDBSSCT0000005583在梅山豬和杜洛克豬的M2卵泡的表達情況進行驗證分析發現,lncRNA-ALDBSSCT0000005583在杜洛克豬M2卵泡中的表達量極顯著高于梅山豬,結果與二代測序的結果相同(<0.01)(圖1),表明了二代測序結果的可靠性。
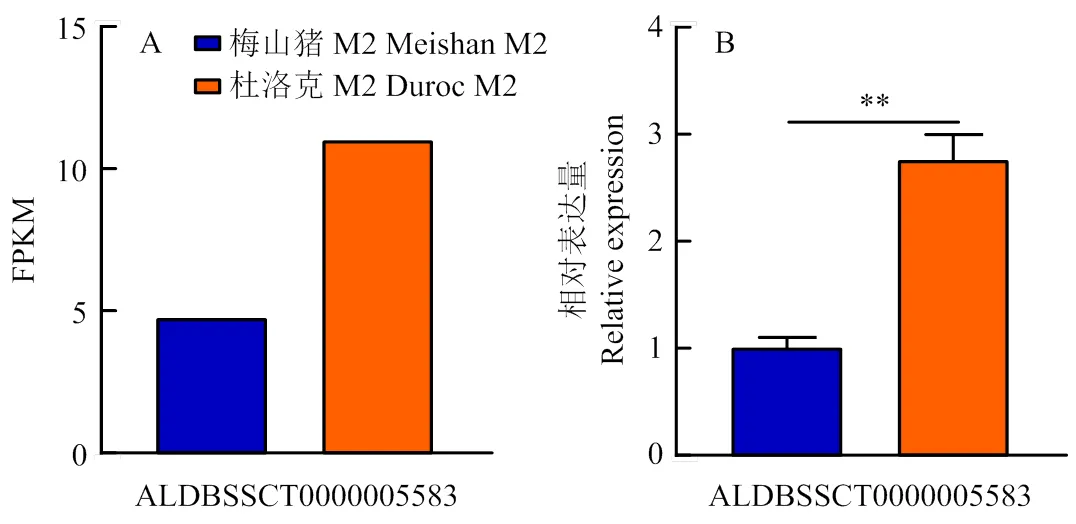
A:二代測序結果ALDBSSCT0000005583在M2卵泡中的表達水平;B:RT-qPCR驗證ALDBSSCT0000005583的表達水平
2.2 lncRNA-ALDBSSCT0000005583序列特征
根據SMARTer RACE 5'3' Kit 擴增豬長鏈非編碼lncRNA-ALDBSSCT0000005583,結果表明,3′RACE片段大小為546 bp(圖2-A),5′RACE片段大小為569 bp(圖2-B),根據3′RACE和5′ RACE測序序列結果拼接確定lncRNA-ALDBSSCT0000005583全長為588 bp(圖2-C)。
2.3 lncRNA-ALDBSSCT0000005583的鑒定
利用在線軟件CPAT和CPC對長鏈非編碼RNA lncRNA-ALDBSSCT0000005583的編碼能力進行分析計算,結果lncRNA-ALDBSSCT0000005583的編碼能力分別為0.0122981(表2)、0.01716(表3),屬于非編碼RNA。
為了進一步驗證lncRNA-ALDBSSCT0000005583是否具有編碼能力,成功構建了pET-28a-lncRNA- ALDBSSCT0000005583(圖3-A)、pET-28a-YY1(圖3-B)原核表達載體,通過原核表達試驗發現lncRNA- ALDBSSCT0000005583與pET-28a(+)空質粒的電泳條帶一致,而相同試驗條件下陽性對照pET- 28a-YY1則在69 kDa處有明顯的新增條帶(圖3-C)。綜合以上研究結果強調lncRNA-ALDBSSCT0000005583是一種真正的非編碼RNA。

表2 CPAT網站預測編碼潛能

表3 CPC網站預測編碼潛能的結果
2.4 顆粒細胞鑒定及lncRNA亞細胞定位
顆粒細胞的鑒定結果如圖4所示,鑒于FSHR是卵巢中唯一表達FSHR的細胞,通過檢測細胞中FSHR的表達情況判斷是否是顆粒細胞,免疫熒光結果顯示,培養的細胞中FSHR陽性率較高,判斷培養的細胞為顆粒細胞,可以用于進行下一步試驗。
利用豬的顆粒細胞進行核質分離試驗,確定了lncRNA-ALDBSSCT0000005583的亞細胞定位,結果顯示,lncRNA-ALDBSSCT0000005583主要存在于細胞質中(圖5)。
2.5 lncRNA-ALDBSSCT0000005583組織表達譜
試驗利用RT-qPCR技術,探究lncRNA- ALDBSSCT0000005583在豬不同組織中的表達情況。組織表達譜結果顯示,lncRNA-ALDBSSCT0000005583在心、下丘腦、胃、卵巢、脾、腎上腺、肝、小腸、子宮中均有不同程度表達,在腎上腺、脾、肝和卵巢中表達量較高,在下丘腦和心中的表達量較低(圖6),以上研究表明lncRNA-ALDBSSCT0000005583在這些組織中可能均具備生物學功能。
2.6 lncRNA-ALDBSSCT0000005583生物信息學分析
根據miRbase 22數據庫,下載豬的所有miRNAs,利用RNAhybrid和miRanda在線軟件分析與lncRNA-ALDBSSCT0000005583具有相互作用的miRNA,共發現9個在人、小鼠、大鼠、牛、豬等物種之間保守的miRNAs(圖7),利用TargetScan(v5.0)、miRanda(v3.3a)對這9個miRNA篩選出7 963個潛在的靶基因,借助Cytoscape軟件構建了lncRNA- miRNA-mRNA靶向關系的網絡構建圖(圖8)。
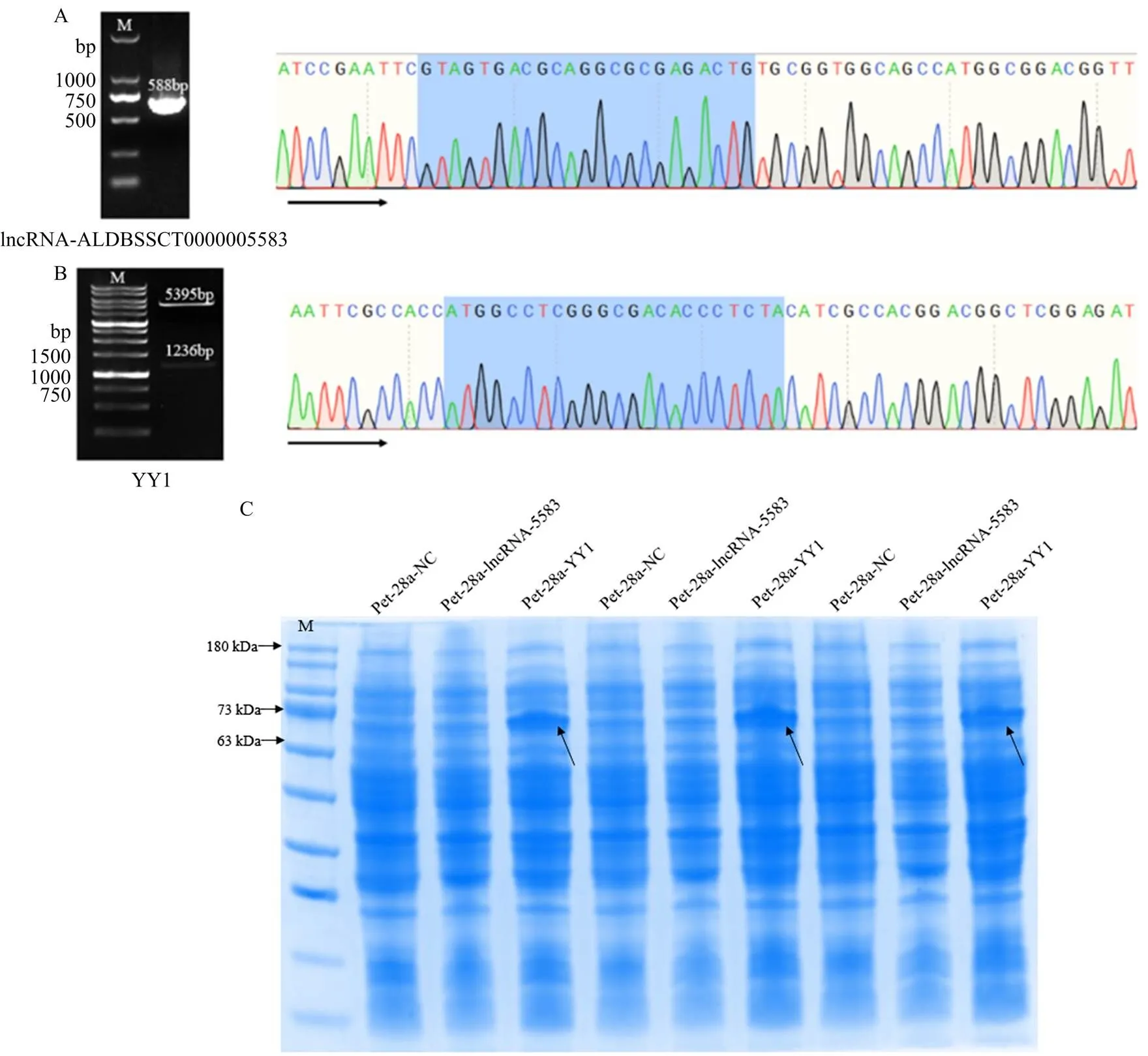
A:lncRNA- ALDBSSCT0000005583全長擴增和連接至pET-28a(+)載體Sanger DNA測序;B:pET-28a-YY1雙酶切結果及Sanger DNA測序;C:SDS-PAGE電泳分析

圖4 顆粒細胞的鑒定
查閱文獻發現miR-193a-5p[13]、miR-361-3p[14]可能與卵巢的發育有關。根據TargetScan(v5.0)和miRanda(v3.3a)分別對miR-193a-5p、miR-361-3p篩選出1 152、3 026個潛在的靶基因,對這兩個miRNA的靶基因求交集共獲得298個靶基因。對這些靶基因進行GO富集分析和KEGG信號通路分析,結果表明,這些miRNAs顯著富集到系統進程和細胞與細胞信號傳導等生物學過程中(表4)。KEGG信號通路分析表明,這兩個miRNA顯著參與到催產素信號通路、Ras信號通路、NF-κB信號通路、促性腺激素釋放激素分泌通路等(表5)。
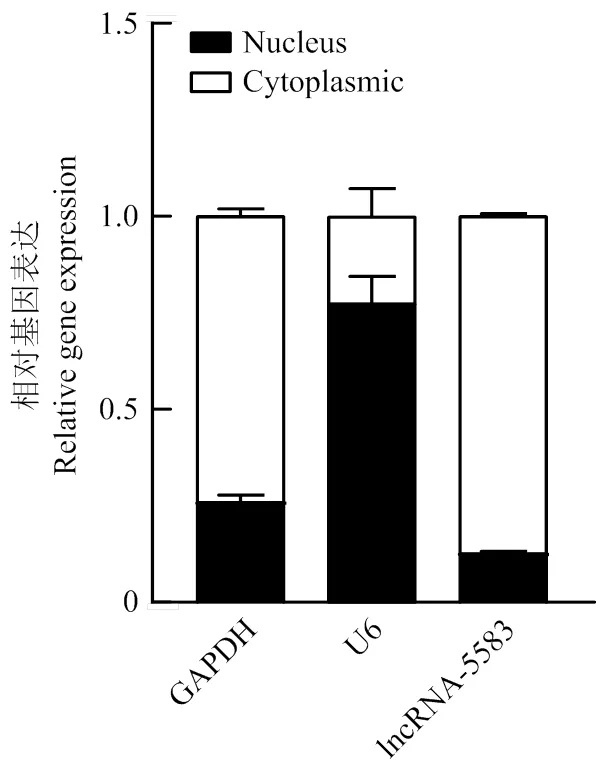
圖5 lncRNA- ALDBSSCT0000005583亞細胞定位
2.7 lncRNA-miRNA表達相關性分析
過表達lncRNA-ALDBSSCT0000005583有顯著的過表達效果(圖9-A),siRNA-ALDBSSCT0000005583有極顯著的抑制效果(圖9-B),結果表明過表達以及干擾lncRNA-ALDBSSCT0000005583的豬卵巢顆粒細胞模型構建成功,可以進行下一步試驗。
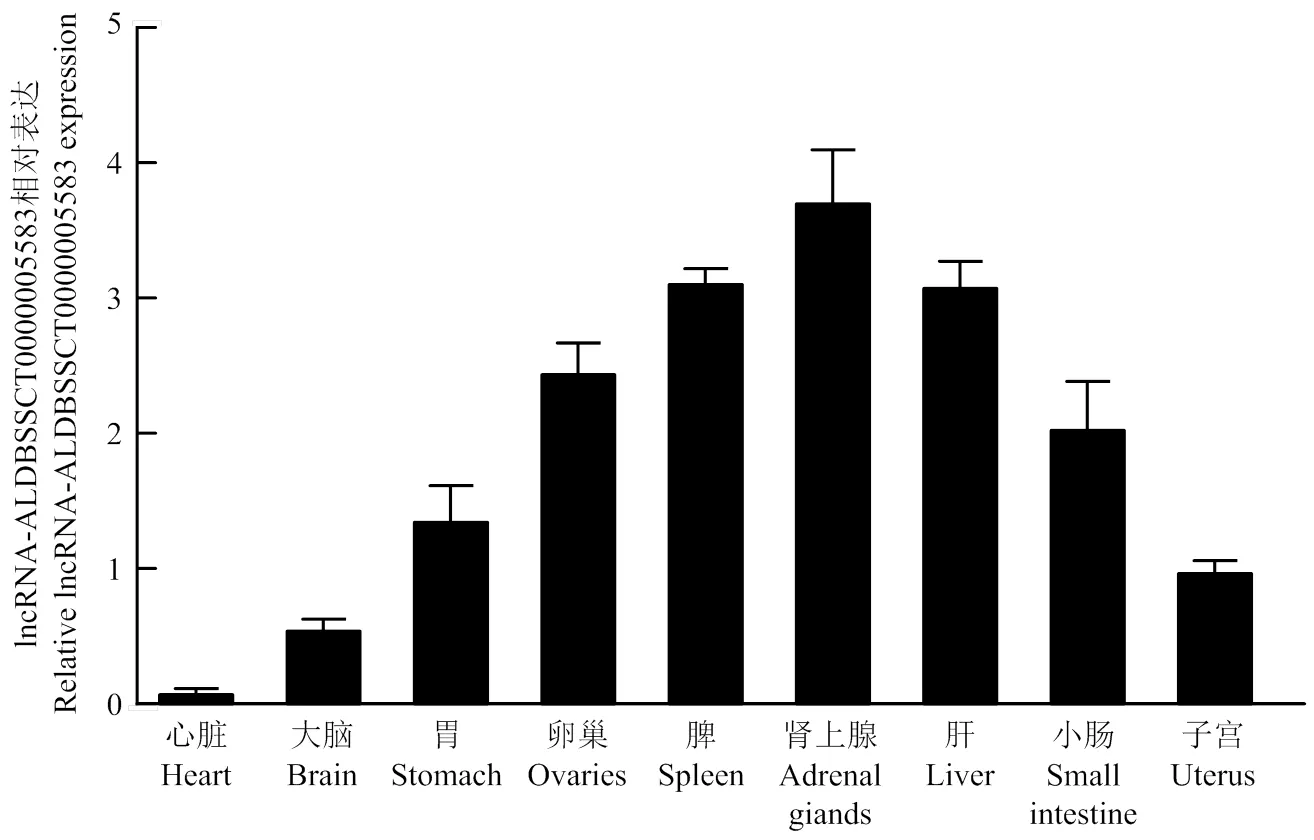
圖6 lncRNA-ALDBSSCT0000005583在杜洛克豬不同組織中的表達情況

圖7 RNAhybrid和miRanda預測miRNA靶向lncRNA-ALDBSSCT0000005583
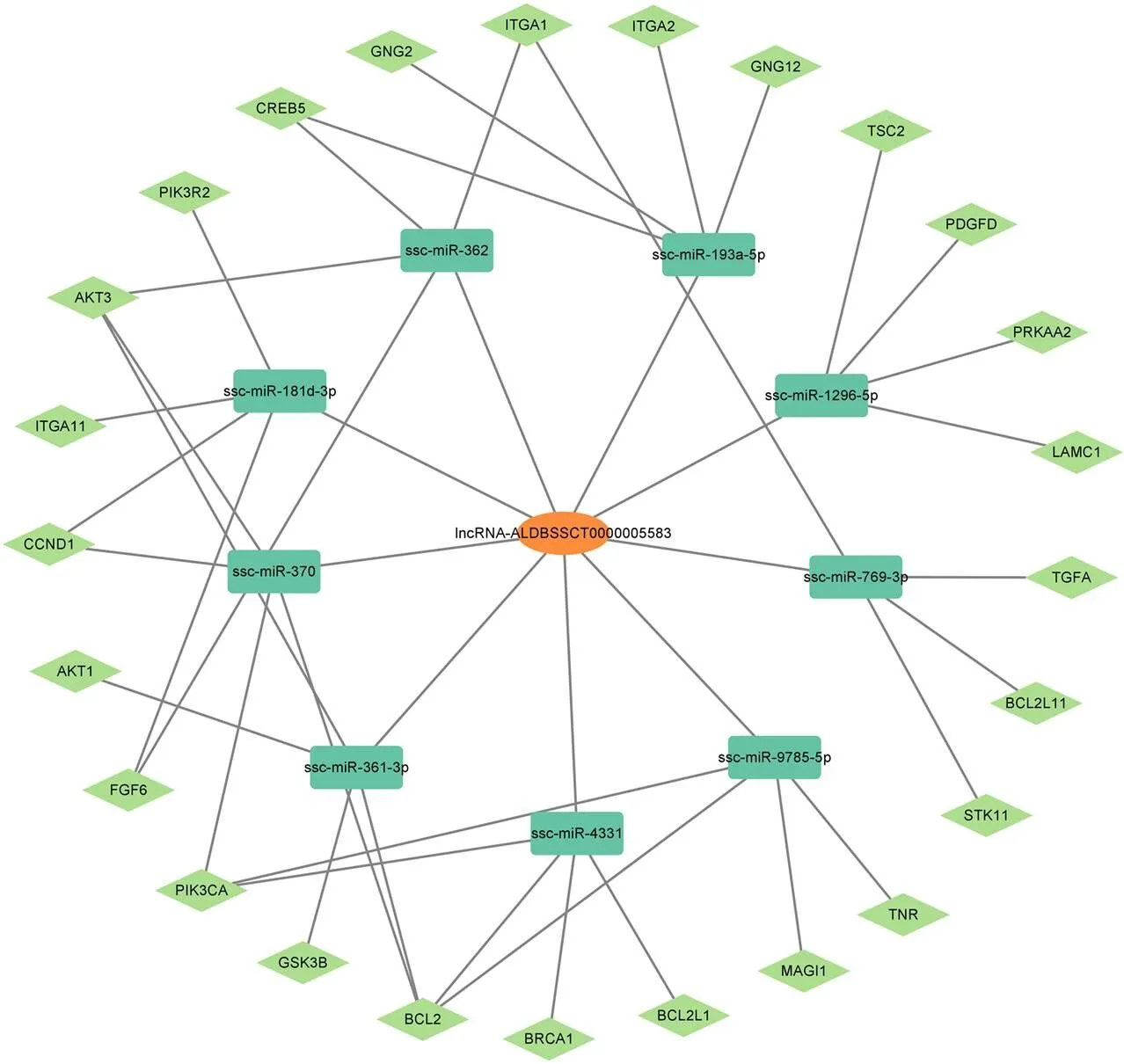
圖8 lncRNA-miRNA-mRNA網絡構建

表4 與lncRNA-ALDBSSCT0000005583相互作用的miRNAs靶基因的GO富集條目
過表達lncRNA后顯著降低了miR-193a-5p和miR-361-3p的表達量(圖9-C、E),但干擾lncRNA后miR-193a-5p和miR-361-3p的表達量有升高的趨勢但不顯著(圖9-D、F),以上研究表明,lncRNA- ALDBSSCT0000005583在豬的卵巢顆粒細胞中可能通過調控miR-193a-5p、miR-361-3p基因的表達,從而參與卵泡的發育過程。
3 討論
3.1 lncRNA-ALDBSSCT0000005583組織表達分析
母豬生產力作為影響生產效率的最重要因素之一,限制窩產仔豬數的主要因素是排卵率,排卵率越高,產仔數越多[12,15]。相對于杜洛克母豬,梅山豬的產仔數更大,以其高繁殖力聞名[16]。相關研究發現,多產的梅山母豬和雜種大白母豬在卵泡期的中后期存在卵泡生長動力學差異,這種差異極大地促進了排卵率[17]。此外,另一項研究表明,梅山母豬和杜洛克母豬在中等大小卵巢卵泡生長調節和生理發育方面也存在顯著差異[18]。目前在動物卵巢中的功能和分子機制的研究發現lncRNA對卵泡的生長發育至關重要。但是關于豬卵巢lncRNA的研究主要集中于篩選出卵巢中差異表達的lncRNA,并未進行下一步驗證。因此本研究通過RACE技術獲取并鑒定了梅山豬和杜洛克豬M2卵泡中差異表達的lncRNA-ALDBSSCT0000005583,其全長588 bp,經原核表達試驗驗證,lncRNA-ALDBSSCT0000005583不具備編碼蛋白的能力,是一個真正的長鏈非編碼RNA。中樞神經系統中的神經元對腎上腺和卵巢具有跨神經支配[19],并且在母羊卵巢摘除后,其腎周圍脂肪組織中生長激素受體(growth hormone receptor,GHR)表達量增加[20]。根據前人研究結果發現腎上腺和卵巢之間存在密不可分的關系。本研究通過組織表達譜分析,發現lncRNA在腎上腺中表達量最高,其次為肝、脾和卵巢,表明lncRNA-ALDBSSCT0000005583在這些組織中可能均具備生物學功能。
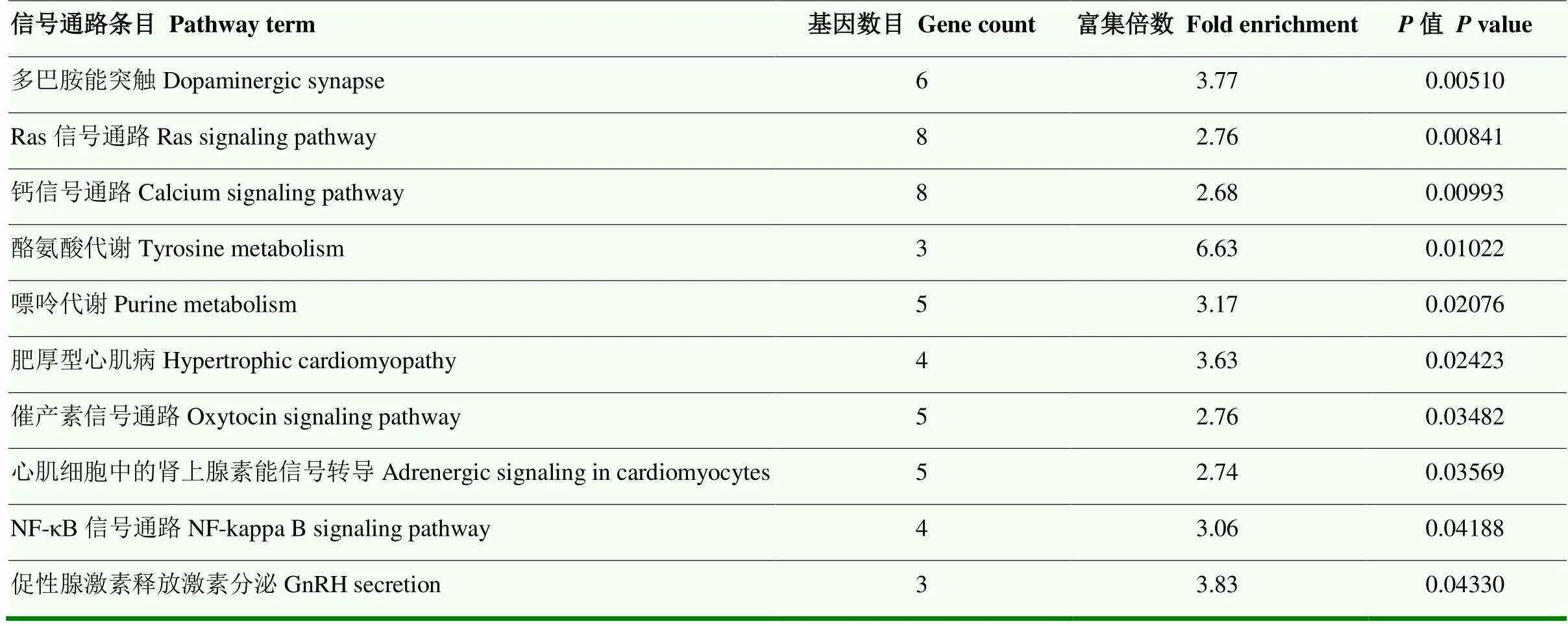
表5 與lncRNA-ALDBSSCT0000005583相互作用的miRNAs靶基因的信號通路
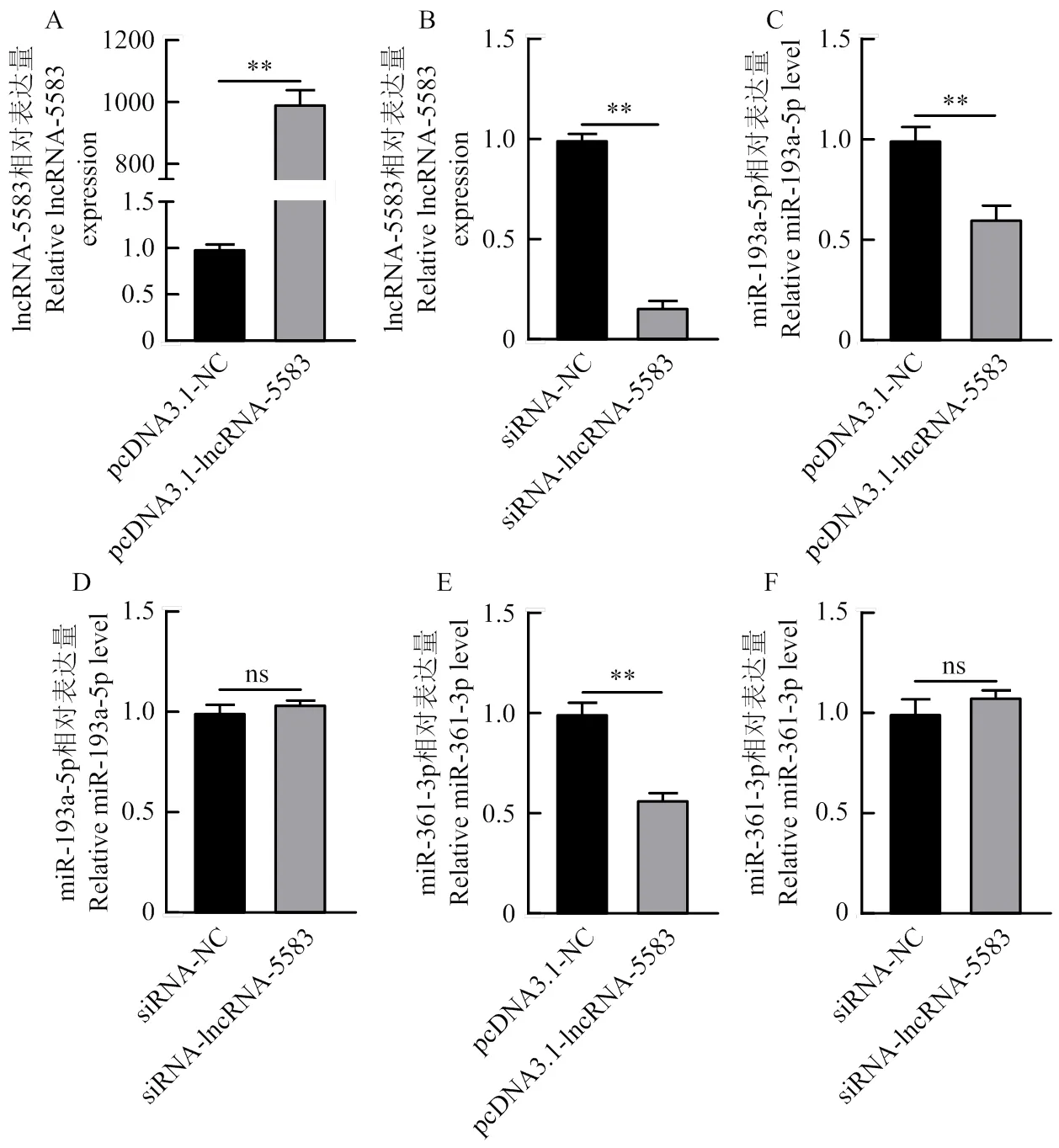
A. 過表達lncRNA-ALDBSSCT0000005583效率檢測;B. siRNA-ALDBSSCT0000005583效率檢測;C. 過表達lncRNA-ALDBSSCT0000005583對miR-193a-5p的影響;D. 過表達lncRNA- ALDBSSCT0000005583對miR-193a-5p的影響;E. 干擾lncRNA-ALDBSSCT0000005583對miR-361-3p的影響;F. 干擾lncRNA- ALDBSSCT0000005583對miR-1361-3p的影響
3.2 lncRNA-ALDBSSCT0000005583-miRNA-mRNA網絡分析
miRNA和lncRNA通過識別序列相互作用,其中lncRNA充當競爭性內源RNA(ceRNA)導致miRNA的隔離,并抑制miRNA對靶mRNA的調節作用[21-22]。ceRNA機制通常發生在細胞質中,這與miRNA在細胞質中起作用的觀點一致[23]。本研究通過核質分離試驗分析了lncRNA-ALDBSSCT0000005583的細胞定位,發現lncRNA主要存在于細胞質中,為了研究lncRNA-ALDBSSCT0000005583是否可以作為ceRNA或miRNA的海綿發揮作用,我們使用生物信息學靶基因預測工具搜索了可以與lncRNA結合的miRNA,總共篩選到9個與lncRNA相互作用的miRNAs。其中,有研究表明miR-193a-5p可以調控RAB11A[13]、HOXA7[24]和CDK14[25]抑制卵巢癌細胞的增殖并促進其凋亡;miR-361-3p可以抑制卵泡刺激素(FSH)的分泌[14],并且miR-361-3p也可以作為LncRNA-BBOX1-AS1[26]、LINC00922[27]的海綿對卵巢的生長發育起到一定的調控作用。結合前人對miRNAs功能的研究篩選出兩個與卵巢發育相關的miRNA:miR-193a-5p和miR-361-3p。對這兩個miRNAs的靶基因通過GO富集和KEGG信號通路表明:這些miRNAs顯著富集到系統進程和細胞與細胞信號傳導等生物學過程中。KEGG信號通路分析表明,這兩個miRNA顯著參與到催產素信號通路、Ras信號通路、NF-κB信號通路等。
3.3 lncRNA-ALDBSSCT0000005583與miRNA相關性分析
眾所周知,miRNA通過與靶mRNA的3′-非翻譯區(3′UTR)結合,成為基因調控的主要參與者,隨后引起mRNA降解或翻譯抑制[28],miRNA也對卵泡發育具有一定的影響[29]。與miRNA結合的lncRNA具有極大的研究價值,因為它們可用于調控miRNA功能。到目前為止,只有少數lncRNA被發現與miRNA結合,這些lncRNA已經過驗證。例如,lncRNA-ZFAS1與miR-129結合進而影響卵巢顆粒細胞的增殖和凋亡[30],lncRNA-NEAT1可以阻止miR-16、miR-483和miR-324-3p的表達來促進卵巢顆粒細胞的增殖并阻止細胞凋亡[31],lncRNA-2300通過抑制miR-365-3p的促凋亡作用來減少豬顆粒細胞的凋亡[32],lncRNA- FDNCR靶向湖羊的miR-543-3p來促進顆粒細胞凋亡[33],lncRNA-MEG3通過負反饋調控miR-23影響卵泡顆粒細胞的凋亡[34]。根據ceRNA假說,lncRNA應與miRNA表達呈負相關,lncRNA下調miRNA的表達水平,降低其對mRNA的抑制作用[35]。本研究通過過表達以及干擾lncRNA發現miR-193a-5p和miR- 361-3p的表達與lncRNA存在著顯著的負相關。由此可以推測,lncRNA可能調控miR-193a-5p、miR-361-3p參與到卵巢的生長發育,但是其具體的作用機制還需進一步后續驗證。
4 結論
lncRNA-ALDBSSCT0000005583在杜洛克豬M2卵泡中的表達量極顯著高于梅山豬,是一個全長588 bp的不具有翻譯蛋白質功能的lncRNA,生物信息學分析發現lncRNA-ALDBSSCT0000005583可能與系統進程和細胞與細胞信號傳導等生物學過程有關;并參與到催產素、Ras、NF-κB、促性腺激素釋放激素分泌通路等通路;并且可能對miR-193a-5p、miR-361-3p的表達存在影響。本研究為進一步探索lncRNA- ALDBSSCT0000005583在豬卵巢的生長發育過程中提供了基礎數據。
[1] GOLICZ A A, BHALLA P L, SINGH M B. lncRNAs in plant and animal sexual reproduction. Trends in Plant Science, 2018, 23(3): 195-205.
[2] KHALIL A M, GUTTMAN M, HUARTE M, GARBER M, RAJ A, RIVEA MORALES D, THOMAS K, PRESSER A, BERNSTEIN B E, VAN OUDENAARDEN A, REGEV A, LANDER E S, RINN J L. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(28): 11667-11672.
[3] LEE J T. Epigenetic regulation by long noncoding RNAs. Science, 2012, 338(6113): 1435-1439.
[4] TSAI M C, MANOR O, WAN Y, MOSAMMAPARAST N, WANG J K, LAN F, SHI Y, SEGAL E, CHANG H Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science, 2010, 329(5992): 689-693.
[5] RINN J L, CHANG H Y. Genome regulation by long noncoding RNAs. Annual Review of Biochemistry, 2012, 81: 145-166.
[6] HU S S, WANG X L, SHAN G. Insertion of an Alu element in a lncRNA leads to primate-specific modulation of alternative splicing. Nature Structural & Molecular Biology, 2016, 23: 1011-1019.
[7] LIU S J, DANG H X, LIM D A, FENG F Y, MAHER C A. Long noncoding RNAs in cancer metastasis. Nature Reviews Cancer, 2021, 21(7): 446-460.
[8] YOUNG L D. Effects of Duroc, Meishan, Fengjing, and Minzhu boars on productivity of mates and growth of first-cross progeny. Journal of Animal Science, 1992, 70(7): 2020-2029.
[9] FOXCROFT G R, HUNTER M G. Basic physiology of follicular maturationin the pig. Journal of Reproduction and Fertility, 1985, (Suppl 33): 1-19.
[10] LI M X, LIU Y, XIE S, MA L P, ZHAO Z C, GONG H B, SUN Y S, HUANG T. Transcriptome analysis reveals that long noncoding RNAs contribute to developmental differences between medium-sized ovarian follicles of Meishan and Duroc sows. Scientific Reports, 2021, 11: 22510.
[11] HU H Y, JIA Q, XI J Z, ZHOU B, LI Z Q. Integrated analysis of lncRNA, miRNA and mRNA reveals novel insights into the fertility regulation of large white sows. BMC Genomics, 2020, 21(1): 636.
[12] LIU Y, LI M X, BO X W, LI T, MA L P, ZHAI T, HUANG T. Systematic analysis of long non-coding RNAs and mRNAs in the ovaries of duroc pigs during different follicular stages using RNA sequencing. International Journal of Molecular Sciences, 2018, 19(6): 1722.
[13] WANG Y Z, LI N, ZHAO J, DAI C W. MiR-193a-5p serves as an inhibitor in ovarian cancer cells through RAB11A. Reproductive Toxicology, 2022, 110: 105-112.
[14] YE R S, LI M, LI C Y, QI Q E, CHEN T, CHENG X, WANG S B, SHU G, WANG L N, ZHU X T, JIANG Q Y, XI Q Y, ZHANG Y L. MiR-361-3p regulates FSH by targeting FSHB in a porcine anterior pituitary cell model. Reproduction, 2017, 153(3): 341-349.
[15] LIU J Y, QI N N, XING W W, LI M X, QIAN Y H, LUO G, YU S L. The TGF-β/SMAD signaling pathway prevents follicular atresia by upregulating MORC2. International Journal of Molecular Sciences, 2022, 23(18): 10657.
[16] ZHENG X R, ZHAO P J, YANG K J, NING C, WANG H F, ZHOU L, LIU J F. CNV analysis of Meishan pig by next-generation sequencing and effects ofgene CNV on pig reproductive traits. Journal of Animal Science and Biotechnology, 2020, 11: 42.
[17] MILLER A T, PICTON H M, CRAIGON J, HUNTER M G. Follicle dynamics and aromatase activity in high-ovulating Meishan sows and in large-white hybrid contemporaries. Biology of Reproduction, 1998, 58(6): 1372-1378.
[18] MA L P, ZHAO Z C, LI T, LI D Q, WANG X Y, SONG C Y, QI Y Y, HUANG T. Identification of differentially expressed microRNAs in middle-size ovarian follicles of Meishan and Duroc sows. Revista Brasileira De Zootecnia, 2019, 48. doi:10.1590/rbz4820170326.
[19] TóTH I E, BANCZEROWSKI P, BOLDOGK?I Z, TóTH J S, SZABó A, HALáSZ B, GERENDAI I. Cerebral neurons involved in the innervation of both the adrenal gland and the ovary: a double viral tracing study. Brain Research Bulletin, 2008, 77(5): 306-311.
[20] 張磊, 王燕燕, 周占琴, 李廣, 付明哲, 張鎖良, 尹海科, 張勝剛, 任寶華. 卵巢摘除對山羊血清GH水平和組織中GHR基因表達的影響. 中國農業科學, 2014, 47(1): 199-208. doi: 10.3864/j.issn.0578- 1752.2014.10.003.
ZHANG L, WANG Y Y, ZHOU Z Q, LI G, FU M Z, ZHANG S L, YIN H K, ZHANG S G, REN B H. Effects of ovariectomy on serum GH levels and expression of GHR in some tissues of female goats. Scientia Agricultura Sinica, 2014, 47(1): 199-208. doi: 10.3864/j.issn. 0578-1752.2014.10.003.(in Chinese)
[21] HAN T S, HUR K, CHO H S, BAN H S. Epigenetic associations between lncRNA/circRNA and miRNA in hepatocellular carcinoma. Cancers, 2020, 12(9): 2622.
[22] WANG H, HUO X S, YANG X R, HE J, CHENG L J, WANG N, DENG X, JIN H J, WANG N, WANG C, ZHAO F Y, FANG J Y, YAO M, FAN J, QIN W X. STAT3-mediated upregulation of lncRNA HOXD-AS1 as a ceRNA facilitates liver cancer metastasis by regulating SOX4. Molecular Cancer, 2017, 16(1): 136.
[23] HUARTE M. The emerging role of lncRNAs in cancer. Nature Medicine, 2015, 21: 1253-1261.
[24] WANG S, DIAO Y J, ZHU B B. MiR-193a-5p suppresses cell proliferation and induces cell apoptosis by regulating HOXA7 in human ovarian cancer. Neoplasma, 2020, 67(4): 825-833.
[25] 楊尊敬, 杜先玲. 細胞周期蛋白依賴性激酶在miR-193a5p調控卵巢癌細胞增殖及上皮細胞間充質轉變中的作用. 中國應用生理學雜志, 2020, 36(2): 176-180.
YANG Z J, DU X L. The role of cyclin-dependent kinases in miR-193a5p regulating ovarian cancer cell proliferation and epithelial mesenchymal transformation. Chinese Journal of Applied Physiology, 2020, 36(2): 176-180. (in Chinese)
[26] YAO H P, CHEN R, YANG Y X, JIANG J. LncRNA BBOX1-AS1 aggravates the development of ovarian cancer by sequestering miR-361-3p to augment PODXL expression. Reproductive Sciences, 2021, 28(3): 736-744.
[27] WANG L P, REN C C, XU Y J, YANG L, CHEN Y N, ZHU Y H. The LINC00922 aggravates ovarian cancer progression via sponging miR-361-3p. Journal of Ovarian Research, 2021, 14(1): 77.
[28] BARTEL D P. MicroRNAs: Target recognition and regulatory functions. Cell, 2009, 136(2): 215-233.
[29] 陳慧芳, 黃綺亮, 胡智超, 潘曉婷, 吳志勝, 白銀山. 外泌體microRNA在豬成熟和閉鎖卵泡中的表達差異及功能分析. 中國農業科學, 2021, 54(21): 4664-4676. doi: 10.3864/j.issn.0578-1752. 2021.21.015.
CHEN H F, HUANG Q L, HU Z C, PAN X T, WU Z S, BAI Y S. Expression differences and functional analysis of exosomes microRNA in porcine mature and atretic follicles. Scientia Agricultura Sinica, 2021, 54(21): 4664-4676. doi: 10.3864/j.issn.0578-1752.2021.21.015.(in Chinese)
[30] ZHU H L, CHEN Y Q, ZHANG Z F. Downregulation of lncRNA ZFAS1 and upregulation of microRNA-129 repress endocrine disturbance, increase proliferation and inhibit apoptosis of ovarian granulosa cells in polycystic ovarian syndrome by downregulating HMGB1. Genomics, 2020, 112(5): 3597-3608.
[31] Sang X , Zhang Y Z. Withdrawal: “long non-coding RNA NEAT1 drives the development of polycystic ovary syndrome via sponging multiple microRNAs” by Xia Sang and Yuzhen Zhang. Cell Biology International, 2022, 46(7): 1175.
[32] WANG M M, WANG Y, YAO W, DU X, LI Q F. Lnc2300 is a cis-acting long noncoding RNA of CYP11A1 in ovarian granulosa cells. Journal of Cellular Physiology, 2022, 237(11): 4238-4250.
[33] YAO X L, GAO X X, BAO Y J, EL-SAMAHY M A, YANG J Y, WANG Z B, LI X D, ZHANG G M, ZHANG Y L, LIU W J, WANG F. lncRNApromotes apoptosis of granulosa cells by targeting the miR-543-3p/DCN/TGF-β signaling pathway in Hu sheep. Molecular Therapy - Nucleic Acids, 2021, 24: 223-240.
[34] HAN X H, PAN Y Y, FAN J F, WANG M, WANG L B, WANG J L, AFEDO S Y, ZHAO L, WANG Y Y, ZHAO T, ZHANG T X, ZHANG R, CUI Y, YU S J.regulatesaxis-mediated apoptosis and autophagy via spongingin granulosa cells of yak tertiary follicles. Cellular Signalling, 2023, 107: 110680.
[35] TANG Y L, ZHANG H, CHEN L L, ZHANG T M, XU N, HUANG Z N. Identification of hypoxia-related prognostic signature and competing endogenous RNA regulatory axes in hepatocellular carcinoma. International Journal of Molecular Sciences, 2022, 23(21): 13590.
Cloning and Identification of Differentially Expressed lncRNAs in Follicles of Meishan Pigs and Duroc Pigs with Their Correlation Analysis with miRNAs
ZHANG HuaPeng1, ZHANG QingZe1, HE Fan1, QI MengFan1, FU BinBin1, LI QingChun1, LI MengXun1, MA LiPeng1, LIU Yi1, HUANG Tao1, 2
1College of Animal Science and Technology, Shihezi University, Shihezi 832000, Xinjiang;2Xinjiang Pig Seed Industry Engineering Technology Research Center, Changji 831100, Xinjiang
【Objective】 The objective of this study was to clone and identify the differentially expressed lncRNA-ALDBSSCT0000005583in M2 follicles on the fourth day of follicular stage in Meishan pigs and Duroc pigs, and to analyze the correlation between the expression of miRNAs in porcine granulosa cells, so as to provide a theoretical basis for exploring the role of lncRNAs in the development of follicles in sows by regulating miRNAs. 【Method】Based on the differentially expressed lncRNA- ALDBSSCT0000005583 screened in Meishan and Duroc M2 follicles in our early research, the full-length sequence of ALDBSSCT0000005583 was verified by RT-qPCR and cloned by RACE; the coding ability of this lncRNA was predicted by the coding potential assessment tool of CAPT and CPC, which was further identified by the primary expression test; the coding ability of this lncRNA was identified by the primary expression test; the subcellular coding ability of NA-ALDBSSCT0000005583 was identified by the nucleoplasmic separation experiment and tested to identify its coding ability; the subcellular localization of lncRNA-ALDBSSCT0000005583 by nucleoplasmic isolation assay and its expression level in various tissues were detected by RT-qPCR; miRBase website was used to locate the miRNA database of pigs, and the combination of RNAhybrid and miRanda online software was used to predicte the relationship with the lncRNA-ALDBSSCT0000005583. The inter-species conserved miRNAs that interacted with lncRNA-ALDBSSCT0000005583 were predicted by TargetScan and miRanda, the target genes that interacted with lncRNA-ALDBSSCT0000005583 were predicted by TargetScan and miRanda, and their target genes were subjected to GO enrichment and KEGG signaling pathway analyses; the effects of target genes on miRNA expression were verified by overexpression as well as interference with lncRNA. 【Result】The expression level of lncRNA-ALDBSSCT0000005583 in M2 follicles of Duroc pigs was significantly higher than that in Meishan pigs, and the size of lncRNA 5′RACE and 3′RACE fragments was 569 bp and 546 bp, respectively, and the sequencing analysis showed that the size of lncRNA-ALDBSSCT0000005583 was 588 bp. Bioinformatics predicted that the encoding potential was low, and the results of prokaryotic expression assay further proved that it did not code for proteins. Tissue expression profiling showed that lncRNA-ALDBSSCT0000005583 was expressed in the adrenal gland, spleen, liver and ovaries, and low in the hypothalamus and heart, while the subcellular localization results showed that the lncRNA was mainly present in the cytoplasm. After bioinformatics analysis, a total of 9 conserved miRNAs were screened for potential interaction with lncRNA-ALDBSSCT0000005583, including two miRNAs related to ovarian development: miR-193a-5p and miR-361-3p. KEGG and GO enrichment analysis showed that the target genes of miR-193a-5p and miR-361-3p were related to phylogenetic processes and biological processes such as cell-to-cell signaling. It was also significantly involved in oxytocin, Ras, NF-κB gonadotropin- releasing hormone secretion and other pathways. Subsequently, lncRNA-ALDBSSCT0000005583 was overexpressed in granulosa cells, and the expressions of miR-193a-5p and miR-361-3p were significantly down-regulated by RT-qPCR (<0.05), but there was no significant effect after interference with lncRNA-ALDBSSCT0000005583. 【Conclusion】lncRNA-ALDBSSCT0000005583 was a lncRNA that did not have the ability to code for proteins. There was a significant difference in the expression level between the medium follicles of Meishan and Duroc pigs, and the expression level was higher in the adrenal glands, spleen, liver and ovaries, which is mainly found in the cytoplasm of granulosa cells, and might be involved in the development of porcine ovarian granulosa cells by interacting with miR-193a-5p and miR-361-3p.
lncRNA-ALDBSSCT0000005583; pig; follicle; granulosa cells; miR-193a-5p; miR-361-3p

10.3864/j.issn.0578-1752.2024.09.014
2023-11-06;
2024-01-11
國家自然科學基金(31960645)、新疆維吾爾自治區天山英才青年科技拔尖人才項目(2022TSYCCX0047)、新疆維吾爾自治區第七師胡楊河市“揭榜掛帥”項目(QS2023010)
張化鵬,E-mail:914566978@qq.com。通信作者黃濤,E-mail:taohuang100@sina.com
(責任編輯 林鑒非)
猜你喜歡
黃河之聲(2019年1期)2019-03-30 03:38:06
少兒美術(快樂歷史地理)(2018年4期)2018-09-25 02:48:54
黃河之聲(2018年6期)2018-05-18 00:47:57
作文新天地(2016年18期)2016-02-13 01:15:01
中國資源綜合利用(2016年5期)2016-02-03 02:56:17
邵陽學院學報(社會科學版)(2015年3期)2015-02-28 16:18:23
水道港口(2014年1期)2014-04-27 14:14:35
電力需求側管理(2014年3期)2014-03-20 13:35:43
水道港口(2013年3期)2013-04-27 14:14:30
湖南人文科技學院學報(2012年6期)2012-04-16 03:32:32
