巖藻多糖降解酶的研究進展
2024-05-20 07:17:40陳金玲
食品科學 2024年9期
魏 真,陳金玲,楊 杰
(1.江蘇海洋大學 江蘇省海洋生物資源與環(huán)境重點實驗室,江蘇 連云港 222005;2.江蘇省海洋生物產(chǎn)業(yè)技術協(xié)同創(chuàng)新中心,江蘇 連云港 222005;3.江蘇省海洋資源開發(fā)研究院,江蘇 連云港 222005)
巖藻多糖降解酶是一類特異性作用于巖藻多糖中糖苷鍵的水解酶,能夠?qū)⒋蠓肿訋r藻多糖水解為中、低分子質(zhì)量的巖藻聚糖[1]。多種巖藻低聚糖具有免疫調(diào)節(jié)、抗腫瘤、抗氧化、美白等特殊的生物學功效[2-5],因而巖藻多糖降解酶成為生產(chǎn)巖藻低聚糖不可或缺的工具酶。除了制備巖藻低聚糖之外,巖藻多糖降解酶還可用于功能性食品研發(fā)、疾病診斷以及推斷復雜巖藻多糖的化學結構等,因此巖藻多糖降解酶在食品、醫(yī)藥等領域有著較高的應用價值。近年來,隨著功能性食品與保健品市場需求的擴大,巖藻多糖降解酶的特殊作用與水解產(chǎn)物的功效也逐漸被重視和研究。由于巖藻多糖的分子結構極為復雜,且不同功效的巖藻低聚糖與其分子質(zhì)量、硫酸基的含量和分子組成密切相關[6-8],因此深入探討不同來源和功能的巖藻多糖降解酶對于開發(fā)和研究多種生理活性的巖藻低聚糖具有十分重要的意義。本文總結近年來不同生物來源的巖藻多糖降解酶的研究進展,從巖藻多糖降解酶的分類、酶學性質(zhì)、作用方式、結構功能及應用領域等方面展開論述,以期為后續(xù)巖藻多糖降解酶的理論研究及應用提供參考。
1 巖藻多糖降解酶的分類和來源
巖藻多糖降解酶歸屬于糖苷水解酶(glucoside hydrolase,GH)類,能夠特異性作用于巖藻多糖中的糖苷鍵[1]。巖藻多糖是一種來源于海洋褐藻及海洋無脊椎動物的硫酸化多糖,其主鏈主要由L-巖藻糖組成,側(cè)鏈分布著少量不同含量的甘露糖、木糖、半乳糖和葡萄糖醛酸等組分[9-10]。研究表明,巖藻多糖的分子結構極為復雜多樣,從不同來源、不同生長季節(jié)褐藻中提取的巖藻多糖化學成分和結構均不相同[8],甘露糖、木糖等單糖組分被替換在L-巖藻糖基的隨機位置,巖藻多糖的硫酸化位點可發(fā)生在L-巖藻糖基的C2、C3或C4等位置[11]。作為能夠水解復雜巖藻多糖的重要酶,巖藻多糖降解酶具有多樣性,不同分類和來源的巖藻多糖降解酶正被不斷地挖掘和認識。
“巖藻多糖降解酶”一詞最早出現(xiàn)在1959年發(fā)表于Nature上的一篇從海洋細菌Pseudomonasatlantica和Pseudomonascarrageenovora中分離的酶能夠降解巖藻多糖的研究中,該酶被命名為“fucoidinase”[12]。直到1967年,有研究報道從鮑魚肝胰腺分離的酶能夠催化巖藻多糖降解[13],同時這種降解巖藻多糖的生物酶被更名為“fucoidanase”并沿用至今。隨著研究者對巖藻多糖降解酶重要性的認識,該酶也在其他某些無脊椎動物的內(nèi)臟和海洋微生物中被發(fā)現(xiàn)。目前巖藻多糖降解酶主要包含3 種酶:巖藻多糖酶(fucoidanase)、α-L-巖藻糖苷酶(fucosidase)和硫酸酯酶(sulfatase)。
1.1 巖藻多糖酶
巖藻多糖酶(EC 3.2.1.211,EC 3.2.1.212)是目前關于降解巖藻多糖研究最多的一種酶,它能夠催化巖藻多糖中α-糖苷鍵的水解[14]。還有β-D-巖藻糖苷酶(EC 3.2.1.38),由于在巖藻多糖中并未發(fā)現(xiàn)D型巖藻糖,因而其不能作用于巖藻多糖。根據(jù)水解位置不同,巖藻多糖酶分為內(nèi)切型和外切型。內(nèi)切巖藻多糖酶可以切割巖藻多糖分子內(nèi)的糖苷鍵,產(chǎn)生不同分子質(zhì)量的巖藻低聚糖,而外切巖藻多糖酶能夠?qū)r藻多糖的非還原端水解為非硫酸化或者硫酸化的巖藻糖分子[1]。巖藻多糖酶分布十分廣泛,在一些海洋微生物及無脊椎動物中均可分離和鑒定到該酶的存在。目前已報道產(chǎn)巖藻多糖降解酶的細菌主要有依氏假交替單胞菌(Pseudoalteromonasissachenkoniisp.nov.)[15]、交替單胞菌屬(Alteromonassp.)[16]、昆布海藻鼠尾菌(Muricaudaeckloniae)[17]、少動鞘氨醇單胞菌(SphingomonaspaucimobilisPF-1)[18]、鮑魚美麗菌(Formosahaliotis)[19]、解巖藻聚糖莊文穎菌(Wenyingzhuangiafucanilytica)[20]、黃桿菌屬(Flavobacteriumsp.)[21]和海綿科貝特氏菌(Cobetia amphilecti)[22]等。此外,從真菌和無脊椎動物的內(nèi)臟也能夠分離出巖藻多糖降解酶,例如真菌來源的鐮刀菌屬(Fusariumsp.LD8)[23]、砂生小樹狀霉(Dendryphiella arenariaTM94)[24]、謝瓦氏曲霉(Aspergilluschevalier)[25]等,以及無脊椎動物來源的鮑屬[13]、蝦夷盤扇貝[26]、光棘球海膽[27]和濱螺[28]等。
1.2 α-L-巖藻糖苷酶
α-L-巖藻糖苷酶(EC 3.2.1.51,EC 3.2.1.63,EC 3.2.1.111,EC 3.2.1.127)專一作用于巖藻多糖及巖藻低聚糖末端的α-L-巖藻糖基,由α-1,2、α-1,3、α-1,4或α-1,6連接的α-巖藻糖苷鍵[29]。α-L-巖藻糖苷酶絕大多數(shù)分離于海洋細菌,例如硫磺礦硫化葉菌(Sulfolobussolfataricus)[30]、解巖藻聚糖莊文穎菌(W.fucanilytica)[31]、類芽孢桿菌屬(Paenibacillussp.)[32]、蠟樣芽胞桿菌(Bacilluscereus)[33]和鼠李糖乳酪桿菌(LacticaseibacillusrhamnosusINIA P603)[34]等。少數(shù)巖藻糖苷酶來源于海洋無脊椎動物,例如海燕海星[35]等。
1.3 硫酸酯酶
硫酸酯酶(EC 3.1.6)以催化巖藻多糖中的氨基磺酸鹽和硫酸酯的水解為主[36],對于巖藻多糖硫酸酯酶的研究還不多。目前發(fā)現(xiàn)能夠催化巖藻多糖降解的硫酸酯酶主要來源于海洋細菌,例如解巖藻聚糖莊文穎菌(W.fucanilyticaCZ1127T)[37]、基里蒂瑪?shù)倬鷮伲↘iritimatiellalessp.)[38]、假交替單胞菌屬(Pseudoalteromonassp.)[39]等。
2 巖藻多糖降解酶的酶學性質(zhì)
由于巖藻多糖降解酶的種類及來源十分廣泛,其理化性質(zhì)也存在著多樣性,物種來源、酶的分子質(zhì)量、環(huán)境溫度、pH值及一些金屬離子等均為影響該酶性質(zhì)差異的重要因素。表1為部分物種來源巖藻多糖酶的基本酶學性質(zhì),其中研究者對分離于海洋細菌的巖藻多糖酶報道最多,且不同細菌來源巖藻多糖酶的分子質(zhì)量差異巨大。例如來源于C.amphilecti的巖藻多糖酶分子質(zhì)量為35 kDa[22],而從LuteolibacteralgaeH18分離得到的巖藻多糖酶分子質(zhì)量可達112 kDa[40]。細菌巖藻多糖酶一般為常溫酶,其最適作用溫度大多分布在24~35 ℃之間,但來自Flavobacteriumsp.SW的巖藻多糖酶最適溫度為45~50 ℃[21]。絕大多數(shù)細菌來源的巖藻多糖酶在pH值中性或弱堿性范圍內(nèi)均有較高的酶活性,其中分離于Flavobacteriumalgicola的巖藻多糖酶最適pH值為9.0,是目前發(fā)現(xiàn)耐受性最強的巖藻多糖酶[41]。與細菌來源的巖藻多糖酶相比,分離于無脊椎動物或海洋真菌的巖藻多糖酶在最適溫度和最適pH值等方面存在較大差異。目前發(fā)現(xiàn)的來源于海洋真菌的巖藻多糖酶最適溫度普遍高于無脊椎動物和海洋細菌,如分離于Fusariumsp.LD8的巖藻多糖酶具有耐熱性,其最適溫度達到了60 ℃[23]。不僅如此,分離于無脊椎動物和海洋真菌的巖藻多糖酶最適pH值均為酸性,其中來源于無脊椎動物光棘球海膽的巖藻多糖酶最適pH值僅為3.0,是典型的嗜酸酶[27]。此外,一些巖藻多糖酶的活性還受到金屬離子和無機鹽離子的影響,例如鈣離子能夠顯著增加巖藻多糖酶Mef2的活力,而抑制巖藻多糖酶Fda1活力[16-17];巖藻多糖酶Fhf1對氯化鈉溶液的最高耐受濃度為0.5 mol/L,巖藻多糖酶Fda1能夠在1 mol/L氯化鈉溶液中保持最佳酶活性[16]。
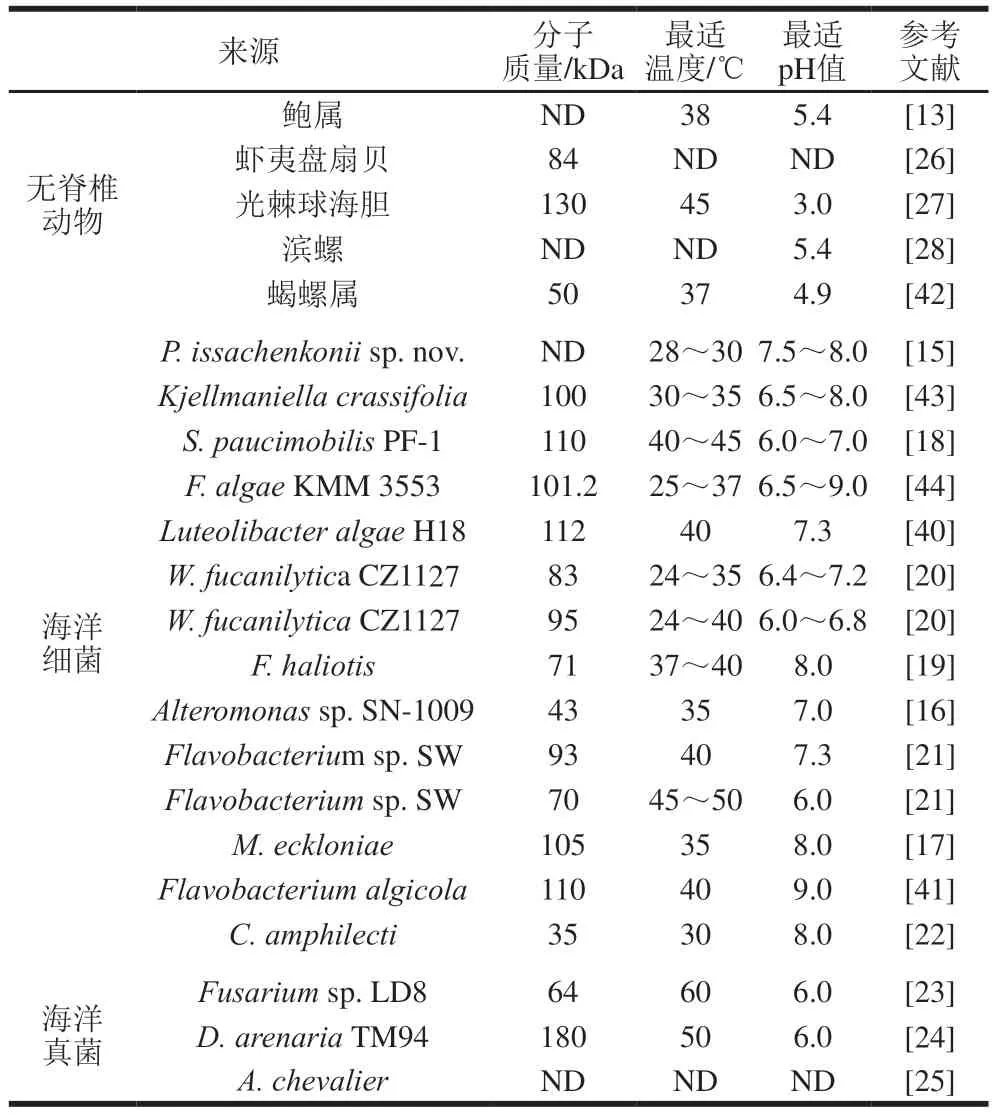
表1 不同來源巖藻多糖酶的酶學性質(zhì)Table 1 Enzymatic properties of fucoidanases from different sources
巖藻糖苷酶一般分離于海洋細菌,且多數(shù)使用大腸桿菌等對其進行異源重組表達和后續(xù)性質(zhì)研究。已有文獻報道的重組巖藻糖苷酶分子質(zhì)量差異較大,約在40~90 kDa之間。不同細菌來源的重組巖藻糖苷酶的最適溫度也不盡相同:幾種巖藻糖苷酶的最佳作用溫度約為50 ℃,例如分離于Paenibacillussp.3179、B.cereus2-8和L.rhamnosusINIA P603的巖藻糖苷酶[32-34],而來自W.fucanilytica的巖藻糖苷酶最適溫度僅為25~30 ℃[31]。絕大多數(shù)巖藻糖苷酶的最適pH值約在4.0~7.0之間,其能夠很好地適應酸性環(huán)境。少數(shù)分離于無脊椎動物的巖藻糖苷酶某些酶學性質(zhì)與細菌相比有較大區(qū)別,例如來源于海燕海星的巖藻糖苷酶最適溫度高達70 ℃,這在細菌巖藻糖苷酶中鮮有報道[35]。
有關巖藻多糖硫酸酯酶的研究相對較少,目前已報道的幾種硫酸酯酶在不同的溫度、金屬離子等環(huán)境中也表現(xiàn)出較大差異。例如Silchenko等[37]發(fā)現(xiàn)來源于菌株W.fucanilytica中的重組巖藻多糖硫酸酯酶SWF1和SWF4最適pH值均為7.8~8.4,最適溫度分別為40~45 ℃和30~35 ℃,且硫酸酯酶SWF1是一種鈣離子依賴性酶,而SWF4的活性不受鈣離子的影響。分離于海參腸道細菌的硫酸酯酶PsFucS1也具有鈣離子依賴性,其最適pH值為6.5,且具有高達68 ℃的最佳催化溫度和較高的熱穩(wěn)定性[39]。
3 巖藻多糖降解酶的作用方式和水解產(chǎn)物
酶最重要的特性是其多樣的作用位點、底物特異性與水解產(chǎn)物。巖藻多糖降解酶的作用底物主要為由L-巖藻糖組成的硫酸化多糖[9-10],L-巖藻糖的連接方式以α-1,3連接(圖1a)或α-1,3、α-1,4交替連接(圖1b)為主,此外還有少數(shù)α-1,2或α-1,6連接[9,45]。側(cè)鏈基團復雜多樣,主要由甘露糖、木糖、半乳糖和葡萄糖醛酸等組成[9]。圖1顯示了3 種巖藻多糖降解酶在部分巖藻多糖中的主要作用位點。由于巖藻多糖底物的復雜性與多樣性,不同種類的巖藻多糖降解酶具有不同的作用位點與水解方式,其水解產(chǎn)物也存在著較大差異。
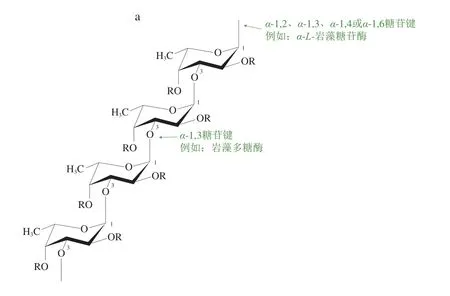
圖1 巖藻多糖的結構模型及巖藻多糖降解酶的一般作用位點[45]Fig.1 Structural models of fucoidans and general action sites of fucoidan-degrading enzymes[45]
巖藻多糖酶分為內(nèi)切型和外切型,兩種作用方式的酶能夠切割巖藻多糖中以α-1,3或α-1,4連接的巖藻糖苷鍵。大部分已發(fā)現(xiàn)的巖藻多糖酶為內(nèi)切型,其水解產(chǎn)物多為不同分子質(zhì)量的巖藻低聚糖,例如Zhu Chenlu等[16]使用高效液相色譜電噴霧離子化串聯(lián)質(zhì)譜對來自Alteromonassp.SN-1009的巖藻多糖酶tFda1B水解產(chǎn)物進行分析,結果表明該酶屬于內(nèi)切型α-1,3巖藻多糖酶,水解產(chǎn)物為多樣的巖藻低聚糖,且證實部分來自厚葉解曼藻的巖藻多糖是由α-1,3糖苷鍵連接。Tran等[17]通過核磁共振、底物特異性和系統(tǒng)聚類分析等對來源于糖海帶的巖藻多糖酶Mef2的水解產(chǎn)物進行研究,結果表明Mef2能夠特異性內(nèi)切α-1,3連接的糖苷鍵并釋放出具有新分支連接的巖藻低聚糖,成為第一個來自糖海帶能夠降解極其復雜巖藻多糖的水解酶。然而,來源于Flavobacterium algicola的巖藻多糖酶OUC-FaFcn1僅具備內(nèi)切α-1,4糖苷鍵的特異性,通過隨機切割方式降解巖藻多糖中的α-1,4糖苷鍵,生成以雙糖為主的產(chǎn)物[41]。同樣地,來自F.haliotis的巖藻多糖酶Fhf2Δ484也有α-1,4位點的切割活性,能夠釋放出含有大量硫酸化2,4-巖藻糖殘基的低聚糖,但其底物特異性較低,對來源于3 種海藻(墨角藻、莫氏馬尾藻、匍枝馬尾藻)的巖藻多糖具有不同程度的水解活性[14]。目前關于外切型巖藻多糖酶的研究比較少,例如Furukawa等[46]從Vibriosp.N-5中分離得到外切型的巖藻多糖酶,其水解產(chǎn)物主要為硫酸化的巖藻糖和巖藻二糖。
巖藻糖苷酶具有較為廣泛的底物特異性,絕大多數(shù)巖藻糖苷酶為外切型,能夠水解以α-1,2、α-1,3、α-1,4或α-1,6連接的巖藻糖苷鍵并生成多樣的巖藻糖或巖藻聚糖等產(chǎn)物,其作用方式與巖藻糖苷酶的來源及GH家族分類密切相關。例如來源于W.fucanilytica的GH29家族巖藻糖苷酶Alf1_Wf能夠水解α-1,4巖藻糖基連接的多糖,與合成底物4-硝基苯基-α-L-巖藻吡喃糖苷生成游離的巖藻糖和巖藻低聚糖[47],而來自Paenibacillusthiaminolyticus的GH151家族巖藻糖苷酶可以水解芳香基巖藻糖和以α-1,2、α-1,3、α-1,4、α-1,6糖苷鍵與乙酰葡糖胺相連接的雙糖[48]。
巖藻多糖硫酸酯酶也具有內(nèi)切或外切酶活性。Silchenko等[37]的研究發(fā)現(xiàn),來源于菌株W.fucanilyticaCZ1127T的硫酸酯酶SWF5為內(nèi)切型酶,可以去除由α-1,3和α-1,4糖苷鍵交替相連的L-巖藻多糖中的4O-硫酸化作用,但不能水解由α-1,3糖苷鍵連接成骨架的巖藻多糖[49]。此外,不同的硫酸酯酶對底物中硫酸基的存在位置和分支存在與否也具有特異性。硫酸酯酶SWF1和SWF4均為外切型酶,硫酸酯酶SWF1不能催化低聚糖4F2,3S(6S)、4F2,3,4S(7S)和6F2,3S(6S)脫硫,但能催化2O-硫酸化四糖4F2S(4S)脫硫,與此相反,硫酸酯酶SWF4可以催化低聚糖4F2,3S(6S)、4F2,3,4S(7S)和6F2,3S(6S)脫硫,但不能催化四糖4F2S(4S)脫硫[37]。
4 巖藻多糖降解酶的結構和催化機制
依據(jù)酶的分子結構和催化特征,不同類型的巖藻多糖降解酶被劃分到GH29、GH95、GH107、GH139、GH141、GH151和GH168等多個GH家族中。除了GH139和GH168家族的巖藻多糖降解酶結構仍未被解析之外,其他家族巖藻多糖降解酶的三維結構已被報道。
GH29家族巖藻多糖降解酶為α-L-巖藻糖苷酶。根據(jù)序列同源性和底物特異性,該家族又被分為GH29-A和GH29-B兩個亞家族:GH29-B僅作用于α-1,3和α-1,4連接的巖藻糖苷鍵,而GH29-A的底物特異性更為廣泛[50-51]。此外,GH29-A對底物4-硝基苯基-α-L-巖藻吡喃糖苷或2-氯-4-硝基苯基-α-L-巖藻吡喃糖苷表現(xiàn)出一定的降解活性,但GH29-B不具有此特性[50]。Summers等[52]對GH29家族的巖藻糖苷酶GH29_0940進行結構解析,發(fā)現(xiàn)該酶由N端的一個催化結構域和C端的兩個碳水化合物結構域(carbohydrate-binding domain,CBM)組成,其整體結構呈現(xiàn)出一個N端TIM桶狀結構域、中心β-三明治結構域和C端β-三明治結構域。TIM桶狀催化結構域為(β/α)8/7的排列,缺失的一個α-螺旋被一個長而曲折的loop所取代,其中Asp162和Glu220分別作為催化親核試劑和質(zhì)子供體[52]。
與GH29家族的酶不同,GH95家族的巖藻糖苷酶具有更加嚴格的底物特異性和更為簡單的結構。例如巖藻糖苷酶AfcA是僅具有α-1,2位點切割活性的GH95家族水解酶,該酶的結構包括4 個區(qū)域:一個N端的β區(qū)域、一個螺旋連接區(qū)域、一個(α/α)6螺旋桶狀區(qū)域和一個C端的β區(qū)域。在其催化結構域中,Glu566和Asn423分別作為催化酸和催化親核試劑[53]。
GH107家族為內(nèi)切α-1,4巖藻多糖水解酶。來源于Flavobacteriaceae的FcnA是GH107家族中第一個被發(fā)現(xiàn)的巖藻多糖酶,該酶由1 個N端結構域(400 個氨基酸組成)、3 個重復的類免疫球蛋白折疊結構域和1 個C端結構域(80 個氨基酸組成)組成[54]。Vickers等[55]對來源于Psychromonassp.SW5A的GH107家族巖藻糖苷酶P5AFcnA進行解析,發(fā)現(xiàn)P5AFcnA的N端存在一個包含(β/α)8的桶狀結構域,但其中的1 個α-螺旋和1 個β-折疊并未形成規(guī)則的二級結構,而是分別形成無規(guī)卷曲。結合X射線晶體學和核磁共振的進一步分析表明,GH107家族酶與GH29家族的巖藻糖苷酶具有相同的結構和催化機制,但GH107家族巖藻糖苷酶的催化殘基高度不保守,可能反映出GH107家族的酶對不同結構的巖藻多糖降解具有特殊性[55]。
來自BacteroidesthetaiotaomicronVPI-5482的α-2-O-甲基-L-巖藻糖苷酶是目前唯一被發(fā)現(xiàn)的GH139家族酶,但其分子結構和催化機制仍未被解析[56]。在GH141家族中,唯一被解析結構的是內(nèi)切巖藻糖苷酶BT_1002,該酶也來源于BacteroidesthetaiotaomicronVPI-5482,其催化結構域折疊成1 個右旋平行β-螺旋,催化中心表面形成1 個延伸的催化口袋,或許有助于容納木糖和巖藻糖組成的雙糖,并且活性口袋中的D523和D564對該酶的催化起到重要作用[56]。對GH151家族酶的研究也比較少,來源于Paenibacillusthiaminolyticus的α-L-巖藻糖苷酶同工酶2也是唯一被解析結構的巖藻多糖降解酶。該酶包括3 個結構域:1 個由(β/α)8的TIM桶狀結構構成的N端結構域、2 個較短的α-螺旋和3 個310螺旋,其中活性結構域位于TIM桶狀結構的中心;中心結構域的結構類似于Rossmann折疊,由4 個α-螺旋和11 個β-折疊組成β桶狀結構;C端結構域是β-三明治折疊類型,由1 個長loop和1 個包含2 個β-折疊的C端loop組成[48]。GH151家族的巖藻糖苷酶已被證實對芳基巖藻糖、由α-1,2鍵與半乳糖相連的巖藻糖、α-1,2(或α-1,3、α-1,4、α-1,6)鍵與乙酰葡糖胺相連的巖藻糖有活性,但對巖藻三糖或具有L-巖藻糖-α-1,2-D-Galp表位的六糖無活性[29]。此外,碳水化合物活性酶數(shù)據(jù)庫(http://www.cazy.org/)中還注釋了一種新型的GH168家族巖藻多糖降解酶,該家族酶首次報道于2020年,是一種來源于細菌W.fucanilytica的內(nèi)切型α-1,3巖藻糖苷酶FunA[57]。超高性能排阻層析-質(zhì)譜和核磁共振分析表明,F(xiàn)unA能夠特異性切割2-O-硫酸化和非硫酸化巖藻糖殘基相連的α-1,3糖苷鏈,定向突變結果表明D206和E264是發(fā)揮FunA功能的關鍵位點[57]。
部分巖藻多糖降解酶晶體結構信息匯總?cè)绫?所示。

表2 部分巖藻多糖降解酶的晶體結構信息Table 2 Crystal structural information of some fucoidan-degrading enzymes
5 巖藻多糖降解酶的應用
5.1 酶促合成具有特殊活性的巖藻低聚糖
巖藻多糖及巖藻低聚糖逐漸成為生物活性分子的重要來源,例如第一個阿爾茨海默病藥物和兩種能夠抑制SARS-CoV-2的抗病毒候選藥物原料均含有巖藻多糖[62]。由于巖藻多糖降解酶對巖藻多糖底物的水解功能,使其具備了酶促合成高附加值巖藻低聚糖的巨大潛力。生物來源的酶促反應具有條件溫和、不污染環(huán)境和反應效率高等優(yōu)點,水解法生產(chǎn)的低分子質(zhì)量巖藻聚糖不僅保留了天然多糖的生物活性,而且作為功能性藥品或食品成分更容易被人體消化吸收,因此酶法制備巖藻低聚糖具有非常好的應用前景。近年來,研究者對巖藻多糖降解酶及其在制備巖藻低聚糖中的應用進行了較多研究,發(fā)現(xiàn)利用巖藻多糖降解酶能夠?qū)崿F(xiàn)巖藻低聚糖的制備,并且產(chǎn)生了一些具有抗癌、抗血栓或功能性益生元等特殊生理活性的巖藻低聚糖。在功能性藥物研發(fā)方面,Silchenko等[63]利用重組表達的巖藻多糖酶FFA1制備了巖藻低聚糖,并且證明該巖藻低聚糖具有一定的抗癌活性。此外,Silchenko等[64]利用來源于海洋細菌F.algae的重組巖藻多糖酶對天然巖藻多糖進行改性,獲得分子質(zhì)量為50.8 kDa的聚合物,并證實聚合物對結腸癌細胞具有抑制作用。Xu Zhigang等[65]使用巖藻多糖酶對海帶多糖進行水解,制備了分子質(zhì)量為25.8 kDa的海帶低聚糖,動物實驗表明海帶低聚糖能夠顯著抑制血栓的形成。在功能性食品研發(fā)方面,使用巖藻糖苷酶能夠生產(chǎn)巖藻糖基化的人乳寡糖,這種人乳寡糖具有益生元、免疫調(diào)節(jié)等生理活性,在嬰兒大腦發(fā)育過程產(chǎn)生諸多有益影響[66]。因此,巖藻多糖降解酶在制備具有特殊生理活性巖藻低聚糖的過程中發(fā)揮了關鍵作用,這些低聚糖具有極高的應用價值,為未來功能性藥物或食品的研發(fā)提供重要基礎。
5.2 疾病診斷與治療
人的血清N-聚糖能夠作為一些癌癥、幽門螺桿菌感染等疾病的免疫反應指標,而具有不同底物特異性的巖藻糖苷酶已被用作驗證和監(jiān)測這些聚糖生物標志物[29,67-68]。此外,巖藻多糖降解酶也可應用到某些疾病的治療中,例如Manivasagan等[69]通過鏈霉菌生產(chǎn)新型巖藻多糖酶以綠色合成金納米顆粒,生物合成的金納米顆粒對HeLa細胞毒性呈劑量依賴性,這表明新型的巖藻糖苷酶能夠應用于抗癌治療。也有研究表明用巖藻多糖內(nèi)切酶Fhf1水解巖藻多糖,其中獲得的低分子質(zhì)量巖藻聚糖會影響骨再生過程[70]。
5.3 推斷巖藻多糖的分子結構
巖藻多糖是一種分子結構極為復雜的硫酸生物多糖。根據(jù)作用方式和底物特異性的不同,巖藻多糖降解酶對巖藻多糖的作用位點和水解產(chǎn)物存在一定的差異性,不同結構的巖藻低聚糖產(chǎn)物的生物活性也存在多樣化。某些特定的巖藻多糖降解酶能夠?qū)R恍缘厮馓厥忸愋偷奶擒真I,結合不同光譜分析、氣相色譜-質(zhì)譜分析及核磁共振等研究方法,能夠有效地推斷復雜巖藻多糖的分子結構。例如一些光譜分析方法可以用于闡明巖藻多糖硫酸基和糖苷鍵的位置或分子質(zhì)量大小,而編碼酶基因的宏基因組學分析可以揭示酶的作用位點和機制,該技術與光譜技術的結合能夠針對性地研究巖藻多糖潛在的水解位點[9]。探究巖藻多糖酶的作用位點以及了解巖藻多糖的結構能夠更好地改進生產(chǎn)巖藻低聚糖的技術,更加有效地制備具有特殊生物學活性和功效的低聚巖藻多糖。
6 結語
巖藻多糖為海洋特異性多糖,每種巖藻多糖的分子組成、結構及硫酸化程度均存在一定的差異。組成巖藻多糖的單糖包含木糖、甘露糖、半乳糖和其他一些稀有的單糖,并且單糖取代基的糖苷鍵類型也不盡相同。除此之外巖藻多糖中還存在大量的非碳水化合物取代基,因此巖藻多糖結構具有十分復雜的多樣性,對巖藻多糖的結構研究也相當困難。盡管如此,不同分子質(zhì)量的巖藻多糖因其具有抗癌、免疫刺激、抗氧化、抗感染等生物學活性和毒性低等特點而備受關注。
巖藻多糖降解酶是進一步挖掘巖藻聚糖資源的重要研究工具,同時巖藻多糖的多樣性也為發(fā)現(xiàn)具有不同特異性的巖藻多糖降解酶提供了機會。目前,國內(nèi)外關于巖藻多糖酶制劑制備的研究還較為缺乏,同時由于不同巖藻多糖降解酶的酶學性質(zhì)、水解方式、家族分布和分子結構等差異較大,對該酶的認識也有待不斷地增加和完善。不同特異性的巖藻多糖降解酶能夠?qū)r藻多糖水解為特定分子質(zhì)量的巖藻聚糖,并且有助于研究不同巖藻聚糖的特殊生物學活性,同時也為進一步闡明巖藻多糖的結構提供新的思路。因此,充分分析不同性質(zhì)、結構和作用方式的巖藻多糖降解酶不僅對酶的功能多樣性研究提供重要的理論支持,而且對于通過基因編輯和蛋白質(zhì)工程實現(xiàn)酶的定向改造,研發(fā)催化性能更好的酶制劑,獲得特殊功效的低分子質(zhì)量巖藻多糖具有重要的應用意義。
