甘薯PEBP基因家族鑒定以及影響甘薯塊根發(fā)育候選PEBP基因的鑒定
2024-06-05 08:36:45黃哲瑞辛曙麗趙添劉永華朱國(guó)鵬
熱帶作物學(xué)報(bào) 2024年3期
黃哲瑞 辛曙麗 趙添 劉永華 朱國(guó)鵬

關(guān)鍵詞:甘薯;PEBP 基因家族;生物信息學(xué);組織表達(dá)特異性;塊根發(fā)育
磷脂酰乙醇胺結(jié)合蛋白(phosphatidylethanolamine-binding protein, PEBP)具有1 個(gè)非常保守的PEBP 結(jié)構(gòu)域,該結(jié)構(gòu)域因能與磷脂酰乙醇胺結(jié)合而得名[1]。PEBP 是一種非常保守的蛋白,在生物界中廣泛存在,無(wú)論是植物、動(dòng)物還是古細(xì)菌中均有存在[2]。植物中的PEBP 最早在擬南芥中發(fā)現(xiàn),可分為3 個(gè)亞家族:FT (FLOWERINGLOCUS T)-like、TFL1 (TERMINAL FLOWER 1)-like、MFT (MOTHER OF FT AND TFL1)- like[3]。隨著研究的深入,在小麥、棉花等作物中發(fā)現(xiàn)PEBP 家族中還存在另一個(gè)亞家族,將其命名為PEBP-like[4-5]。
PEBP 家族中成員較多,功能多樣[3]。在擬南芥中存在6 個(gè)PEBP 家族成員,其中2 個(gè)屬于FT-like(AtFT、AtTSF)、3 個(gè)屬于TFL1-like(AtTFL1、AtBFT、AtATC),1 個(gè)屬于MFT-like(AtMFT)。擬南芥AtFT 基因與AtTFL1 基因在開(kāi)花誘導(dǎo)方面顯示出顯著的拮抗功能[6]。在長(zhǎng)日照條件下,擬南芥AtFT 的上游基因CONSTANS(CO)表達(dá)上升并與光敏色素作用因子PIF4 相互作用于AtFT的啟動(dòng)子區(qū)域,從而激活了葉片中FT 的表達(dá)[7],F(xiàn)T 蛋白在葉片中產(chǎn)生后,通過(guò)維管束運(yùn)輸?shù)巾敹朔稚M織的細(xì)胞質(zhì)中[8],與bZIP 轉(zhuǎn)錄因子FD 蛋白結(jié)合形成FT/FD 蛋白復(fù)合物,從而誘導(dǎo)下游成花相關(guān)基因的表達(dá),促進(jìn)擬南芥開(kāi)花[9]。在短日照條件下,AtTFL1 基因表達(dá)上升,而FD 既能和FT 蛋白結(jié)合,也能和TFL1 蛋白結(jié)合,因此TFL1蛋白通過(guò)與FT 蛋白競(jìng)爭(zhēng)和FD 的結(jié)合,從而抑制擬南芥開(kāi)花[10]。擬南芥FT-like 的另一成員TSF會(huì)協(xié)助FT 對(duì)FD的競(jìng)爭(zhēng)。而TFL1-like 成員AtBFT、AtATC 也會(huì)協(xié)助TFL1 對(duì)FD 的競(jìng)爭(zhēng)。因此擬南芥開(kāi)花不僅取決于FT 和TFL1,還與FT-like、TFL1-like 其他成員的表達(dá)水平有關(guān)[11]。過(guò)表達(dá)研究表明,AtMFT 有促進(jìn)擬南芥開(kāi)花的作用,AtMFT 基因還可以通過(guò)調(diào)控ABA 和GA信號(hào)來(lái)調(diào)節(jié)種子的萌發(fā)[12-14]。與擬南芥FT-like 類(lèi)似,水稻FT-like的RICE FLOWERING LOCUS T1(RFT1)基因在長(zhǎng)日照時(shí)具有誘導(dǎo)開(kāi)花的作用[15]。
除了調(diào)控植物開(kāi)花和種子萌發(fā)外,PEBP 還可調(diào)控植物地下部?jī)?chǔ)藏器官如鱗莖、塊莖等的生長(zhǎng)發(fā)育。如洋蔥(Allium cepa L)FT-like 中的AcFT1和AcFT4 對(duì)洋蔥磷莖的發(fā)育有拮抗作用。在長(zhǎng)日照條件下,AcFT1 基因表達(dá)量升高,能夠促進(jìn)洋蔥產(chǎn)生鱗莖,而在短日照條件下,AcFT4 表達(dá)量增加,抑制AcFT1 基因的轉(zhuǎn)錄,使洋蔥提早開(kāi)花,同時(shí)鱗莖的形成受到抑制[16]。在馬鈴薯中,過(guò)表達(dá)FT 同源基因StSP6A 會(huì)抑制蔗糖轉(zhuǎn)運(yùn)蛋白StSWEET11 的活性,使蔗糖的卸載途徑從質(zhì)外體途徑轉(zhuǎn)變?yōu)樘寝D(zhuǎn)運(yùn)效率更高的共質(zhì)體途徑,從而提高蔗糖向塊莖的轉(zhuǎn)運(yùn)和馬鈴薯塊莖的形成[17]。而另一個(gè)FT 同源基因StSP5G 可以通過(guò)抑制StSP6A 的表達(dá)阻礙馬鈴薯塊莖的形成[18]。馬鈴薯StCEN1 屬于TFL1-like,與脫落酸和細(xì)胞分裂素的信號(hào)傳導(dǎo)有關(guān)[19],StCEN1 的過(guò)表達(dá)也可以抑制馬鈴薯塊莖的形成[20]。水稻FT 同源基因Hd3a的產(chǎn)物在短日照條件下會(huì)與細(xì)胞質(zhì)的14-3-3 蛋白結(jié)合形成二級(jí)復(fù)合體,然后該復(fù)合體進(jìn)入細(xì)胞核中與bZIP 轉(zhuǎn)錄因子FD 蛋白結(jié)合形成三級(jí)復(fù)合體FAC,誘導(dǎo)水稻提早開(kāi)花[21]。此外,水稻Hd3a基因在馬鈴薯中過(guò)表達(dá)能夠促進(jìn)馬鈴薯塊莖的形成[18]。還有研究表明PEBP 基因也在番茄[22]、大麥[23-24]、毛竹[25]、木薯[26]、蘋(píng)果[27-28]、郁金香[29]等多種作物的生長(zhǎng)發(fā)育中發(fā)揮著重要的調(diào)控作用。
甘薯(Ipomoea batatas)別名甜薯、地瓜、番薯,為旋花科草本植物。甘薯起源于中南美洲,在16 世紀(jì)末才傳入中國(guó)[30]。由于甘薯環(huán)境適應(yīng)能力強(qiáng)、產(chǎn)量較高且易于栽培管理,因此很快就在中國(guó)廣泛種植[31]。甘薯塊根、莖、葉均可食用,富含淀粉、可溶性糖、維生素C、維生素E、胡蘿卜素、花青素等人體所需物質(zhì)[32]。此外,甘薯還具有重要的經(jīng)濟(jì)價(jià)值,可作為飼料、淀粉加工原料和工業(yè)原材料[33]。甘薯已經(jīng)成為全球第七大農(nóng)作物,我國(guó)甘薯栽培面積和產(chǎn)量分別占全球的42%和68%,穩(wěn)居世界第一(FAO,2020)。目前我國(guó)甘薯單產(chǎn)已處于平臺(tái)期,近20 年來(lái)(1999—2020 年)一直徘徊在22 t/hm2 左右,沒(méi)有進(jìn)一步提升(FAO, 2020)。雖然和大多數(shù)發(fā)展中國(guó)家相比我國(guó)甘薯單產(chǎn)較高,但和發(fā)達(dá)國(guó)家相比,我國(guó)甘薯單產(chǎn)仍有一定差距,例如美國(guó)和澳大利亞平均單產(chǎn)分別高達(dá)25.4 t/hm2 和39 t/hm2(FAO,2020)。如何進(jìn)一步提升甘薯產(chǎn)量是我國(guó)甘薯產(chǎn)業(yè)健康、可持續(xù)發(fā)展所亟需解決的重要問(wèn)題之一。
與馬鈴薯塊莖和洋蔥鱗莖等類(lèi)似,甘薯的主要食用器官塊根也屬于地下部?jī)?chǔ)藏器官,因此對(duì)其PEBP 家族成員進(jìn)行研究將有助于進(jìn)一步提高甘薯產(chǎn)量。雖然已有研究報(bào)道甘薯PEBP 基因家族,但該研究?jī)H從甘薯基因組中鑒定出13 個(gè)PEBP 家族成員,不包含PEBP-like 成員[34]。截至目前,尚未有報(bào)道系統(tǒng)鑒定和闡明與甘薯塊根發(fā)育相關(guān)的候選PEBP 家族成員。鑒于此,本研究首先利用生物信息學(xué)方法對(duì)甘薯基因組中PEBP家族成員的數(shù)量和種類(lèi)進(jìn)行系統(tǒng)鑒定,然后通過(guò)生物信息學(xué)分析PEBP 基因表達(dá)的組織特異性(幼葉、成熟葉、莖、纖維根、柴根、薯皮、塊根、花),初步鑒定出可能調(diào)控甘薯塊根發(fā)育的候選PEBP 家族成員,最后通過(guò)對(duì)候選PEBP 基因在不同發(fā)育時(shí)期塊根中表達(dá)水平的動(dòng)態(tài)變化及其與SWEET 基因表達(dá)水平之間的相關(guān)性分析,進(jìn)一步確認(rèn)其在調(diào)控甘薯塊根發(fā)育中的重要作用。本研究將為后續(xù)深入研究甘薯PEBP 基因家族的功能奠定基礎(chǔ),同時(shí)為進(jìn)一步提高我國(guó)甘薯產(chǎn)量提供理論支撐。
1 材料與方法
1.1 材料
選取甘薯品種高系14 為實(shí)驗(yàn)材料,于海南大學(xué)(20°2?39.73?N,110°18?26.93?E)進(jìn)行露地栽培,以史丹利復(fù)合肥(N∶P∶K=15∶15∶15)450 kg/hm2 和羊糞有機(jī)肥12 000 kg/hm2 作為基肥,在種植前先整地起壟,按照40 cm 間隔起壟,壟寬80 cm,每壟只種1 行甘薯,株距為25 cm。種植時(shí)間為2021 年9 月20 號(hào)至2021 年12 月3號(hào),開(kāi)花后(74 d)取樣。為了研究不同組織的基因表達(dá),選擇高系14 的幼葉(莖尖周?chē)奈凑归_(kāi)葉)、成熟葉(從莖尖向下數(shù)的第5 片完全展開(kāi)葉片)、莖、花(不包括花柄和花萼)、纖維根(白色細(xì)長(zhǎng)呈纖維狀的根)、柴根(直徑0.2~2.0 cm 粗細(xì)均勻的紅色根)、塊根(直徑約4 cm 的膨大根)的薯皮和薯肉8 個(gè)部位(圖1),每個(gè)部位取0.2 g,3 個(gè)生物學(xué)重復(fù)。
2022 年1—5 月在海南大學(xué)海甸校區(qū)溫室大棚盆栽種植高系14。花盆規(guī)格為:外徑29.6 cm,內(nèi)徑25.4 cm,高19.7 cm,底部直徑17.8 cm。基肥用羊糞有機(jī)肥和史丹利復(fù)合肥(N∶P∶K=15∶15∶15),田園土和羊糞有機(jī)肥按3∶1 的體積比加上每盆0.8 g 的史丹利復(fù)合肥(N∶P∶K=15∶15∶15)混合均勻,裝入花盆。剪取25 cm 長(zhǎng)帶莖尖的甘薯莖段,以直插法進(jìn)行扦插,深度為10 cm。在甘薯扦插后的30、60、90、120 d 取樣,取0.2 g塊根薯肉提取總RNA,每個(gè)取樣時(shí)期均含4 個(gè)生物學(xué)重復(fù)。取樣后液氮處理,置于–80 ℃超低溫冰箱保存?zhèn)溆谩?/p>
1.2 方法
1.2.1 甘薯PEBP 基因家族成員鑒定 從甘薯基因組數(shù)據(jù)庫(kù)(https://ipomoea-genome.org/)下載甘薯的全基因組文件和全基因組注釋文件。從NCBI 數(shù)據(jù)庫(kù)(https://www.ncbi.nlm.nih.gov/)下載擬南芥PEBP 家族的蛋白序列[26]。從Phytozomev13 數(shù)據(jù)庫(kù)(https://phytozome-next.jgi.doe.gov/)下載木薯PEBP 家族的蛋白序列[26]。從馬鈴薯的Spud DB 數(shù)據(jù)庫(kù)(http://solanaceae.plantbiology.msu.edu)下載馬鈴薯PEBP 家族的蛋白序列[35]。
為了鑒定甘薯PEBP 候選基因,使用2 種方法搜索甘薯的蛋白質(zhì)序列。第一種方法:準(zhǔn)備擬南芥(6 個(gè))、木薯(10 個(gè))、馬鈴薯(15 個(gè))的PEBP基因家族蛋白序列,通過(guò)TBtools[36](https://github.com/CJ-Chen/TBtools)在線軟件將甘薯全蛋白序列進(jìn)行本地Blastp 比對(duì),參數(shù)閾值設(shè)置為E-value≤1e–5,其他參數(shù)為默認(rèn)值。第二種方法:通過(guò)查找PEBP 家族蛋白的隱馬爾可夫模型(HMM)的ID(PF01161)、下載甘薯全蛋白序列和Pfam-A.hmm 文件(ftp://ftp.ebi.ac.uk/pub/databasesfam/current_release/Pfam-A.hmm.gz)。將ID(PF01161)、甘薯全蛋白序列、Pfam-A.hmm 文件用TBtools 軟件檢索出可能的甘薯PEBP 基因。
將2 種方法得到的候選基因,提交給Pfam 數(shù)據(jù)庫(kù)[37](http://pfam.xfam.org/)、NCBI 網(wǎng)站[38]( https://www.ncbi.nlm.nih.gov/Structu-re/cdd/cdd.html ) 和SMART 網(wǎng)站[39] ( http://smart.embl-Heidelberg.de/smart/set_mode.cgi?NORMAL=1)進(jìn)一步確認(rèn)是否含有保守的PEBP 結(jié)構(gòu)域。
1.2.2 甘薯PEBP 家族成員染色體定位 使用TBtools 軟件分析甘薯PEBP 家族成員基因的注釋信息。
1.2.3 甘薯PEBP 家族蛋白系統(tǒng)發(fā)育分析 為了對(duì)甘薯的PEBP家族基因進(jìn)行分類(lèi)系統(tǒng)發(fā)育分析,使用MEGA-X(https://www.megasoftware.net/)在線軟件的鄰接法(Neighbor-joining method)將擬南芥、木薯、馬鈴薯、甘薯的PEBP 蛋白的全長(zhǎng)氨基酸序列構(gòu)建系統(tǒng)發(fā)育樹(shù),其中參數(shù)設(shè)定的自展值為1000,其他參數(shù)默認(rèn)。使用Evolview( https://www.evolgenius.info/evolview/)在線軟件對(duì)系統(tǒng)發(fā)育樹(shù)進(jìn)行美化。此外,按上述方法用MEGA-X 軟件將甘薯PEBP 基因與其他物種已知功能的PEBP 基因(馬鈴薯的StSP6A、StSP5G、StCEN1;洋蔥的AcFT1 和AcFT4;水稻的Hd3a)構(gòu)建系統(tǒng)發(fā)育樹(shù),以此推斷相關(guān)甘薯PEBP 基因的功能。
1.2.4 甘薯PEBP 家族成員理化性質(zhì)分析 使用ExPASy( http://web.expasy.org/protparam/)在線軟件預(yù)測(cè)PEBP 蛋白的理化性質(zhì)(等電點(diǎn)、分子量、氨基酸數(shù)目)。使用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)在線軟件進(jìn)行亞細(xì)胞定位預(yù)測(cè)。
1.2.5 甘薯PEBP 基因家族成員表達(dá)分析 使用Premier 5.0 軟件設(shè)計(jì)實(shí)時(shí)定量PCR 所需引物,并在NCBI 網(wǎng)站檢驗(yàn)引物特異性。其中IbFT1、IbFT3與IbFT2 的CDS 序列相似度高且前2 個(gè)基因的CDS 序列比IbFT2 短,IbTFL1 與IbTFL2 相比也是如此,因此IbFT1、IbFT3、IbTFL1 無(wú)法設(shè)計(jì)特異性引物,后續(xù)不再對(duì)IbFT1、IbFT3、IbTFL1進(jìn)行基因表達(dá)研究。具體引物信息如表1 所示,其中,5 個(gè)SWEET 基因(IbSWEET4、IbSWEET7、IbSWEET11、IbSWEET16 和IbSWEET19)的引物信息參照張文杰[40]的研究。
利用CWBIO 公司的RNA 提取試劑盒[OmniPlantRNA Kit(Dnase I)]提取總RNA,并通過(guò)瓊脂糖凝膠電泳檢查其完整性;逆轉(zhuǎn)錄使用諾唯贊HiScript III 1st Strand cDNA Synthesis Kit 逆轉(zhuǎn)錄試劑盒;使用諾唯贊ChamQ Universal SYBRqPCR Master Mix 試劑盒,以甘薯的β-Actin 基因?yàn)閮?nèi)參,用德國(guó)耶拿qTOWER3G 定量PCR 儀進(jìn)行基因表達(dá)水平測(cè)定。具體反應(yīng)條件如下:95 ℃預(yù)變性5 min;95 ℃變性5 s,58 ℃退火30 s,72 ℃延伸30 s,40 個(gè)循環(huán)。采用2–ΔΔCT 法計(jì)算基因的相對(duì)表達(dá)量。
1.3 數(shù)據(jù)處理
利用Excel 2020 軟件和Graphpad prism 8.0 軟件處理甘薯PEBP 基因相對(duì)表達(dá)數(shù)據(jù),使用SPSS22.0 軟件對(duì)數(shù)據(jù)進(jìn)行單因素方差分析(ANOVA),多重比較采用Duncans 法分析。
2 結(jié)果與分析
2.1 甘薯PEBP 基因家族成員的鑒定
通過(guò)本地Blastp 比對(duì)和隱馬爾可夫模型(HMM)分析,從甘薯基因組中找到16 個(gè)PEBP候選基因,然后將其提交到Pfam 數(shù)據(jù)庫(kù)、NCBI網(wǎng)站和SMART 網(wǎng)站驗(yàn)證,發(fā)現(xiàn)編號(hào)為g46241.t1的蛋白無(wú)PEBP 結(jié)構(gòu)域,將其剔除。因此,從甘薯基因組中總共鑒定出15 個(gè)PEBP 家族成員。用MEGA-X 軟件將鑒定出的15 個(gè)甘薯PEBP 家族成員與已經(jīng)公布的擬南芥、木薯、馬鈴薯的PEBP家族成員用鄰接法構(gòu)建系統(tǒng)發(fā)育樹(shù)(圖2)。結(jié)果顯示,15 個(gè)甘薯PEBP 家族成員被分為4 個(gè)亞家族,其中包含5 個(gè)FT-like 成員、6 個(gè)TFL1-like成員、2 個(gè)MFT-like 成員和2 個(gè)PEBP-like 成員。根據(jù)每個(gè)亞家族成員在染色體上位置的先后順序進(jìn)行命名,分別命名為IbFT1~5、IbTFL1~6、IbMFT1~2、IbPEBP1~2(表2)。值得注意的是,現(xiàn)有研究?jī)H在甘薯基因組中鑒定出13 個(gè)PEBP 家族成員,包括5 個(gè)FT-like 成員、6 個(gè)TFL1-like成員、2 個(gè)MFT-like 成員,但缺少2 個(gè)PEBP-like成員[34]。
2.2 甘薯PEBP 基因家族成員的功能預(yù)測(cè)
為了鑒定出可能與甘薯塊根發(fā)育相關(guān)的PEBP家族成員,將其他農(nóng)作物中已知會(huì)促進(jìn)或抑制地下部?jī)?chǔ)藏器官膨大的PEBP 基因(包括馬鈴薯的StSP6A、StSP5G 和StCEN1 基因[17-20],洋蔥的AcFT1 和AcFT4 基因[16],以及水稻的Hd3a 基因[18])和甘薯的15 個(gè)PEBP 家族成員進(jìn)行聚類(lèi)分析(圖3),通過(guò)親緣關(guān)系的遠(yuǎn)近預(yù)測(cè)可能和塊根發(fā)育相關(guān)的甘薯PEBP 基因。結(jié)果發(fā)現(xiàn),在15 個(gè)甘薯PEBP 基因中,IbFT5 與洋蔥基因AcFT1 和水稻基因Hd3a 的親緣關(guān)系較近,而這2 個(gè)基因可分別促進(jìn)洋蔥鱗莖[16]和馬鈴薯塊莖的發(fā)育[18]。因此,推測(cè)IbFT5 可能和甘薯塊根發(fā)育密切相關(guān)。此外,與其他甘薯PEBP 基因相比,IbTFL3 和馬鈴薯中抑制塊莖膨大的TFL1-like 基因StCEN1(StPEBP5)[20, 35]的親緣關(guān)系較近,因此推測(cè)IbTFL3 可能會(huì)抑制甘薯塊根的發(fā)育。
2.3 甘薯PEBP 基因的組織特異性表達(dá)分析
為進(jìn)一步揭示甘薯PEBP 家族成員的功能,對(duì)甘薯PEBP 基因在不同組織(幼葉、成熟葉、莖、纖維根、柴根、薯皮、薯肉、花)中的表達(dá)特異性進(jìn)行qRT-PCR 分析(圖4)。結(jié)果表明,IbTFL5、IbMFT1 和IbPEBP2 在所有組織中均具有較高的表達(dá)水平,其可能在不同組織的發(fā)育中均發(fā)揮著重要作用,而與塊根的膨大無(wú)必然聯(lián)系。
IbFT2 只在幼葉和成熟葉中表達(dá),為葉片特異表達(dá)基因。IbMFT2 在花中的表達(dá)水平遠(yuǎn)高于其他組織,為花特異性表達(dá)基因。IbFT4 僅在花和成熟葉中有表達(dá),且在花中的表達(dá)水平顯著高于成熟葉,為花和葉片特異表達(dá)基因。IbTFL3、IbTFL4和IbTFL6 不僅在花中高表達(dá),也在塊根的薯皮和薯肉中高表達(dá)。根系特異表達(dá)的基因?yàn)镮bFT5、IbTFL2 和IbPEBP1,其中IbFT5 僅在非膨大根(纖維根和柴根)中高表達(dá),而IbTFL2 和IbPEBP1不僅在非膨大根中高表達(dá),也在膨大塊根的薯皮和薯肉中高表達(dá)。上述結(jié)果表明,甘薯PEBP 基因家族成員的表達(dá)具有明顯的組織特異性,推測(cè)甘薯PEBP 家族成員存在顯著的功能分化,其在不同組織中發(fā)揮著不同的功能。
由圖4 可知,在根系(膨大根和非膨大根)中特異表達(dá)基因?yàn)镮bFT5、IbTFL2 和IbPEBP1。
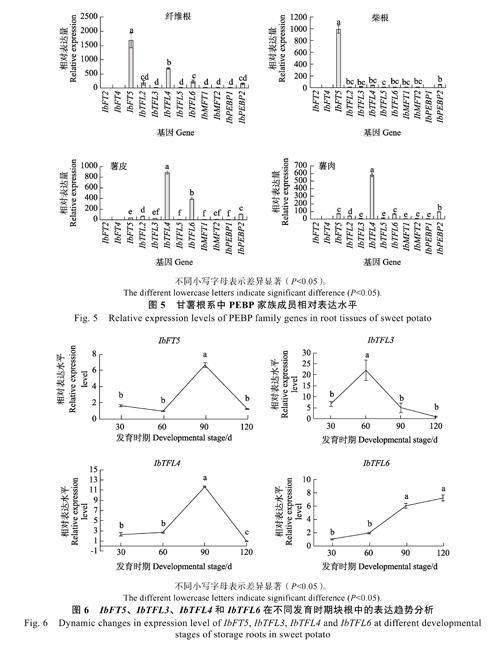
此外,IbTFL3、IbTFL4 和IbTFL6 也在根系中特別是薯皮和薯肉中高表達(dá)。推測(cè)上述6 個(gè)甘薯PEBP 基因可能與甘薯塊根發(fā)育密切相關(guān)。為縮小PEBP 基因成員的選擇范圍,本課題組進(jìn)一步對(duì)特定根系組織(纖維根、柴根、薯皮和薯肉)中不同PEBP 家族成員的相對(duì)表達(dá)水平進(jìn)行測(cè)定(圖5)。結(jié)果發(fā)現(xiàn),在纖維根中,IbFT5 和IbTFL4為高表達(dá)基因,且IbFT5 的表達(dá)水平顯著高于IbTFL4。在柴根中,IbFT5 為高表達(dá)基因,其余基因的表達(dá)水平極低。在膨大塊根薯皮中,IbTFL4、IbTFL6 為高表達(dá)基因,其余基因低表達(dá)或不表達(dá)。在膨大塊根薯肉中,IbTFL4 為高表達(dá),其余基因?yàn)榈捅磉_(dá)或不表達(dá)。
綜上所述,在根系中高表達(dá)的6 個(gè)PEBP 基因中,IbFT5、IbTFL4 和IbTFL6 不僅在根系中的表達(dá)水平高于在其他組織,而且也高于根系中其他PEBP 家族成員。因此推測(cè)這3 個(gè)PEBP 基因在塊根膨大中發(fā)揮著重要作用。
2.4 甘薯PEBP 基因在不同發(fā)育時(shí)期塊根中的表達(dá)及其與SWEET 基因表達(dá)的相關(guān)性
聚類(lèi)分析揭示IbFT5 可能會(huì)促進(jìn)甘薯塊根的發(fā)育,而IbTFL3 則可能會(huì)抑制甘薯塊根的發(fā)育(圖3);此外,組織表達(dá)特異性分析則表明IbFT5、IbTFL4 和IbTFL6 與塊根發(fā)育密切相關(guān)(圖4、圖5)。因此,本課題組進(jìn)一步對(duì)上述4 個(gè)基因(IbFT5、IbTFL3、IbTFL4 和IbTFL6)在不同發(fā)育時(shí)期塊根(定植后30、60、90、120 d)中的表達(dá)水平動(dòng)態(tài)變化進(jìn)行測(cè)定,觀察上述基因表達(dá)水平和塊根膨大之間的相關(guān)性。
結(jié)果表明,在塊根發(fā)育的前期(30~60 d),IbFT5、IbTFL4 的表達(dá)水平無(wú)顯著變化,而在塊根發(fā)育中期(60~90 d),這2 個(gè)基因的表達(dá)水平快速大幅增加,但在塊根發(fā)育后期(90~120 d),其表達(dá)水平均快速下降(圖6)。與IbFT5、IbTFL4不同,IbTFL6 的表達(dá)水平在整個(gè)塊根發(fā)育時(shí)期一直呈上升的趨勢(shì),在整個(gè)發(fā)育時(shí)期呈‘S形曲線,即在塊根發(fā)育前期(30~60 d)和后期(90~120 d),其表達(dá)水平上升速度較慢,而在發(fā)育中期則上升速度較快。
在上述4 個(gè)基因(IbFT5、IbTFL3、IbTFL4和IbTFL6)中,只有IbTFL3 基因根據(jù)聚類(lèi)分析(圖3)被認(rèn)為可能會(huì)抑制甘薯塊根的發(fā)育。IbTFL3 基因表達(dá)水平的動(dòng)態(tài)分析表明,其表達(dá)水平在塊根膨大前期(30~60 d)呈上升趨勢(shì),而在塊根發(fā)育的中后期(60~120d)其表達(dá)水平快速下降。總的來(lái)講,伴隨著IbFT5、IbTFL4 和IbTFL6表達(dá)水平的上升,IbTFL3 的表達(dá)水平則呈現(xiàn)下降趨勢(shì),表明這2 類(lèi)基因之間可能存在拮抗作用,前者可能促進(jìn)塊根發(fā)育,而后者可能抑制塊根發(fā)育。
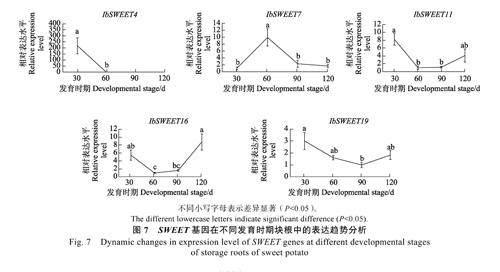
SWEET 蛋白為蔗糖轉(zhuǎn)運(yùn)蛋白,其主要功能是將蔗糖和己糖(葡萄糖和果糖)由細(xì)胞內(nèi)向外轉(zhuǎn)運(yùn),其表達(dá)量低往往代表糖分的轉(zhuǎn)運(yùn)途徑以高效的共質(zhì)體途徑為主,而低效的質(zhì)外體途徑則屬于次要途徑。例如,在馬鈴薯中,過(guò)表達(dá)PEBP基因StSP6A 會(huì)抑制蔗糖轉(zhuǎn)運(yùn)蛋白StSWEET11 的表達(dá),從而促進(jìn)蔗糖由葉片通過(guò)共質(zhì)體途徑向塊莖的高效轉(zhuǎn)運(yùn),最終促進(jìn)馬鈴薯塊莖的形成[17]。研究表明,在甘薯塊根中表達(dá)的SWEET 基因有5 個(gè),分別為IbSWEET4、IbSWEET7、IbSWEET11、IbSWEET16 和IbSWEET19[40]。本課題組對(duì)不同發(fā)育時(shí)期塊根中這5 個(gè)SWEET 基因的表達(dá)水平進(jìn)行測(cè)定。結(jié)果表明,隨著塊根膨大和塊根發(fā)育相關(guān)PEBP 基因(IbFT5、IbTFL4 和IbTFL6)表達(dá)水平的上升,IbSWEET4、IbSWEET11、IbSWEET16和IbSWEET19 的表達(dá)均呈現(xiàn)不斷下降的趨勢(shì),在定植后60 d(IbSWEET19 是在定植后90 d)達(dá)到最低值,其表達(dá)水平在塊根膨大后期(90~120 d)又呈現(xiàn)回升的趨勢(shì),而IbSWEET7 基因的表達(dá)在塊根膨大前期(30~60 d)呈現(xiàn)上升的趨勢(shì)(圖7)。根據(jù)上述結(jié)果,可推測(cè)甘薯PEBP 基因很可能也是通過(guò)抑制SWEET 基因的表達(dá)來(lái)促進(jìn)塊根發(fā)育。
3 討論
PEBP 家族在植物中廣泛存在,近年來(lái)研究人員已經(jīng)在多種植物中發(fā)現(xiàn)了PEBP 基因家族。PEBP 基因在植物生長(zhǎng)發(fā)育中發(fā)揮著重要作用,不僅調(diào)控?cái)M南芥和水稻等植物的開(kāi)花[6, 12, 15, 21],還參與調(diào)控地下部?jī)?chǔ)藏器官的形成如馬鈴薯塊莖[17-20]和洋蔥鱗莖等[16]。與馬鈴薯塊莖和洋蔥鱗莖等類(lèi)似,甘薯的主要食用器官塊根也屬于地下部?jī)?chǔ)藏器官,因此對(duì)其PEBP 家族成員進(jìn)行研究將有助于進(jìn)一步提高其塊根產(chǎn)量。雖然近期有研究報(bào)道了甘薯PEBP 家族的生物信息學(xué)分析結(jié)果,但該研究從甘薯基因組中僅鑒定出13 個(gè)PEBP 家族成員,且只包含F(xiàn)T-like、TFL1-like 和MFT-like,并不包含PEBP-like[34]。此外,目前尚未有報(bào)道系統(tǒng)研究并篩選可能和甘薯塊根發(fā)育相關(guān)的候選PEBP家族成員。
為了解決上述存在的問(wèn)題,本研究利用生物信息學(xué)方法從甘薯基因組中共鑒定出16 個(gè)PEBP候選基因,然后將其提交到Pfam 數(shù)據(jù)庫(kù)、NCBI網(wǎng)站和SMART 網(wǎng)站驗(yàn)證,發(fā)現(xiàn)除了編號(hào)g46241.t1的成員,其他15 個(gè)甘薯PEBP 成員和前人研究的擬南芥、木薯、馬鈴薯結(jié)果相同,均存在著保守的PEBP 結(jié)構(gòu)域。將g46241.t1 剔除,得到15 個(gè)甘薯PEBP 家族成員。在植物進(jìn)化過(guò)程中,許多植物都經(jīng)歷過(guò)1次或多次多倍體化從而導(dǎo)致基因家族成員擴(kuò)張,擬南芥和木薯為二倍體,甘薯為六倍體,且擬南芥PEBP 家族成員(6 個(gè))和木薯PEBP 家族成員(10 個(gè))與甘薯成員相比,甘薯PEBP 家族成員數(shù)量更多,說(shuō)明甘薯在進(jìn)化過(guò)程中隨著染色體倍數(shù)的增加,導(dǎo)致了甘薯PEBP 家族的擴(kuò)張。此外,系統(tǒng)發(fā)育樹(shù)結(jié)果顯示與擬南芥和木薯相比,甘薯除了有FT-like、TFL1-like 和MFT-like 外,還多了PEBP-like,而在他人研究的馬鈴薯、小麥、棉花等多個(gè)物種中也鑒定出PEBP-like[4-5, 35]。甘薯有5 個(gè)FT-like 成員、6 個(gè)TFL1-like 成員、2 個(gè)MFT-like 成員和2 個(gè)PEBP-like成員,比較發(fā)現(xiàn)不管是在擬南芥、木薯還是馬鈴薯中的TFL1-like 成員數(shù)量在亞家族中都是最多的,推測(cè)在進(jìn)化過(guò)程中TFL1-like 受環(huán)境選擇發(fā)生的基因復(fù)制和變異的頻率更高。和已有的研究相比[34],本研究從甘薯基因組中多鑒定出2 個(gè)PEBP基因,且這2 個(gè)基因均為PEBP-like 成員。
在植物中PEBP 家族基因能夠影響貯藏器官的形成。在馬鈴薯中,塊莖為馬鈴薯的貯藏器官,過(guò)表達(dá)StSP6A 能夠促進(jìn)塊莖的形成,而過(guò)表達(dá)StSP5G 和StCEN1 抑制了塊莖的形成[17-20]。此外過(guò)表達(dá)水稻的Hd3a 基因也能夠促進(jìn)馬鈴薯塊莖的形成[18]。在洋蔥中,鱗莖為洋蔥的貯藏器官,過(guò)表達(dá)AcFT1 能夠促進(jìn)鱗莖的形成,而過(guò)表達(dá)AcFT4 能夠抑制鱗莖的形成[16]。甘薯、馬鈴薯、洋蔥、水稻聚類(lèi)分析發(fā)現(xiàn),IbFT5 與洋蔥基因AcFT1和水稻基因Hd3a 的親緣關(guān)系較近,而這2 個(gè)基因可分別促進(jìn)洋蔥鱗莖[16]和馬鈴薯塊莖的發(fā)育[18]。推測(cè)IbFT5 可能也會(huì)促進(jìn)甘薯塊根的發(fā)育。此外,和其他甘薯PEBP 基因相比,IbTFL3 和馬鈴薯StPEBP5(StCEN1)[20, 35]的親緣關(guān)系較近,推測(cè)IbTFL3 可能會(huì)抑制甘薯塊根的發(fā)育。
植物PEBP 基因家族具有組織特異性。木薯中MeFT1 和MeFT2 在成熟葉中高表達(dá),推測(cè)成熟葉是產(chǎn)生花誘導(dǎo)信號(hào)的部位。MeMFT1 在幼葉和花芽的生長(zhǎng)組織中高表達(dá),而MeMFT2 在須根和莖中高表達(dá)[26]。本研究中,甘薯根系(膨大根和非膨大根)特異表達(dá)的基因?yàn)镮bFT5、IbTFL2和IbPEBP1。此外,IbTFL3、IbTFL4 和IbTFL6除了在花中高表達(dá),也在根系中特別是薯皮和薯肉中高表達(dá),但在其他組織中為低表達(dá)或不表達(dá)。推測(cè)上述6 個(gè)PEBP 基因和甘薯塊根發(fā)育密切相關(guān)。為進(jìn)一步研究發(fā)現(xiàn),上述6 個(gè)基因在甘薯根系中(纖維根、柴根和膨大塊根薯皮和薯肉)中高表達(dá)的PEBP 基因只有IbFT5、IbTFL4、IbTFL6等3 個(gè)基因,其余基因?yàn)榈捅磉_(dá)。因此,在根系中高表達(dá)的6 個(gè)PEBP基因中,只有IbFT5、IbTFL4和IbTFL6 不僅在根系中的表達(dá)水平高于在其他組織中的表達(dá)水平,而且和根系中表達(dá)的其他PEBP 家族成員相比,這3 個(gè)基因的表達(dá)水平也是最高的。推測(cè)這3 個(gè)PEBP 基因(IbFT5、IbTFL4和IbTFL6)在根系膨大中發(fā)揮著重要的作用。
綜合上述2 個(gè)方面的結(jié)果,推測(cè)IbFT5、IbTFL4 和IbTFL6 可能會(huì)促進(jìn)塊根的發(fā)育,而IbTFL3 則可能會(huì)抑制塊根的發(fā)育。已有的研究表明,甘薯塊根定植后30 d 開(kāi)始逐漸膨大,定植后60~90 d 進(jìn)入快速膨大期,隨后膨大速度逐漸減慢[41]。對(duì)上述4 個(gè)基因在不同發(fā)育時(shí)期塊根(定植后30、60、90、120 d)中的表達(dá)水平進(jìn)行的測(cè)定結(jié)果表明,在塊根發(fā)育的前期(30~60 d),IbFT5、IbTFL4 的表達(dá)水平無(wú)顯著變化,而在塊根快速膨大期(60~90 d),這2 個(gè)基因的表達(dá)水平快速大幅增加。與IbFT5、IbTFL4 不同,IbTFL6的表達(dá)水平在整個(gè)塊根發(fā)育時(shí)期一直呈現(xiàn)上升的趨勢(shì),特別在塊根快速膨大期(60~90 d)其上升速度較快。這與馬鈴薯中的StSP6A 表達(dá)相似。馬鈴薯的StSP6A 也在貯藏器官發(fā)育時(shí)期呈現(xiàn)上升趨勢(shì),當(dāng)?shù)叵沦橘肭o彎鉤角度為150°時(shí),StSP6A的表達(dá)量急劇增加,發(fā)育成小薯和大薯時(shí),表達(dá)量也還保持在較高水平[42]。推測(cè)IbFT5、IbTFL4和IbTFL6 這3 個(gè)基因可能促進(jìn)甘薯塊根的發(fā)育,根據(jù)聚類(lèi)分析推測(cè)IbTFL3 基因可能會(huì)抑制甘薯塊根的發(fā)育。IbTFL3 基因表達(dá)水平的動(dòng)態(tài)變化分析表明,其表達(dá)水平在塊根快速膨大期(60~90 d)開(kāi)始快速下降,表明其很可能對(duì)塊根膨大起抑制作用。此外,IbTFL3 在柴根中表達(dá)水平顯著高于其他根系(纖維根、塊根的薯肉和薯皮),而柴根是生長(zhǎng)停滯的塊根,這進(jìn)一步證明IbTFL3 很可能會(huì)抑制塊根的膨大。
SWEET 蛋白為蔗糖轉(zhuǎn)運(yùn)蛋白,其主要功能是將蔗糖和己糖(葡萄糖和果糖)由細(xì)胞內(nèi)向外轉(zhuǎn)運(yùn),從而促進(jìn)糖分經(jīng)由低效率的質(zhì)外體途徑轉(zhuǎn)運(yùn),而不是經(jīng)由高效率的共質(zhì)體途徑轉(zhuǎn)運(yùn)。前人研究表明,在甘薯塊根中高表達(dá)SWEET 基因有5 個(gè),分別為IbSWEET4、IbSWEET7、IbSWEET11 、IbSWEET16 和IbSWEET19[40]。不同發(fā)育時(shí)期的塊根中,隨著塊根膨大和PEBP 基因(IbFT5、IbTFL4和IbTFL6)表達(dá)水平的上升,4 個(gè)SWEET 基因( IbSWEET4 、IbSWEET11 、IbSWEET16 和IbSWEET19)的表達(dá)均呈現(xiàn)不斷下降的趨勢(shì),在定植后60 d(IbSWEET19 是在定植后90 d)達(dá)到最低值。推測(cè)甘薯PEBP 基因很可能是通過(guò)抑制SWEET 基因的表達(dá)來(lái)促進(jìn)塊根發(fā)育。在馬鈴薯的研究中,過(guò)表達(dá)PEBP 基因,StSP6A 會(huì)抑制蔗糖轉(zhuǎn)運(yùn)蛋白StSWEET11 的表達(dá),從而促進(jìn)蔗糖通過(guò)共質(zhì)體途徑向塊莖的高效轉(zhuǎn)運(yùn),最終促進(jìn)馬鈴薯塊莖的形成[18]。此外,甘薯塊根中SWEET 基因表達(dá)水平的下降也可以解釋為何甘薯塊根在定植后40 d 塊根開(kāi)始膨大時(shí)其蔗糖轉(zhuǎn)運(yùn)途徑由質(zhì)外體途徑轉(zhuǎn)變?yōu)楣操|(zhì)體途徑[41]。
本研究不僅確定了甘薯基因組中PEBP 基因家族成員的數(shù)量和種類(lèi),而且通過(guò)生物信息學(xué)分析和PEBP 基因表達(dá)的組織特異性分析初步鑒定出可能調(diào)控甘薯塊根發(fā)育的候選PEBP家族成員。此外,通過(guò)對(duì)相關(guān)PEBP 基因在不同發(fā)育時(shí)期塊根中表達(dá)水平的動(dòng)態(tài)變化及其與SWEET 基因表達(dá)水平之間的相關(guān)性分析,初步推斷PEBP 可能通過(guò)調(diào)控糖分轉(zhuǎn)運(yùn)來(lái)影響甘薯塊根發(fā)育。本研究將為后續(xù)深入研究甘薯PEBP 基因家族的功能提供理論依據(jù)。
