腰果CBF基因家族的鑒定及響應冷脅迫的表達模式
2024-06-05 08:36:45崔恒久曾教科李雯黃海杰彭世清郭冬蒲金基周永凱李輝亮
熱帶作物學報 2024年3期
崔恒久 曾教科 李雯 黃海杰 彭世清 郭冬 蒲金基 周永凱 李輝亮
關鍵詞:腰果;低溫響應;CBF 轉錄因子;基因家族;qRT-PCR
植物生長受多種環境因素的控制,其中溫度是重要的因素之一[1]。低溫脅迫會影響甚至破壞細胞的動態平衡,導致植物生長發育受到嚴重影響[2]。C-repeat binding factor(CBF)轉錄因子,也叫做dehydration responsive element binding factor1蛋白(DREB1 蛋白)[3-4],在響應低溫脅迫中起重要作用,冷應答途徑是植物感受低溫信號重要的調控途徑之一。ICE-CBF-COR 是被研究最透徹的一條通路。植物對低溫等非生物脅迫抗性的提高與脯氨酸、可溶性糖等物質的積累密不可分,而相關物質的積累需要下游基因的激活表達。下游基因的激活表達需要CBF 轉錄激活因子與下游COR 基因啟動子中的DRE/CRT (C-repeat/dehydration responsiveelement)順式作用元件特異性結合[5-8]。
CBF轉錄因子屬于Apetala2/Ethylene responsivefactor 基因家族(AP2/ERF 基因家族),AP2/ERF由AP2 亞家族、ERF 亞家族和RAV 亞家族組成。目前,CBF 基因家族已經在多種植物中得以研究,擬南芥(Arabidopsis thaliana)研究表明,擬南芥基因組中含有6 個CBF 基因[9]。此外,CBF/DREB1轉錄因子還存在于其他植物中,如番茄[10-13]、水稻[14-20]、大豆[21-25]等。CBF/DREB1 轉錄因子具有高度保守的AP2/ERF 結構域。AP2/ERF 結構域含約60 個氨基酸[26]。
腰果(Anacardium occidentalie Linn)被譽為世界四大著名堅果之一。是典型的熱帶果樹,原產地在巴西東北部地區,16 世紀被引入亞洲、非洲以及南美洲的熱帶國家和地區[27]。栽培的緯度范圍在北緯20°和南緯20°之間,但主要分布在15°以內地區。我國腰果種植主要集中在云南、海南等地。
腰果仁有豐富的營養價值,用途十分廣泛,可作為甜品、點心、菜肴的原材料。副產品主要有果梨、果殼液等,果殼液被作為重要的工業原料被大量使用,果梨可以食用,用在果酒、果醬、腌菜等[28]。因此,種植腰果可以帶來很大的經濟效益、社會效益和生態效益。在中國熱帶地區脫貧攻堅和鄉村振興中起到重要作用。腰果對低溫比較敏感,當溫度低于15 ℃時生長緩慢,在8 ℃時受到嚴重傷害,此前,海南等地的腰果因低溫受災嚴重。目前,對腰果的研究主要集中在生理層面,對于分子層面的研究較少。抗逆基因已經在許多物種中進行了挖掘和應用,本研究將腰果作為實驗材料,對腰果的CBF 基因家族進行相關生物信息學分析,分離鑒定得到AoCBF 基因,為后續深入開展腰果耐寒性研究提供基礎數據。
1 材料與方法
1.1 材料
將保存于中國熱帶農業科學院種質庫的腰果種質資源CJSX 作為實驗材料。通過篩選得到有活力的種子,將篩選的種子進行催芽,長出真葉后移栽至育苗盤中,在溫度為25 ℃,16 h 光照,2800 lx 的環境下生長。生長到6 片真葉時,移栽至人工氣候屋,保持光照條件不變,對照溫度25 ℃(CK)不變,溫度分別調至8 ℃、16 ℃,2個溫度下處理時間為6 h、24 h,共4 個處理(T1:8 ℃ 6 h,T2:8 ℃ 24 h,T3:16 ℃ 6 h,T4:16 ℃24 h)。將低溫處理過的腰果葉片采摘,液氮處理,送公司進行Illumina 轉錄組測序。
1.2 方法
1.2.1 腰果CBF 基因家族的鑒定和分析 在擬南芥資源網站TAIR(https://www.arabidopsis.org/)上獲取相關AtCBFs 蛋白序列。將得到的AtCBFs蛋白序列進行本地BLAST 分析, 相關參數E_value 設置為10e–50;蛋白的篩選標準為identity≥60%,align_ratio≥60%。將蛋白是否含有AP2結構域作為重要的判斷依據,使用NCBI-CDD 數據庫與Pfam_scan 軟件進行驗證,由此得到具有AP2 結構域的候選腰果CBF 基因。利用在線軟件Expasy( https://web.expasy.org/protparam/)分析腰果AoCBF 蛋白序列,對每個AoCBF 蛋白的等電點、長度、相對分子質量進行預測[29]。使用Cell-PLoc 2.0(http://www. csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)在線軟件對候選的腰果CBF 基因進行亞細胞定位預測[30]。
1.2.2 AoCBF 蛋白保守結構域和保守基序(motif)分析 對每個AoCBF 氨基酸序列使用MEGA-X軟件進行比對,使用MEME 在線軟件(https://meme-suite.org/meme/)對AoCBF 蛋白保守結構域對進行分析。對相關AoCBF 蛋白的保守基序進行分析[31],設置為5 個基序,設置基序為10~200個氨基酸長度,其他參數均為默認值。使用SWISS軟件對AoCBF 蛋白進行三維結構預測[26]。
1.2.3 AoCBF 蛋白系統進化分析為了進一步研究腰果和擬南芥之間的進化關系,將6 個AoCBF 蛋白和6 個AtCBF 蛋白采取Neighborjoining(NJ)法在MEGA-X 軟件上構建系統發育樹[24],Bootstrp method 參數值設為1000。
1.2.4 AoCBF 基因上游序列的順式作用元件分析從 JGI(https://phytozome-next.jgi.doe.gov/info/Aoccidentalev0_9)網站上獲取AoCBF 基因上游2000 bp 的序列,利用PlantCARE 軟件對其進行順式作用元件預測分析[32]。
1.2.5 AoCBFs 在冷脅迫下的表達模式 用TBtools(v1.082)軟件提取轉錄組數據庫中腰果CBF 基因家族的CDS 序列,并通過IDT(https://sg.idtdna.com/pages)在線網站進行引物設計(表1)。使用多糖多酚植物總RNA 提取試劑盒(成都福際生物技術有限公司)按照說明書從每個樣品中提取總RNA,并對其濃度和純度進行檢測,結果顯示A260/A280 和A260/A230 的參數均在2.0 左右,表明RNA 無明顯的DNA 和蛋白污染,此外,通過瓊脂糖電泳也檢測RNA 的完整性。隨后通過逆轉錄試劑盒(蘇州莫納生物科技有限公司)按照說明書合成cDNA 第一鏈并檢測其濃度,稀釋后備用。并使用2–ΔΔCT 方法計算每個AoCBFs 的最終相對表達水平。使用GraphPad Prism 8 軟件制圖,每個處理3 個生物學重復。
2 結果與分析
2.1 腰果CBF 基因家族成員的鑒定和分析
用轉錄組數據庫和6 個已知的擬南芥AtCBFs序列進行本地BLAST 分析,共得到15 個注釋的DREB 基因。經過結構域分析,鑒定出11 個腰果CBF 基因。設計特異性引物進行PCR 擴增,獲得6 條腰果序列(AoCBF1~AoCBF6)。
理化性質分析結果顯示,AoCBFs 編碼的氨基酸序列存在較大差異,預測蛋白的長度為189~334 aa,最長的為AoCBF1,最短的為AoCBF4。蛋白分子量介于21.31~37.89 kDa 之間;6 個AoCBFs全部呈酸性,其理論等電點(pI)介于4.93~5.79之間。通過亞細胞定位分析得出所有AoCBFs 都有核定位信號(表2)。
2.2 AoCBF 蛋白保守基序分析
保守基序(motif)預測結果表明(圖1),AoCBFs 含有5 個保守motifs。motif 1、motif 2存在所有AoCBFs 中,表明2 個基序高度保守。AoCBF1只包含motif1、motif2,AoCBF2、 AoCBF3、AoCBF4、AoCBF5、AoCBF6 包含motif1、motif2、motif3、motif5,原因可能是串聯重復過程中堿基丟失造成的。對上述保守基序檢測發現motif 1 含有AP2 結構域。根據其在結構上有所不同,推測不同AoCBF 在功能上存在一定差異。
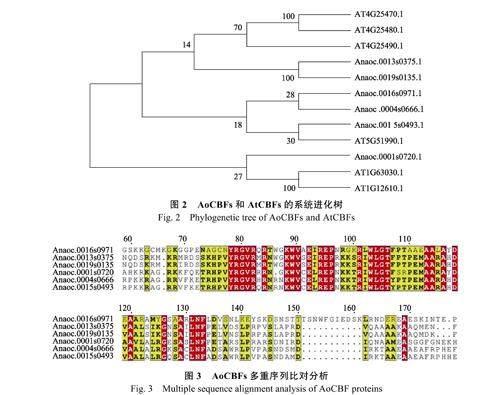
2.3 腰果和擬南芥CBF 蛋白的系統發育樹分析
將6 個AoCBFs 氨基酸序列和6 個AtCBFs氨基酸序列構建系統發育樹。結果顯示,上述CBF氨基酸序列可分成A、B 兩個亞家族。AoCBFs在A 組有1 個,B 組有5 個。AtCBFs 在A 組有2個,B 組有6 個(圖2)。
2.4 AoCBFs 三維結構及序列特征分析
6 個AoCBF 蛋白序列相似性在46%~58%之間。一段長度為60 個氨基酸左右較為保守的序列在N 段被發現,軟件預測其為CBF 蛋白的DNA結合結構域(圖3)。AoCBFs 的identity 最高為AoCBF4 和AoCBF6。最高的同源性為58.21%。對AoCBFs 進行三維結構建模分析,1 個α-螺旋和2 個β-折疊構成AoCBFs 的三維結構。
2.5 AoCBFs 上游序列的順式作用元件分析
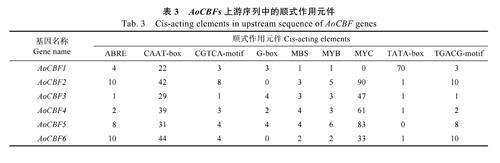
利用PlantCARE 軟件注釋AoCBFs 基因上游2000 bp 啟動子區域序列,在其上游發現了參與激素反應的相關順式作用元件(茉莉酸甲酯、脫落酸、赤霉素、生長素、水楊酸有關的順式作用元件)、參與低溫、干旱和高鹽誘導脅迫相關的響應元件、光響應元件。此外,普遍存在于啟動子與增強子區的核心元件、生長發育相關調控順式作用元件也被發現。結果表明,AoCBF 基因在抵御非生物脅迫、生長發育、激素調控中發揮重要作用(表3)。
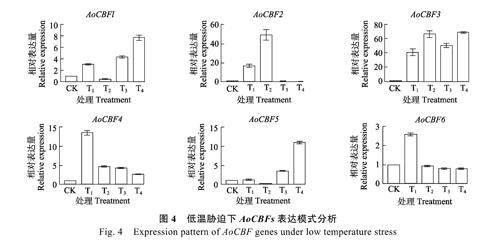
2.6 AoCBFs 在冷脅迫下的表達模式
為了探明AoCBFs 在冷脅迫下的表達模式,對AoCBFs 進行實時熒光定量PCR 分析。結果表明,經8 ℃和16 ℃低溫誘導處理6 h 和24 h 后,除了AoCBF5 在8 ℃時相對表達量變化較少,在16 ℃表達變化較大,其他基因在8 ℃的表達量均顯著升高。表明AoCBF 基因參與響應冷脅迫相關生物學過程(圖4)。
3 討論
腰果是典型的熱帶果樹,對低溫極敏感。主要在我國熱區的海南、云南等地種植。但是近年來冬季寒潮頻發,溫度降低使腰果受到冷害的問題愈加嚴重。植物中CBF 家族在面對低溫時發揮調控作用,減少低溫對植株的損傷。因此,本課題組對腰果CBF 家族進行生物信息學分析,以期發現腰果CBF 基因在低溫脅迫下的調控機制。
本研究共鑒定獲得6 個AoCBF 基因。并對其理化性質、亞細胞定位、系統發育進化樹、蛋白三維結構、順式作用元件以及對低溫脅迫下的表達方式進行研究。由于AoCBFs 的等電點為4.93~5.79,蛋白呈酸性,故在酸性亞細胞環境下,AoCBF 蛋白可能會發揮作用。AoCBFs 可以分為2 個亞族:AoCBF1 為A 族,AoCBF2、AoCBF3、AoCBF4、AoCBF5、AoCBF6 為B 族。而B 族中AoCBF3、AoCBF4 具有更高親緣性,AoCBF2、AoCBF5、AoCBF6 可能具有類似的調控方式。
目前CBF 基因在十字花科植物、禾本科植物、豆科植物、漆樹科植物、茄科植物中均有研究。最早是STOCKINGER 等[33]在模式植物擬南芥中克隆得到相關CBF 基因。擬南芥的6 個家族成員,受外源ABA 誘導的有CBF1、CBF2 和CBF3。CBF1 和CBF3 的表達受CBF2 的負調控。CBF4不能響應低溫脅迫誘導但是能夠響應干旱和ABA誘導[34]。DREB1E 和DREB1F 均可以受高鹽脅迫誘導。本研究在6 個AoCBFs 啟動子上不僅篩選到響應低溫誘導、干旱誘導、光響應元件有關的順式作用元件,而且篩選到與茉莉酸、生長素、水楊酸、脫落酸、赤霉素等激素有關的順式作用元件。后續可開展分子對激素和冷信號調控的相關機制。
AoCBF 基因家族6 個成員均不存在內含子,所以AoCBF 基因為內含子缺失型。內含子缺失型基因在面對脅迫時,可通過減少轉錄處理的方法提高轉錄本積累的效率[35]。
本研究發現,腰果經過冷脅迫處理后,6 個AoCBF 基因均對低溫產生響應,除了AoCBF1、AoCBF3、AoCBF4、AoCBF5 在16 ℃表達顯量著提高,其余的基因均在8 ℃時表達量顯著提高。
相較于傳統的雜交育種、誘變育種等方式,基因工程技術有高效率、短周期、提高植物抗逆性等優勢。科研工作者可以利用基因克隆技術、基因重組技術、轉基因技術等基因工程技術提升植物抵御低溫脅迫的能力。本研究通過對AoCBF基因家族進行生物信息學分析,分析AoCBF 基因在低溫脅迫下的調控機制,為未來的腰果抗性分子輔助育種研究工作奠定基礎。
