無核荔枝僵果病的病原菌鑒定、入侵途徑及快速檢測技術
2024-06-05 15:00:32王偉博李松剛胡美姣曹學仁李敏李煥苓李芳高兆銀
熱帶作物學報 2024年3期
關鍵詞:檢測
王偉博 李松剛 胡美姣 曹學仁 李敏 李煥苓 李芳 高兆銀

關鍵詞:無核荔枝;僵果病;病原菌;鑒定;檢測
荔枝(Litchi chinensis Sonn.)是原產于我國的重要的熱帶果樹之一。無核荔枝是海南選育的地方特色品種,其果大皮紅,果肉晶瑩、口感清脆無渣,食用時不吐核,備受消費者喜愛。隨著荔枝栽培技術的提高,并蒂雙生的無核荔枝成為無核荔枝生產的主流,在果實長至花生米大小時,通過疏除單果、有核果實,保留并蒂成對的無核果實,實現并蒂雙生無核荔枝的生產,其售價高,是單果售價的2~4倍,經濟效益顯著[1-3]。近年來無核荔枝上發生了一種為害果實的新病害,與荔枝常見的果實病害,如炭疽病、霜疫霉病、酸腐病引起的果皮褐變,在樹上或者采后果實腐爛不同,這種新病害主要為害幼果,果實長至蠶豆大小時,發病果實的外果皮局部顏色變暗,剖開后變暗部位內果皮發生不規則褐變,果實不再膨大,形成無肉僵果,命名為無核荔枝僵果病。該病害僅導致幼果發育停止、內果皮呈灰褐色,褐變位置多發生在果實縱莖中間位置,較少發生在果柄和果頂端部位,這種蠶豆大小的僵果一直維持到果實成熟期不腐爛,果皮比正常果實提前3~7 d 變紅。如果并蒂雙生無核荔枝果實中有1個果實發生這種病害,其經濟價值將顯著降低。2019—2021 年本課題組調查該病害的發病率在5%~15%之間,嚴重影響并蒂雙生果實的生產,造成嚴重的經濟損失。該病害的癥狀與GAO 等[4]報道的桂花香荔枝變紅鐮刀菌(Fusarium incarnatum)果腐病的癥狀相似,不同的是變紅鐮刀菌果腐病的癥狀為果實外觀無明顯病變表現,近果柄處果皮變硬,剖開后內果皮果柄周圍呈灰褐色褐變,發病部位在果柄處或近果柄處居多,變紅鐮刀菌果腐病不影響桂花香荔枝果肉的正常生長和食用品質,果實可以正常成熟。而目前國內外均未見無核荔枝僵果病的報道。本研究對無核荔枝僵果病的病原菌進行分離、鑒定,并研究其入侵途徑和分子快速檢測、診斷技術,為該病的鑒別和高效防治提供技術支撐。
1 材料與方法
1.1 材料
無核荔枝果實(感病果實和健康果實)采自海南澄邁無核荔枝基地。
1.2 方法
1.2.1 病原菌的分離采回感病無核荔枝果實后,參照胡美姣等[5]的方法進行病原菌分離。從PDA 培養分離物中挑取菌絲置于28 ℃恒溫條件下培養2~3 d,長出菌絲后在菌落邊緣用無菌鑷子剪取形態單一的小菌絲塊接種至PDA 培養基進行純化。培養6 d 待菌落長出孢子后采用標記法獲得單孢分離物,用于后續致病性試驗、病菌鑒定和檢測等研究。
1.2.2 致病性測定致病力測定方法參考胡美姣等[6]的方法,選取果實縱經約1 cm,大小一致(約花生米大小)的健康果實,采用無刺傷接種和昆蟲針刺傷果面接種2 種方式。用打孔器將培養7 d的單孢分離菌株PDA 培養基打成直徑為5 mm 的菌餅,菌絲一面貼到荔枝果皮上,用脫脂棉保濕,24 h 后去除脫脂棉,每個菌株接種12 個果實,3 d后統計發病情況,共接種64 株單孢分離物,以空白PDA 培養基瓊脂餅為對照(CK)。
1.2.3 病原菌鑒定(1)形態學鑒定。培養7 d后,在徠卡生物顯微鏡下觀察病原菌分生孢子和產孢結構,根據形態學進行病原菌鑒定。(2)分子鑒定。使用CATB 法提取病原菌DNA[7],進行ITS、RPB2、EF-1α 擴增。ITS 擴增引物序列參照WHITE 等[8]設計的真菌rDNA-ITS 序列,PCR 條件參考胡美姣等[5]的方法。RPB2、EF-1α 基因的引物序列及PCR 條件參照WANG 等[9]的方法。凝膠電泳檢驗合格后將產物送生工生物工程(上海)股份有限公司測序,基于ITS 測序結果在NCBI 上進行比對,確定病原菌的屬,再從NCBI上下載相關屬種的序列及外群序列,基于RPB2、EF-1α 構建系統發育樹,確認病原菌的生物學種類[10]。
1.2.4 病原菌的侵染途徑試驗田間幼果經農事活動碰傷、自然風力較大擦傷以及昆蟲口器刺傷等傷害后,果皮受傷部位會流出明顯的黑色汁液。選取縱徑為10 mm 的3 串健康果實,每串4 個果實,共12 個流黑汁的果實進行侵入途徑試驗。接種方法和發病統計同1.2.2。以15 cm×20 cm 大小的紗網套住果穗,每個紗網放入5 只荔枝蝽,24 h后選擇果皮變黑的果實接種病原菌,以表面無損傷的果皮接種病原菌為對照(CK),探究病原菌的侵染途徑。
1.2.5 病原菌的分子檢測以伯氏鐮刀菌和分離獲得的其他鐮刀菌屬真菌基因組DNA 為模板,利用鐮刀菌屬通用引物TUB2F/TUB2R 對其進行PCR 擴增,PCR 產物委托生工生物工程(上海)股份有限公司進行純化和測序。根據測序獲得的序列信息,采用MEGA 11 軟件進行序列比對,找到伯氏鐮刀菌和其他鐮刀菌基因序列的差異性位點,利用NCBI 設計檢測伯氏鐮刀菌的特異性引物序列,選用契合度最高的一對引物F1D/R1D 進行PCR 擴增,根據凝膠電泳結果判斷引物的特異性。引物由生工生物工程(上海)股份有限公司合成。為了進一步鑒定篩選獲得的伯氏鐮刀菌檢測引物的特異性,以分離獲得的所有菌株為測試對象,進行PCR 檢測,根據凝膠電泳結果判斷引物的特異性;從盛花期后第10 天開始取樣(不同生長期共取9 次樣,編號為1~9),每個時期取樣18 個單果,表面經75%酒精消毒30 s,用滅菌濾紙吸干水分,單果包裝置于–80 ℃保存備用。以不同時期的單果DNA為檢測目標進行PCR 檢測,使用Plant Genomic DNA Kit 試劑盒[天根生化科技(北京)有限公司]進行果實DNA 的提取,根據擴增結果判斷病原菌入侵情況。
1.3 數據處理
試驗數據采用Excel 軟件進行統計分析,用SAS 9.1 軟件進行差異顯著性分析(P<0.05)。
2 結果與分析
2.1 病原菌的分離、純化與致病性測定
從發病無核荔枝果實中分離獲得64 株菌株,根據形態特征分為14 種,每種菌株接種1 株,所有菌株均未能侵染無刺傷果實,導致刺傷果實發病的共有8 株,分為3 個種類,菌株編號分別為SLZ-1-2、02、32。其中致病性最強的病原菌株為SLZ-1-2,發病癥狀與自然發病果實癥狀一致(圖1)。從刺傷接種發病果實中,再次分離獲得的菌株與SLZ-1-2 一致,表明菌株SLZ-1-2 為本研究的主要致病菌。
2.2 病原菌的形態學鑒定
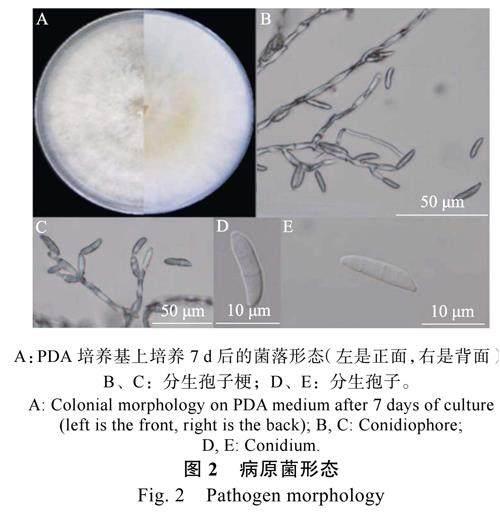
病原菌菌株SLZ-1-2 在PDA 平板培養基上生長的菌絲密集,氣生菌絲呈淡黃色,菌落背面呈淡棕色(圖2)。分生孢子的形態呈鐮刀型,整體稍彎曲,1~5 個分隔。根據培養特征和形態特征,鑒定該病原菌為鐮刀菌(Fusarium)。
2.3 病原菌的分子鑒定
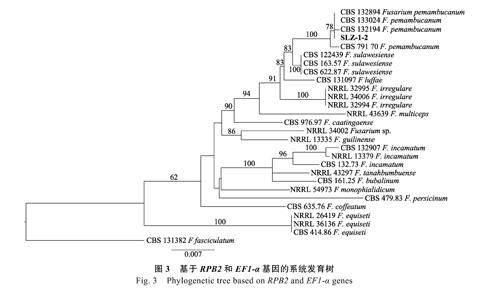
將病原菌菌株SLZ-1-2 的RPB2 和TEF1 序列與GenBank 中下載的27 個菌株的相關基因序列進行同源性比對,以PhyloSuite 進行拼接,用Cipres 以伯氏鐮刀菌(F. pernambucanum)為外群構建系統發育樹(圖3)。本研究致病菌株SLZ-1-2與伯氏鐮刀菌以100%的高支持率聚為一支,支持形態學鑒定結果,因此,基于形態學和分子生物學特征,將該致病菌株鑒定為伯氏鐮刀菌(F.pernambucanum)。
2.4 病原菌的入侵途徑
在田間無刺傷接種病原菌時,果實不發病,機械損傷接種后發病率為92.24%;刺吸式害蟲為害后,發病率為82.27%(圖4)。因此,無核荔枝果實表面出現損傷時病原菌能入侵致病。
2.5 伯氏鐮刀菌引物的特異性驗證
用 F1D(5-CCTGGCAGGAGTGATGAAAC-3)/R1D(5-AATCAGACCGGTCAGTGCGT-3)對伯氏鐮刀菌和分離獲得的其他鐮刀菌屬真菌及非鐮刀菌屬真菌進行PCR擴增以驗證引物的特異性,結果表明,僅伯氏鐮刀菌基因組DNA 可以擴增出1 條220 bp 左右的特異性片段,與預期目標條帶大小一致,而從無核荔枝上分離供試的3種鐮刀菌屬真菌和其他非鐮刀菌屬真菌無目的條帶出現,CK 也無目的條帶出現(圖5,圖6),試驗結果驗證了該對引物的特異性,可用于田間檢測病原菌。
2.6 發病果實的分子檢測
在田間隨機采集無核荔枝僵果果實,利用特異性引物F1D/R1D 對其進行PCR 擴增,結果顯示,僵果均可以擴增出目標條帶,而CK 無條帶產生。說明該特異性引物可以用于生產中無核荔枝僵果病病原菌及病果的快速檢測(圖7)。
2.7 病原菌入侵時期
從表 1 可以看出,無核荔枝幼果期帶菌率較高,特別是果實縱徑小于10 mm 時,果實帶菌率比較高,隨著荔枝果實的生長,果實角質層逐漸增厚,帶菌率逐漸降低。分別以田間隨機采集的果實和伯氏鐮刀菌基因組DNA 為模板,利用特異性引物F1D/R1D 對其進行PCR 擴增,結果顯示,部分無癥狀幼果可以擴增出目標條帶,其中以果實縱徑為3.2~4.8 mm 時期檢出率最高,說明此階段為病原菌的主要入侵時期,果實大于9.8 mm后,病原菌入侵逐漸降低。
3 討論
鐮刀菌屬真菌病害是造成植物發生病害的主要病原之一,如枯萎、腐爛和枯死[11-15]。OMAR等[16]報道了由伯氏鐮刀菌(F. pernambucanum)引起的馬來西亞芒果葉斑病。GUO 等[17]報道了由伯氏鐮刀菌引起的我國芒果的葉斑病。ZHANG等[18]報道了由伯氏鐮刀菌引起的甜瓜僵果病,發病部位呈灰褐色、水漬狀,發病后期可導致整個果實腐爛,果實表面布滿白色至玫瑰紅色的霉層。LU 等[19]報道了由伯氏鐮刀菌引起的李葉枯病,發病葉片邊緣或尖端出現不規則的棕色斑點,發病后期,整個葉片枯萎褐變、卷曲或脫落。本研究發現引起無核荔枝的新型僵果病的病原菌為伯氏鐮刀菌。該病原菌主要為害幼果,發病時幼果果實外觀無明顯變化,但無核荔枝完成疏果后,果實長至蠶豆大小時,發病果實的外果皮變暗,剖開后內果皮發生不規則褐變,果實不再膨大,形成僵果。
GUO 等[17]的研究表明,由伯氏鐮刀菌引起的芒果葉斑病在葉片表面存在傷口時才能入侵致病。本研究中也發現,通過農事操作,如打藥、施肥等人工擦傷,大風后機械摩擦損傷,荔枝蝽為害等方式,在荔枝果實表面造成傷口時,伯氏鐮刀菌則入侵致病,果皮無傷口時不致病。經過全面的田間調查,未發現該病原菌為害無核荔枝的嫩梢和葉片。
分子檢測技術以其高效、靈敏、簡便的特點,已廣泛應用于植物病害的檢測[20]。李亭潞等[21]利用環介導等溫擴增(LAMP)技術建立了荔枝霜疫霉和炭疽病的快速檢測技術。而利用PCR 或qPCR(quantitative real-time PCR)技術可檢測到的植物病原已達50 余種[22]。本研究根據伯氏鐮刀菌的序列信息,利用MEGA 11 軟件與其他鐮刀菌進行序列比對,找到差異性位點,合成引物F1D/R1D 進行PCR 擴增,對帶致病菌果實的檢測率達到100%。
無核荔枝是海南本地選育的優良品種,其獨特的品質和生產模式,以及高價值的經濟效益已經成為海南重要的農產品地理標志產品。無核荔枝新型僵果病果實的果皮受傷后,由伯氏鐮刀菌入侵為害,該菌主要通過農事操作、風、刺吸口器昆蟲造成荔枝果皮傷口后侵入感染,果實縱徑為3.2~4.8 mm 時為病原菌的主要入侵時期。本研究建立的無核荔枝僵果病的快速檢測方法,不僅靈敏度高,能區分其他屬病原菌,也能區分親緣關系較近的鐮刀菌屬真菌。可為僵果病的早期預防,遏制病害的發生和傳播提供技術保障。但該病原菌是否為害葉片,以及是否入侵如蒂蛀蟲等其他因素造成的幼果傷口有待進一步研究。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
