
外源赤霉素處理影響湖北梣種子萌發過程的生理機制
2024-06-25 21:14:10趙怡寧劉彥宏鄭鵬麗周明芹
湖北農業科學 2024年5期
關鍵詞:研究
趙怡寧 劉彥宏 鄭鵬麗 周明芹
收稿日期:2023-12-07
基金項目:國家自然科學基金資助項目(31200528)
作者簡介:趙怡寧(1998-),女,湖南常德人,在讀碩士研究生,研究方向為園林植物,(電話)19936731678(電子信箱)814601873@qq.com;通信作者,周明芹(1978-),女,湖北荊州人,教授,博士,主要從事園林植物種質資源的利用與創新、植物系統分類等方面研究,(電子信箱)500548@ yangtzeu.edu.cn。
趙怡寧,劉彥宏,鄭鵬麗,等. 外源赤霉素處理影響湖北梣種子萌發過程的生理機制[J]. 湖北農業科學,2024,63(5):70-76,83.
摘要:以常溫貯藏2年的湖北梣(Fraxinus hupehensis Chiú. Shang et Su)種子為研究對象,試驗前去除種子胚根端被覆組織以露出胚根。以清水作對照,用150 mg/L赤霉素浸種24 h后,測定種子萌發過程中生理生化指標及內源激素含量的變化。結果表明,赤霉素浸種可以提高種子發芽率,赤霉素處理后種子發芽率為24.67%,對照為16.67%;萌發過程中,赤霉素處理后種子可溶性糖含量呈先上升再下降的變化趨勢;可溶性蛋白質含量呈下降趨勢,各時期均顯著高于對照;超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性、過氧化氫酶(CAT)活性整體呈上升趨勢;赤霉素處理后的種子內源茉莉酸甲酯(JA-Me)含量均顯著低于對照,更有利于內源赤霉素(GA3)、玉米素核苷(ZR)含量的增加;生長素比脫落酸(IAA/ABA)與赤霉素比脫落酸(GA3/ABA)和玉米素核苷比脫落酸(ZR/ABA)分別在0.01和0.001水平上呈顯著正相關;GA3/ABA與ZR/ABA在0.001水平上呈顯著正相關。赤霉素浸種顯著提高了湖北梣種子的發芽率,處理后的種子平衡保護酶活性系統較強,質膜受損延緩,抗逆能力較好。較低內源JA-Me含量和較高內源GA3、ZR有利于種子萌發的啟動和休眠的解除。
關鍵詞:湖北梣(Fraxinus hupehensis Chiú. Shang et Su); 種子; 生理生化; 萌發; 赤霉素
中圖分類號:S792.41???????? 文獻標識碼:A
文章編號:0439-8114(2024)05-0070-07
DOI:10.14088/j.cnki.issn0439-8114.2024.05.013??????????? 開放科學(資源服務)標識碼(OSID):
Physiological mechanisms of exogenous gibberellin treatment affecting the seed germination process of Fraxinus hupehensis Chiú. Shang et Su
ZHAO Yi-ning, LIU Yan-hong, ZHENG Peng-li, ZHOU Ming-qin
(College of Horticulture and Gardening, Yangtze University, Jingzhou? 434000, Hubei,China)
Abstract: The seeds of Fraxinus hupehensis stored at normal temperature for 2 years were taken as the research object. Before the experiment, the radicle end covering tissue was removed to expose the radicle. Taking water as a control, after soaking the seeds with 150 mg/L gibberellin solution for 24 h, the physiological and biochemical indexes and the contents of endogenous hormones in seeds during germination were measured. The results showed that gibberellin treatment could enhance seed germination rate, with a germination rate of 24.67% after gibberellin treatment compared to 16.67% in the control group. During germination, gibberellin-treated seeds exhibited a trend of initially increasing and then decreasing in soluble sugar content. Soluble protein content showed a decreasing trend but remained significantly higher than the control group at all stages. Superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) activities showed an increasing trend on the whole. The content of endogenous jasmonic acid methyl ester (JA-Me) in seeds after gibberellin treatment was significantly lower than that in the control, which favored an increase in endogenous gibberellin (GA3) and zeatin riboside (ZR) content. The ratios of gibberellin to abscisic acid (GA3/ABA) and zeatin riboside to abscisic acid (ZR/ABA) were significantly positively correlated with the ratio of indole-3-acetic acid to abscisic acid (IAA/ABA) at 0.01 and 0.001 levels, respectively, and GA3/ABA was significantly positively correlated with ZR/ABA at 0.001 level. Gibberellin immersion significantly increased the germination rate of Fraxinus hupehensis seeds, and the seeds treated with gibberellin exhibited a stronger balance of protective enzyme activity system, delayed membrane damage, and the resistance was better. The lower levels of endogenous JA-Me and the higher levels of endogenous GA3 and ZR favored the initiation of seed germination and the release of dormancy.
Key words: Fraxinus hupehensis Chiú. Shang et Su; seed; physiology and biochemistry; germination; gibberellin
湖北梣(Fraxinus hupehensis Chiú. Shang et Su)為木犀科白蠟屬的落葉喬木,原生于湖北省大洪山余脈的京山市、鐘祥市一帶,屬于湖北省特有珍稀新品種保護植物和國家Ⅱ級珍稀瀕危植物[1,2]。其樹姿豐盈,耐修剪,觀賞價值高,抗逆性強,在園林建設中應用廣泛[2,3]。孔令雷等[4]的研究表明湖北梣提取物有抗癌作用,具有藥用價值。有研究發現湖北梣種胚被覆組織機械阻礙了種子萌發[5]。楊湘虹等[6]認為休眠可能是幾種因素共同作用的結果,內源抑制物的存在與胚生理發育未完全是造成湖北梣種子休眠的主要原因。湖北梣種子深度休眠的特性限制了其栽培與應用,而關于其種子萌發的生理機制研究卻鮮有報道。
赤霉素(GA3)作為一種植物激素,影響植物生長和發育的許多方面[7],并已被廣泛研究。GA3能打破種子休眠、促進萌發[8]。羅富成等[9]發現用赤霉素對種子進行處理,可提高其發芽率,類似的結論在蒺藜苜蓿種子[10]和海棗種子[11]的研究中也有體現。Song等[12]研究發現外源GA3提高了湖北梣種子的萌發率、活力和吸水率,改變了其內源激素含量。植物內源激素對種子的生長發育具有不同的效應,其含量變化也客觀反映了種子的代謝活力,不同激素之間的協同、相互作用,共同調控種子萌發[13-15]。GA3在打破種子休眠后所引起的內源激素變化能反映種子萌發的程度,是評價種子萌發的關鍵指標[16-19]。綜上,有關外源GA3處理在湖北梣種子萌發過程中的變化研究尚為鮮見。
本研究通過施用外源赤霉素,測定種子不同時間的生理生化指標和內源激素變化,探究內源激素在種子萌發中的作用,借以初步研究抑制湖北梣種子萌發的內部因素,以期提出切實有效的休眠破除方法,為后續湖北梣種子內物質代謝、內源激素調控及信號傳導等方面研究做鋪墊,為其繁育及開發應用提供理論依據。
1 材料與方法
1.1 材料及其處理
湖北梣種子采集于湖北省宜昌市三峽植物園,將種子保留果翅,常溫貯藏。去翅處理后,各處理選取1 000粒果粒飽滿的種子用于發芽試驗。試驗開始前,將種子用高錳酸鉀(0.2%)消毒后去除胚根端被覆組織,以150 mg/L赤霉素浸種24 h,用清水作對照。
1.2 方法
用電導法測定種子的質膜透性變化,用蒽酮比色法測定種子可溶性糖含量[20],采用考馬斯亮藍法[21]測定可溶性蛋白質含量,采用氮藍四唑法[22]測定超氧化物歧化酶(SOD)活性,采用愈創木酚顯色法測定[19]過氧化物酶(POD)活性,采用紫外吸收法[23]測定過氧化氫酶(CAT)活性,用電導儀測定浸泡液和對照溶液的電導率。
取處理后的種子50粒均勻散布于鋪有2層中速定性濾紙、直徑9 cm的玻璃培養皿中,置于4 ℃冰箱中低溫層積處理,每處理20個重復,隨機選取3個重復標記為萌發試驗處理,剩下17個重復用于內源激素含量測定。每3 d灑水1次,保證培養皿中濾紙處于濕潤狀態,每30 d取樣1次,直至試驗結束。
取萌發試驗的0、30、60 d種子各1 g研磨成粉末,加入40 mL 80%冰甲醇,4 ℃冰箱下浸提8 h以上,離心過濾,取上清液,用旋轉蒸發儀將提取液濃縮至原有體積的1/3。加入0.5 mol/L Na2HPO4,將pH調至8.00,用石油醚萃取3次后加水、交聯聚乙烯吡咯烷酮(PVPP)置于搖床上振蕩30 min,過濾后加入2 mol/L檸檬酸調pH至3.00,乙酸乙酯萃取3次,酯相于真空濃縮儀中蒸干,殘留物用3 mL pH=3.00的Na2HPO4溶解。將收集液過C18固相萃取小柱進一步純化,并將收集的洗脫液于旋轉蒸發儀上蒸干,用色譜純甲醇溶解殘留物于2 mL離心管中,定容至1 mL,過0.45 μmL微孔濾膜待測。測定內源激素有茉莉酸甲酯(JA-Me)、赤霉素(GA3)、玉米素核苷(ZR)、生長素(IAA)、蕓苔素(BR)、脫落酸(ABA)。
1.3 數據分析
采用Excel軟件計算數據,利用IBM SPSS Statistics 26、GraphPad Prism軟件和R語言進行顯著性及相關性分析并制圖。
2 結果與分析
2.1 湖北梣種子萌發過程中的生理生化分析
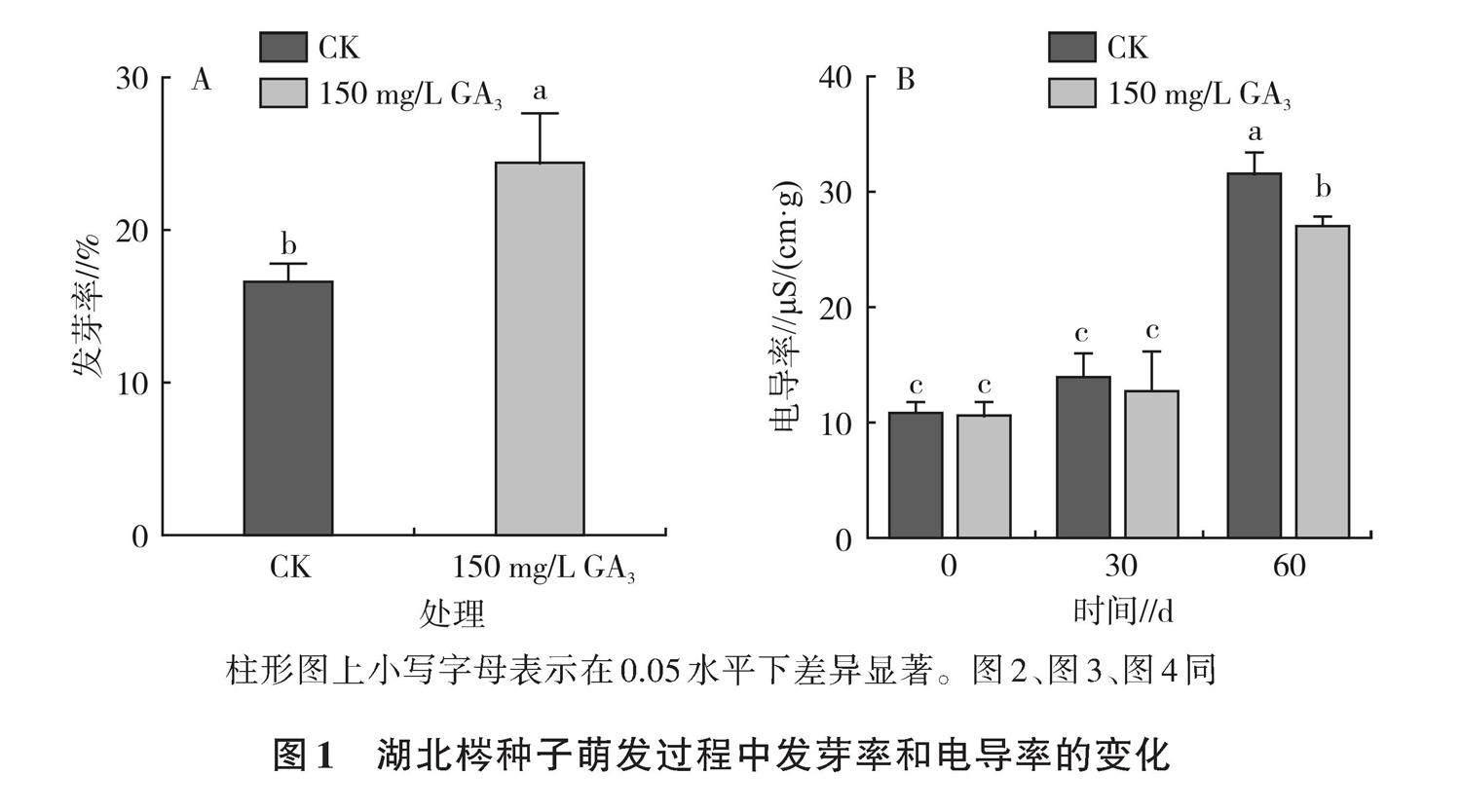
2.1.1 湖北梣種子萌發過程中發芽率和電導率的變化 如圖1A所示,外源赤霉素處理能夠顯著提高湖北梣種子的發芽率。種子經150 mg/L赤霉素溶液浸泡24 h后,發芽率為24.67%;與清水對照(發芽率為16.67%)相比,其發芽率顯著提高了8個百分點。電導率反映種子活力的高低,如圖1B所示,0~30 d,湖北梣種子的電導率均呈較低水平,赤霉素浸種后種子電導率均低于對照,隨著時間的增加,電導率均呈增長趨勢,60 d電導率均較0 d顯著增加,分別為27.16 μS/(cm·g)和31.76 μS/(cm·g),赤霉素浸種處理與對照相比在各處理時間內均呈較低水平。
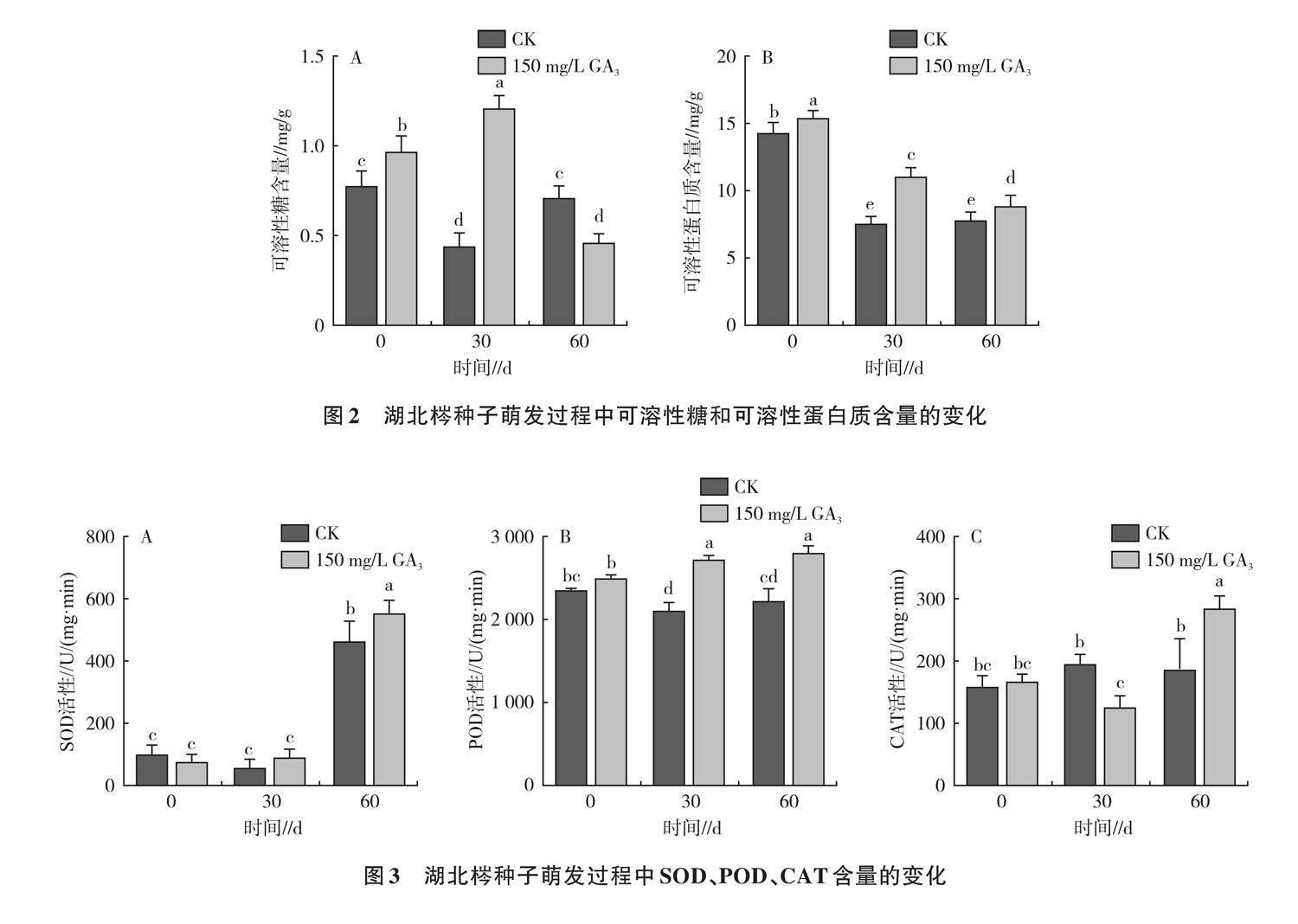
2.1.2 湖北梣種子萌發過程中可溶性糖和可溶性蛋白質含量的變化 可溶性糖含量能反映種子生理代謝過程中能量供應情況,如圖2A所示,赤霉素處理的種子可溶性糖含量呈先升后降的趨勢,與對照之間差異顯著。試驗30 d可溶性糖含量達到最大值,為1.21 mg/g,相比0 d增加了24.74%,60 d呈最低值,為0.46 mg/g,較0 d降低了52.58%。較低的可溶性蛋白質含量意味著種子內部代謝活動更為旺盛,如圖2B所示,對照和赤霉素處理可溶性蛋白質含量的變化均呈下降趨勢,0 d呈最大值(分別為14.81、15.36 mg/g),分別在30 d和60 d呈最低值(分別為7.53、8.87 mg/g),赤霉素處理可溶性蛋白質含量均顯著高于對照。
2.1.3 湖北梣種子萌發過程中SOD、POD、CAT活性的變化 SOD維持活性氧代謝系統的平衡,如圖3A所示,萌發0 d,各處理SOD處于較低水平;30 d時,對照和赤霉素處理SOD活性分別為53.47、88.42 U/(g·min);隨后SOD活性顯著上升,60 d時各處理SOD活性達到最高,赤霉素處理SOD活性顯著高于對照。赤霉素處理后POD活性比SOD活性表現更加敏感,POD是種子貯藏過程中的重要保護酶,如圖3B所示,赤霉素處理POD活性呈上升趨勢,萌發30、60 d時POD活性顯著高于對照。CAT可清除逆境下產生的過氧化氫,減弱植物所受的損傷,CAT活性的大小反映了細胞代謝的強度,如圖3C所示,隨著萌發時間的延長,赤霉素處理CAT活性隨著萌發時間的延長呈先降低后升高的趨勢,60 d時其活性為283.37 U/(g·min),較0 d增加了69.68%,與對照之間差異顯著。
2.2 湖北梣種子萌發過程中內源激素含量的動態變化
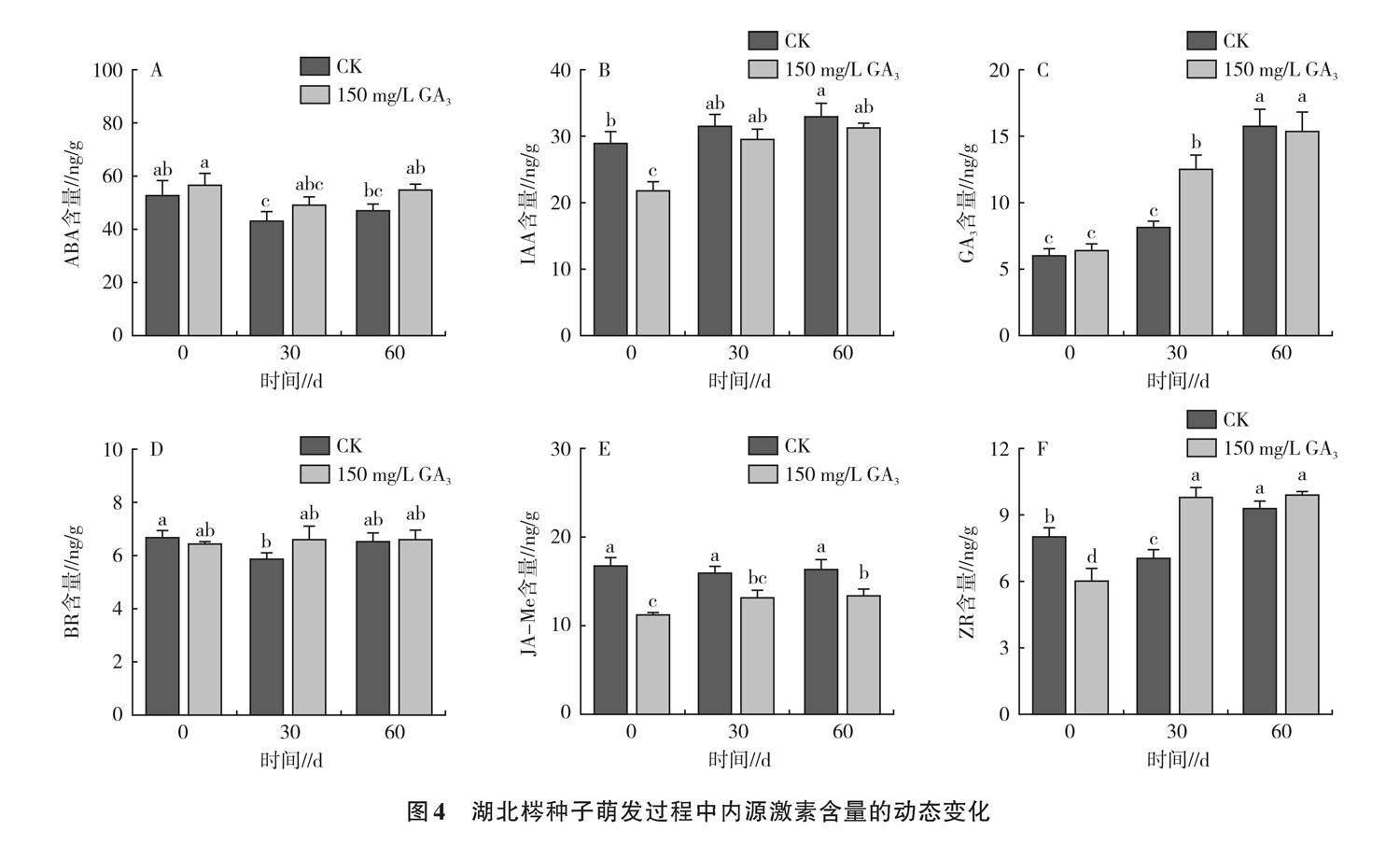
植物內源激素對植物的生理影響不是孤立發生的,二者之間存在復雜的關系網絡[24]。如圖4A所示,湖北梣種子內源ABA含量隨著萌發時間的推移,呈先減后增的趨勢,赤霉素處理ABA含量高于對照,在30 d時呈最低值(49.40 ng/g),60 d時赤霉素處理與對照ABA含量分別為55.32、47.24 ng/g,較0 d分別降低了2.76%和10.92%。表明試驗前期種子ABA合成受到抑制,一定程度上利于破除種子休眠,而試驗后期ABA含量的增加可能導致種子貯存合成物質增加,種子萌發受阻。此外,對內源激素進行相關性分析(圖5)得出,ABA與JA-Me和IAA分別在0.01和0.001水平上呈顯著負相關,相關系數(r)分別為-0.42和-0.48。
各處理種子IAA含量呈不斷上升趨勢,且赤霉素處理含量在每個試驗階段均低于對照(圖4B)。0 d時赤霉素處理IAA含量顯著低于對照,60 d時赤霉素處理和對照的IAA含量分別較0 d顯著增加了42.67%、13.67%,這表明隨著時間的推移,種子中的IAA含量在赤霉素處理下增加更為顯著,但較高含量的IAA可能對解除種子休眠起到了負面影響。從圖5可以看出,IAA與JA-Me在0.01水平上呈顯著正相關(r=0.69),與IAA/ABA在0.001水平上呈顯著正相關(r=0.87),與發芽率在0.01水平上呈顯著負相關(r=-0.48)。
如圖4C所示,各處理隨著萌發時間的增加,GA3含量不斷增加。30 d時,赤霉素浸種后種子GA3含量(12.61 ng/g)顯著高于對照(8.27 ng/g);60 d時,GA3含量在各處理間無顯著差異。說明外源赤霉素浸種可增加種子萌發過程中內源GA3含量,促進種子萌發。GA3與GA3/ABA、ZR/ABA分別在0.001和0.01水平上呈顯著正相關(圖5)。
蕓苔素(BR)作為世界第六大激素,研究表明,其兼具赤霉素、細胞分裂素和生長素的綜合功效,在平衡其他植物激素的同時還可以打破休眠、促進種子發芽[25]。赤霉素處理BR呈上升趨勢,對照呈先下降后上升的趨勢(圖4D)。表明采用赤霉素浸種較采用清水浸種更有利于增加種子在萌發過程中的BR含量。BR與IAA/ABA在0.05水平上呈顯著負相關(r=-0.45),與發芽率在0.05水平上呈顯著正相關(r=0.27)(圖5)。
如圖4E所示,0 d時,赤霉素處理JA-Me含量顯著低于對照32.6%,表明采用150 mg/g赤霉素浸種后種子質量更佳,具有較強抗逆性,而采用清水處理后種子損傷更大,品質更差,種子抗逆反應加強,JA-Me含量更高。各處理JA-Me含量隨著試驗時間推進不斷上升,赤霉素處理試驗各階段JA-Me含量均顯著低于對照,這可能是種子抗逆性逐漸降低,種子增加JA-Me含量來增強保護酶活性的原因,是種子對逆境做出的反應,赤霉素浸種后的種子更適應萌發環境。從圖5可以看出,JA-Me與種子發芽率在0.001水平上呈顯著負相關(r=0.90)。
玉米素核苷屬于細胞分裂素,具有促進細胞分裂、抑制器官老化、誘導花芽分化、打破種子休眠的作用。如圖4F所示,0 d時,采用赤霉素浸種種子的ZR含量低于采用清水的處理,可能是由于在試驗前清水浸種使種子內部生理反應更為活躍。30 d時,赤霉素處理種子的ZR含量顯著高于對照,較0 d增加62.68%,表明赤霉素處理更有利于促進萌發過程中ZR含量的增加。從圖5可以看出,ZR與GA3、GA3/ABA、ZR/ABA均在0.001水平上呈顯著正相關,r分別為0.81、0.78、0.87。
2.3 內源激素含量比例的變化
種子的萌發不僅與植物內源激素的絕對含量有關,還與各類激素之間的相互調節有關,特別是促進生長的激素與抑制生長的激素之間的比例及平衡有關[26]。本研究將湖北梣種子的IAA、GA3、BR、JA-Me、ZR測定值與ABA含量相比較,如圖6所示,隨著試驗時間的推進,對照GA3/ABA、BR/ABA、ZR/ABA呈逐漸上升的趨勢,IAA/ABA、JA-Me/ABA呈倒“V”形變化;赤霉素處理IAA/ABA、ZR/ABA、BR/ABA、JA-Me/ABA呈倒“V”形變化,GA3/ABA呈逐漸上升的趨勢。各處理的ZR/ABA與GA3/ABA及IAA/ABA與JA-Me/ABA變化趨勢基本相似,其比值在整個試驗期間以GA3/ABA變化最為明顯。從圖5可以看出,IAA/ABA與GA3/ABA、ZR/ABA分別在0.01和0.001水平上呈顯著正相關,相關系數分別為0.59、0.72;GA3/ABA與ZR/ABA在0.001水平上呈顯著正相關(r=0.82)。
3 討論與小結
湖北梣種子的萌發受到多重因素的綜合影響,外源赤霉素處理湖北梣種子可以顯著提高發芽率。種子萌發是物質和能量轉化的過程,膜系統調節著各種物質的運輸及各種酶的活性[27,28]。種子的抗逆性使得60 d可溶性蛋白質含量低于0 d,隨著萌發時間的推進,細胞膜功能受損,種子通過消耗內部滲透物質維持其生理機能和萌發,致使60 d時各處理種子滲透調節物質含量低于0 d,此結果與魏朝陽[29]、楊萬霞[30]的研究結論一致。SOD、POD、CAT能清除逆境條件下種子產生的活性氧,保護膜質系統[31]。在種子萌發的過程中,SOD活性表現出先下降后上升的趨勢。在萌發初期(0 d),各處理組的SOD活性都較低。隨著時間的推移,SOD活性逐漸上升,且在60 d時達到最高水平。赤霉素處理組的SOD活性在萌發30 d和60 d時均高于對照組。赤霉素處理后,種子中POD活性呈持續上升的趨勢。在30 d和60 d時,赤霉素處理組的POD活性顯著高于對照組,表明赤霉素處理能夠顯著提高種子中POD的活性,降低過氧化物對種子細胞膜的傷害,從而增強種子對逆境的抵抗能力,保持種子活性[32]。赤霉素處理后,種子中CAT活性的變化呈先降低后上升的趨勢。在60 d時,赤霉素處理組的CAT活性顯著高于對照組,表明赤霉素處理能夠促進CAT活性的提高,這可能與種子萌發過程中細胞代謝的強度和逆境應對能力有關。綜上所述,赤霉素處理對湖北梣種子萌發過程中SOD、POD和CAT活性產生了顯著影響,表明赤霉素處理能夠調節這些抗氧化酶的活性,從而有效降低種子在萌發過程中產生的活性氧,促進種子萌芽,與何秀燕等[32]在油樟中的研究相一致。
內源激素對種子萌發調控是一個復雜的生理生化過程[33-35],是各激素間的相互平衡、制約、協同[36]。Nambara等[37]的研究表明赤霉素浸種能增加種子抗逆性,增強保護酶活性,降低萌發內源抑制物含量,加快種子的萌發。本研究中,試驗前赤霉素浸種的種子內源JA-Me含量顯著低于對照,萌發過程中,IAA、ZR、BR含量增加幅度顯著高于對照,對萌發抑制物的拮抗作用使種子內部生理生化反應保持活躍狀態,為種子萌發做充分準備。IAA參與種子休眠的調控,促進GA3的合成[38],但高含量的IAA會抑制種子萌發。種子萌發時較高的IAA含量能提高代謝水平[39],但也使種子活力加速衰弱,類似研究在楊立學等[40]對紫椴的研究中也有所報道。Li等[36]對水稻種子的研究表明,BR參與反饋調節并抑制ABA產生,其含量與種子營養含量成正比,能提高發芽率和發芽勢,縮短萌發時間。試驗初期IAA、ZR、GA3與ABA較低的比值說明種子內較高的ABA可能是種子休眠的主要原因,GA3/ABA變化最為顯著,綜合表明IAA、ZR、GA3在調節種子萌發過程中存在一定的促進作用,可能是通過GA3與ABA的拮抗作用或各激素間互相拮抗與促進來調控。ABA不是惟一控制種子休眠與萌發的內源激素[41],ABA與GA3的關系可能為拮抗或者與兩者絕對含量升降無關,種子萌發時ABA含量較高,而試驗過程中ABA增長幅度顯著低于GA3,減輕了對GA3的抑制作用,縮短了種子萌發時間。蘇海蘭等[42]的研究表明出苗前ABA含量持續上升,可能是為了阻止胚的過早萌發和淀粉萌發物的水解,該結果在浦梅等[43]的研究中得到了證實。與“三因子學說”一致,本研究中,較高的GA3與ABA比值水平表明,ABA并未影響GA3的合成或分解,萌發中ZR含量的增加可能解除了ABA對GA3的抑制作用,類似發現在矮金蓮花種子[44]中得以體現。赤霉素浸種后,可能是通過改變種子透性,提高種子的呼吸作用,促使種子與外界氣體交換,以解決生理后熟現象,同時調節種子內部萌發抑制物和促進物的相對水平,類似研究在二喬玉蘭種子上得以運用[45]。
綜上所述,用赤霉素浸種可以顯著提高湖北梣種子發芽率,降低種子萌發過程中電導率;可溶性蛋白質含量呈下降趨勢,各時期均顯著高于對照;SOD活性、POD活性、CAT活性呈上升趨勢,60 d時均顯著高于對照,表明處理后的種子平衡保護酶活性系統較強,質膜受損延緩,抗逆能力較好。種子內源JA-Me含量均顯著低于對照,更有利于內源GA3、ZR含量的增加, 較高的JA-Me含量對種子萌發的啟動和休眠的解除產生了抑制作用,BR、ZR、GA3含量的增加對萌發的啟動具有一定的正調控作用。相關性分析表明,IAA/ABA與GA3/ABA和ZR/ABA分別在0.01和0.001水平上呈顯著正相關;GA3/ABA與ZR/ABA在0.001水平上呈顯著正相關,IAA、ZR、GA3在調節種子萌發過程中存在一定的促進作用,可能是GA3與ABA的拮抗作用或各激素間互相拮抗與促進共同調節控制。
參考文獻:
[1] 張 建.對節白蠟研究概況及展望[J].湖北林業科技,2015,??? 44(2):27-29.
[2] 楊藝濤,袁龍義,向君妍.鄉土樹種對節白蠟在城市景觀空間設計中的應用[J].世界林業研究,2023,36(3): 147.
[3] 鄭鵬麗,程建如,周明芹.瀕危植物對節白蠟遺傳多樣性的cpSSR分析[J].西南農業學報,2020,33(7): 1399-1403.
[4] 孔令雷,胡金鳳,陳乃宏.香豆素類化合物藥理和毒理作用的研究進展[J].中國藥理學通報,2012, 28(2):165-168.
[5] 劉雅琦,張 建,李 剛,等.對節白蠟種子發芽特性的研究[J].種子,2016,35(10):27-31.
[6] 楊湘虹,趙榮秋,劉樂承.對節白蠟的應用與繁殖研究進展[J].長江大學學報(自然科學版),2016,13 (27):4-7.
[7] GUAN Y R,XUE J Q,XUE Y Q,et al. Effect of exogenous GA3 on flowering quality, endogenous hormones, and hormone- and flowering-associated gene expression in forcing-cultured tree peony (Paeonia suffruticosa)[J]. Journal of integrative agriculture,2019,?????? 18(6):1295-1311.
[8] MA H Y,ZHAO D D,NING Q R,et al. A multi-year beneficial effect of seed priming with gibberellic Acid-3 (GA3) on plant growth and production in a perennial grass, Leymus chinensis[J].Scientific reports,2018,8(1):13214.
[9] 羅富成,郭軼敏,彭 健,等.外源激素對納羅克非洲狗尾草種子休眠的破除效果[J].草業科學,2015, 32(3):406-412.
[10] 劉文瑜,魏小紅,許可成,等.蒺藜苜蓿種子休眠機制及其破除方法研究[J].草地學報,2015,23(2):358-365.
[11] DOAIGEY A R,AL-WHAAIBI M H,SIDDIQQUI M H,et al. Effect of GA3 and 2,4-D foliar application on the anatomy of date palm (Phoenix dactylifera L.) seedling leaf[J].Saudi journal of biological sciences,2013,20(2):141-7.
[12] SONG Q,CHENG S,CHEN Z,et al. Comparative transcriptome analysis revealing the potential mechanism of seed germination stimulated by exogenous gibberellin in Fraxinus hupehensis[J].BMC Plant Biol,2019,19(1):199.
[13] FENG R W,LEI L,SU J M,et al. Toxicity of different forms of antimony to rice plant: Effects on root exudates, cell wall components, endogenous hormones and antioxidant system[J].Science of the total environment,2020,771:134589.
[14] QU Z M,QI X C,LIU Y L,et al. Interactive effect of irrigation and polymer-coated potassium chloride on tomato production in a greenhouse[J].Agricultural water management,2020,235:106149.
[15] ZHAO X H,YU H Q,WEN J,et al. Response of root morphology, physiology and endogenous hormones in maize (Zea mays L.) to potassium deficiency[J].Journal of integrative agriculture,2016,15(4):785-794.
[16] JIANG H K,SHUI Z W,XU L,et al. Gibberellins modulate shade-induced soybean hypocotyl elongation downstream of the mutual promotion of auxin and brassinosteroids[J].Plant physiology and biochemistry,2020,150:209-221.
[17] SONG S W,LEI Y L,HUANG X M,et al. Crosstalk of cold and gibberellin effects on bolting and flowering in flowering Chinese cabbage[J].Journal of integrative agriculture,2019,18(5):992-1000.
[18] YANG L,LIU D C,HU W,et al. Fruit characteristics and seed anatomy of ‘Majia pomelo pollinated with cobalt-60?gamma-ray-irradiated pollen[J].Scientia horticulturae,2020,267(1):109335.
[19] CHEN Y,SONG X H,GUO Y Y,et al. Stalk length should be considered for storage quality of broccoli heads based on the investigation of endogenous hormones metabolism[J].Scientia horticulturae,2020,267:109338.
[20] 陳愛葵,韓瑞宏,李東洋,等.植物葉片相對電導率測定方法比較研究[J].廣東教育學院學報,2010,30(5): 88-91.
[21] CHENG Y F,LIU Y L,WEI H M,et al. Quantitation of low concentrations of polysorbates 80 in protein formulations by Coomassie brilliant blue[J].Analytical biochemistry,2019,573:67-72.
[22] CHENG C W,CHEN L Y,CHOU C W,et al. Investigations of riboflavin photolysis via coloured light in the nitro blue tetrazolium assay for superoxide dismutase activity[J].Journal of photochemistry and photobiology B: Biology,2015,148:262-267.
[23] 李合生.植物生理生化試驗原理和技術[M].北京:高等教育出版社, 2000.
[24] 李冬林,張亞楠,金雅琴,等.毛梾種子低溫層積過程中內源激素的變化及其與發芽的關系[J].經濟林研究,2014,32(3):36-41.
[25] 李振華.外源生長素和赤霉素信號調控煙草種子休眠與萌發的機理[D].北京:中國農業大學,2017.
[26] KHAN A A. Primary, preventive and permissive roles of hormones in plant systems[J].Botanical review,1975,41:391-420.
[27] LI Q F,ZHOU Y,XIONG M,et al. Gibberellin recovers seed germination in rice with impaired brassinosteroid signalling[J].Plant science,2020,293:110435.
[28] KIM S Y,WARPEHA K M,HUBER S C. The brassinosteroid receptor kinase, BRI1, plays a role in seed germination and the release of dormancy by cold stratification[J].Journal of plant physiology,2019,241:153031.
[29] 魏朝陽.冬青種子休眠機理及解除方法研究[D].南京:南京林業大學,2007.
[30] 楊萬霞.青錢柳種子休眠原因及萌發生理的初步研究[D].南京:南京林業大學,2004.
[31] SHAH K,NAHAKPAM S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars[J].Plant physiology and biochemistry,2012,57:106-113.
[32] 何秀燕,倪婷婷,劉孟婕,等.不同濃度赤霉素對油樟種子生理及萌芽的影響[J].四川林業科技,2023,44(3):110-115.
[33] DOAIGEY A R,AL-WHAIBI M H,SIDDIQUI M H,et al. Effect of GA3 and 2,4-D foliar application on the anatomy of date palm (Phoenix dactylifera L.) seedling leaf[J].Saudi journal of biological sciences,2013,20(2):141-147.
[34] WEN T,DONG L J,WANG L,et al. Changes in root architecture and endogenous hormone levels in two Malus rootstocks under alkali stress[J].Scientia horticulturae,2018,235:198-204.
[35] ZHANG Y J,LI A,LIU X Q,et al. Changes in the morphology of the bud meristem and the levels of endogenous hormones after low-temperature treatment of different Phalaenopsis cultivars[J].South African journal of botany,2019,125:499-504.
[36] LI Q,YANG A,ZHANG W H. Higher endogenous bioactive gibberellins and α-amylase activity confer greater tolerance of rice seed germination to saline-alkaline stress[J].Environmental and experimental botany, 2019,162:357-363.
[37] NAMBARA E,MARION-POLL A. ABA action and interactions in seeds[J].Trends in plant science,2003, 8:213-217.
[38] NOOR A,ZAIF K,Amjad M,et al. Synthetic auxins concentration and application time modulates seed yield and quality of carrot by altering the umbel order[J].Scientia horticulturae,2020,262:109066.
[39] 廖文燕.金錢松種子貯藏過程中的生理生化變化[D].南京:南京林業大學,2011.
[40] 楊立學,王海南,張 琳,等.低溫層積過程中紫椴種子內源激素含量的變化[J].經濟林研究,2012,30(2): 15-18.
[41] 李建軍,連笑雅,王 蘭.忍冬花蕾延遲開花與內源激素調控研究[J].園藝學報,2019,46(7):1399-1408.
[42] 蘇海蘭,周先治,李 希,等.七葉一枝花種子萌發不同階段貯藏物質及淀粉酶活性變化[J].福建農業學報,2017,32(10):1145-1149.
[43] 浦 梅,孫永玉,高成杰,等.滇重樓種子內源激素含量與種胚長度和萌發的關系[J].林業科學研究,2016, 29(2):268-273.
[44] 張雁明.矮金蓮花種子萌發的內源激素調控及其環境響應規律[D].蘭州:蘭州大學,2018.
[45] 沈 超. 二喬玉蘭種子萌發特性及促萌因素解析[D].南京:南京林業大學, 2013.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
