三都澳大黃魚養殖區域浮游植物群落特征及其與環境因子的關系
2024-06-27 00:00:00吳舒英林雄平陳玲珊肖詩琪陳思晨陳潔
南方農業·上旬 2024年4期

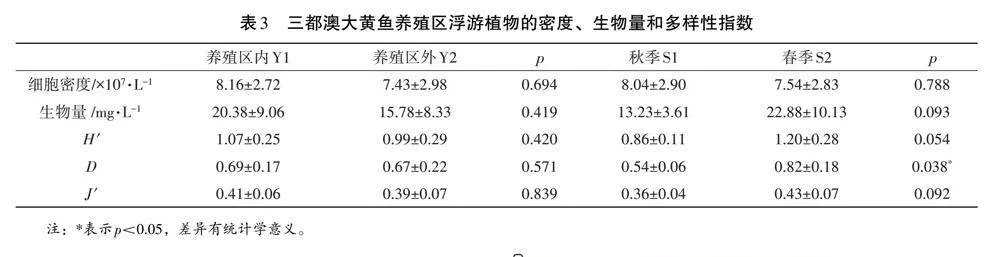


摘 要 為探究三都澳大黃魚養殖區浮游植物群落特征,于 2022 年 11 月、2023 年 4 月對福建省寧德市三都澳大黃魚養殖區浮游植物及水質進行調查。結果表明:三都澳大黃魚養殖區共有浮游植物 6 門、32 種屬,優勢類群為綠藻門和藍藻門。三都澳秋季、春季 6 個位點平均細胞密度為 8.04×107±2.90×107 個·L-1,7.54×107±2.83×107 個·L-1,平均生物量為 13.23±3.61 mg·L-1,22.88±10.13 mg·L-1,養殖區內、外的平均細胞密度為 8.16×107±2.72×107 個·L-1、7.43×107±2.98×107個·L-1,平均生物量為 20.38±9.06 mg·L-1、15.78±8.33 mg·L-1,均不存在顯著性差異(p>0.05)。對浮游植物的細胞密度、生物量、Shannon-Wiener 多樣性指數(H')、Pielou 均勻度指數(J')、豐富度指數(D)進行分析,三都澳大黃魚養殖區內的浮游植物細胞密度、生物量、H'、D、J'均值均高于養殖區外。RDA分析表明總磷是影響三都澳養殖區浮游植物生物量分布的關鍵因子(p<0.05)。相關性分析表明三都澳大黃魚養殖區不同時間段資源差異較明顯,不同生境的浮游植物的資源差異不顯著。
關鍵詞 浮游植物;群落特征;環境因子;三都澳大黃魚養殖區
中圖分類號:Q948.8 文獻標志碼:A DOI:10.19415/j.cnki.1673-890x.2024.07.053
作為海洋最重要的初級生產者,浮游植物對海洋生態、水質評價、海產養殖業發展都有著重要作用[1]。在海洋生態系統層面,浮游植物能通過食物鏈或其他傳遞方式來實現物質循環和能量流動,調節海洋水體的生態穩定。在未被污染的水域中,浮游藻類的種類構成隨季節和環境因素的變化而出現有規律的改變,在已經被污染過的水體中,浮游藻類的種類構成隨污染程度無規律地變化[2]。在海洋養殖業發展層面,浮游植物具有雙重性,一方面能為水產養殖提供所需的生長要素,可作為魚類和其他經濟動物直接或間接的天然餌料;另一方面又可能因過度繁殖而對水體造成污染從而對養殖業造成經濟損失。除此之外,水體環境因子的變化與水體中浮游植物群落特征變化存在相關性,如徐良等通過溫度對藻類生長變化作用的數值進行模擬,證實了高溫對藻類生長變化具有抑制作用[3];顧兆俊等對養殖池塘水體溶解氧調控效果進行的研究表明,溶氧量低的水體底層浮游植物數量較少[4]。中國海域養殖區浮游植物群落結構的研究主要集中于渤海、南海、東海的海灣和島嶼。渤海海域扇貝養殖區浮游植物群落特征變化趨于一致,以硅藻門為主要優勢類群,其浮游植物密度變化呈現出季節差異性,浮游藻類優勢種互有更迭[5-7]。南海海域養殖區養殖方式多樣,養殖區浮游動物群落結構多變,其浮游植物豐度受季節變化較明顯,呈現出夏高冬低現象[8]。東海海域大黃魚養殖區以網箱養殖為主,受不同材質的網箱影響,其浮游植物物種豐富度、多樣性、均勻度及優勢度的月份變化都較明顯。與其他海域一樣,浮游植物細胞豐度、種類分布受季節影響,也以硅藻門藻類為主要浮游植物優勢類群[9]。三都澳大黃魚養殖區因受養漁業發展所造成的客觀因素,水質參數周年變化未形成一致趨勢,且水體富營養化程度較重,而浮游植物群落結構相關調查基本沒有[10-11]。本研究以三都澳大黃魚網箱養殖區為試驗區域,分別于春秋兩季對其進行水質監測及浮游植物群落特征分析,并探索該養殖區浮游植物群落特征與環境因子的相關性,為大黃魚生態養殖提供基礎數據參考。
1 "材料與方法
1.1 "研究區域概況
三都澳又名三沙灣,位于福建省沿海東北部的寧德市境內,地形口小腹大,其海灣出灣口只有唯一的東沖口,寬度僅2.6 km,是世界上少有的海湖[12],其海域面積為714 km2,是中國最大的大黃魚養殖基地,也是世界級天然深水良港,具有避風、水深、水質好等許多優點[10]。
1.2 "水樣采集與樣品處理
1.2.1 水樣采集位點設置
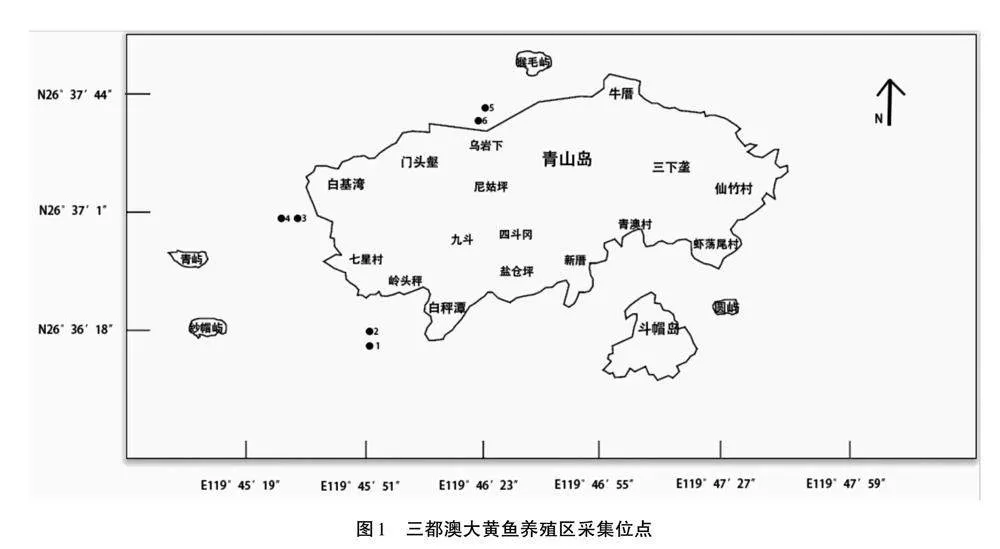
所選采集的地點位于福建省寧德市蕉城區三都澳海域大黃魚養殖區,按照同一大黃魚養殖區分對應的內外共設6個采集位點(圖1),1、3、5為大黃魚養殖池內,2、4、6為大黃魚養殖池外。
1.2.2 "水樣采集
分別于2022年11月13日和2023年4月8日進行秋春季水樣采集。使用采水器采集對應樣點水樣1 L,并滴加10 mL的魯格試劑固定,另外用25號浮游生物網采集到的水樣裝入EP管中,避光保存帶回實驗室。水質參數中的水溫(WT)、pH值、溶氧量(DO)、電導率(Cond.)4項指標通過當天現場利用 YSI 儀器測定得出,透明度(mmg)使用賽氏盤測定。
1.2.3 "樣品處理
將EP管放置于冰箱冷藏并避光保存,用于浮游植物的定性分析;將水瓶里的水靜置48 h后放入虹吸機進行虹吸,將其濃縮后裝入棕色樣品瓶中,避光保存,用于浮游植物的定量計數。
1.3 "指標鑒定
1.3.1 "水體各項指標測定
透明袋剩余水量按照參考文獻[13]用于室內4個理化指標測定,即總磷(TP)、總氮(TN)、硝態氮(N-NO3)、氨氮(N-NH4)。
1.3.2 "浮游生物測定
定性分析:取適量 EP 管里的水,放入顯微鏡(尼康E100)下觀察其特征并拍照記錄,通過比對《中國海域常見浮游硅藻圖譜》等多部藻類分類著作,判斷并識別各個位點中的浮游植物種類。定量計數:在檢測前需上下搖勻,處于無污染的環境下,快速用100 μL的手動移液槍吸取樣液加入浮游生物計數框中。采用視野法,隨機選取25個視野進行浮游植物的細胞計數并進行記錄,其種類鑒定參照《中國海藻志》[14]。
1.4 "數據處理分析
將前期所得數據按照養殖區內、養殖區外、秋季、春季進行分類整理,得到Y1(秋內+春內)、Y2(秋外+春外)、S1(秋總)、S2(春總)。浮游植物密度、生物量、優勢度采用 Excel 公式統計進行處理,用 Origin 9.0 制作浮游植物生物密度與生物量堆積圖[15]。而浮游植物多樣性分析采用H'、D和J'來描述,指數運算公式參照文獻[16-18]。利用軟件SPSS 27.0,分別采用成對樣本t檢驗、獨立樣本t檢驗分析不同季節、不同位點間水質參數的差異性。浮游植物與環境因子的排序分析利用軟件Canoco 5.0采用多元分析方法,最終繪制相關性矩陣圖。
2 "結果與分析
2.1 "水質指標分析
2022年11月13日、2023年4月8日調查三都澳大黃魚養殖區水體理化參數見表1。春秋兩季TP、N-NH4、TN、Cond.存在差異性(p<0.05),DO、WT、mmg、N-NO3差異性極顯著(p<0.01),而pH差異性不顯著。養殖區內外的差異均不顯著。秋季的WT、mmg、N-NO3、N-NH4、TN、Cond.均值均高于春季,而春季的pH、TP、DO均值均高于秋季。養殖區內pH、N-NO3、DO、Cond.均值低于養殖區外,其余指標均高于養殖區。
2.2 "浮游植物定性分析
三都澳大黃魚養殖區春秋兩季調查共檢出浮游植物32種屬(見表2),其中,秋季和春季浮游植物分別為21、27種屬,養殖區內外浮游植物分別為24、26種屬。根據優勢度Y值,優勢種共9種屬,藍藻門4種屬,綠藻門3種屬,硅藻門、隱藻門共2種屬。微囊藻(Microcystis sp.)、小球藻(Chlorella sp.)為春秋兩季6個位點共有的優勢種;弓形藻(Schroederia sp.)、澤絲藻(Limnothrix sp.)為養殖區內的優勢種,棒膠藻(Rhabdogloea sp.)、長孢藻(Dolichospermum sp.)為養殖區外的優勢種。除以上優勢種外,還有小環藻(Cyclotella sp.)、嚙蝕隱藻(Cryptomonaserosa sp.)、卵囊藻(Oocystis sp.)。
2.3 "浮游植物細胞密度、生物量和群落多樣性分析
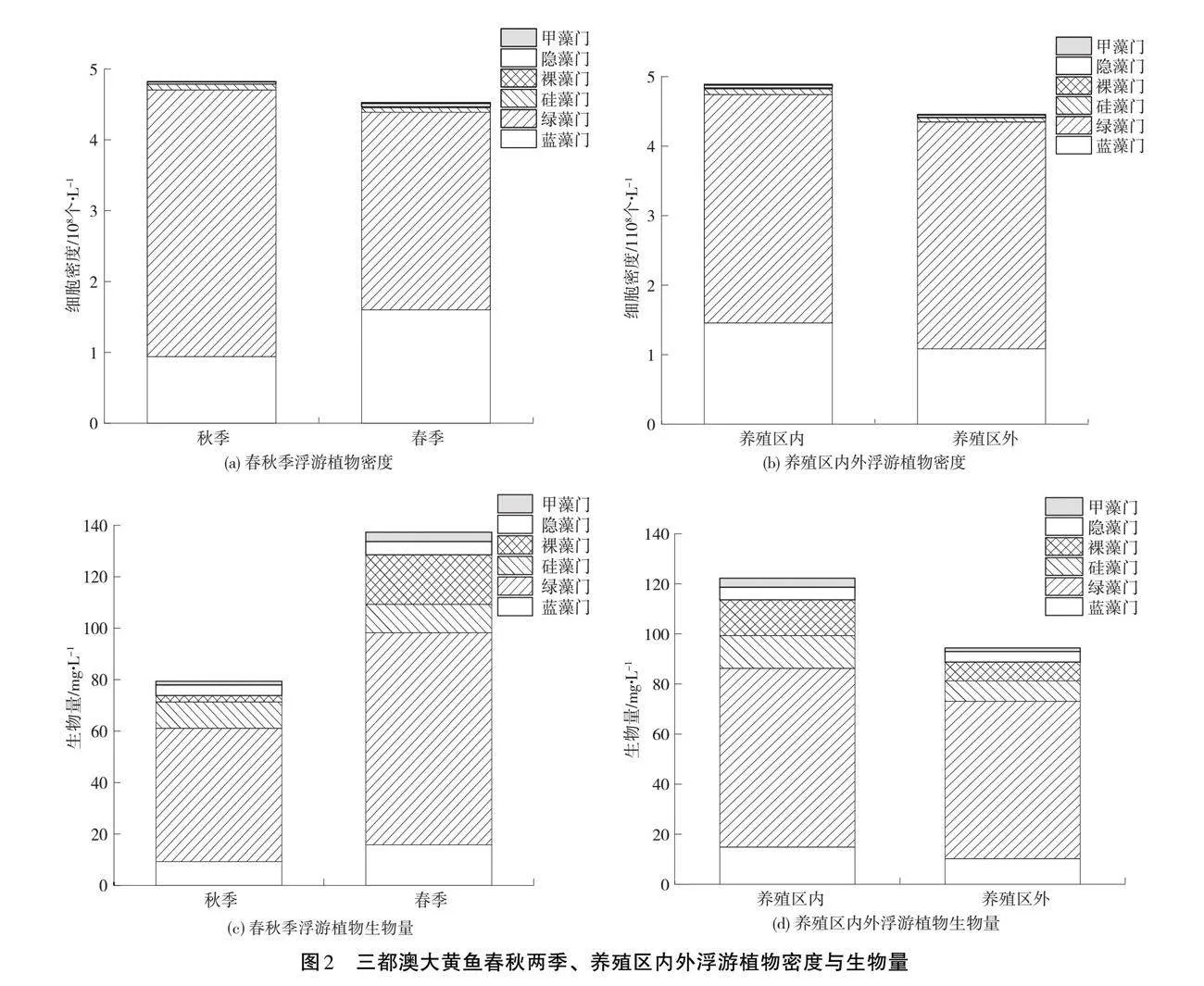
大黃魚養殖區秋春總浮游植物細胞密度分別為4.82×108 個·L-1、4.53×108 個·L-1,總生物量分別為79.37、137.31 mg·L-1,養殖區內外總浮游植物細胞密度分別為4.89×108 個·L-1、4.46×108 個·L-1,總生物量分別為122.26、94.42 mg·L-1(見圖2)。藍藻門、綠藻門為三都澳大黃魚養殖區春秋兩季浮游植物優勢類群,也是養殖區內外浮游植物優勢類群。三都澳大黃魚養殖區總浮游植物細胞密度秋季大于春季,而生物量春季大于秋季。養殖區內的總浮游植物密度和總生物量都大于養殖區外。春秋兩季、養殖區內外浮游植物細胞密度、生物量、H'、J'差異性不大、而春秋兩季的D有顯著性差異(p<0.05),養殖區內外D無明顯差異(表3)。
2.4 "浮游植物群落結構與環境因子的關系
根據三都澳大黃魚養殖區浮游植物密度,選出用于排序的15種浮游植物,用Canoco 5.0進行樣點與環境因子間分析、RDA分析,結果如圖3。RDA分析表明總磷是影響三都澳養殖區浮游植物群落結構相關性較強的因子(p<0.05),養殖區內外浮游植物與環境因子間相關性不強。秋季的浮游植物分布主要與WT、mmg、N-NO3、N-NH4、TN、Cond.呈正相關。春季的浮游植物主要與pH、TP、DO呈正相關。TP、pH和DO分別與第一軸顯著正相關(0.583 8、0.496 5、0.573 8),而TN、N-NH4、N-NO3、WT、Cond.和mmg分別與第一軸顯著負相關(-0.673 3、-0.507 1、-0.591 1、-0.516 3、-0.707 8)(圖3a)。RDA排序圖還表明弓形藻、澤絲藻、嚙蝕隱藻、小球藻、小環藻與N-NH4、TN、N-NO3、WT、Cond、mmg呈正相關,與pH、DO、TP呈負相關。囊裸藻、微囊藻、衣藻、翼膜藻、多甲藻、長孢藻與pH、TP、DO呈正相關,與TN、N-NO3、N-NH4、WT、Cond.、mmg呈負相關。空球藻與TN、N-NO3、WT、Cond.、mmg呈正相關,與DO、TP呈負相關,而棒膠藻與之相反。
3 "討論
3.1 "三都澳大黃魚養殖區春秋水質特征
3.1.1 "水溫特征
三都澳大黃魚養殖區各采集位點水溫范圍為15.9 ~ 22.6 ℃,與測量地氣溫相差不大,符合GB11607—89《漁業水質標準》[19]。三都澳大黃魚養殖區春秋WT差異極顯著(p<0.01),養殖區內、外差異不顯著。這與挑選的采集點有關,基本處于同緯度地帶,海水溫度變化不大[20],而春秋季節溫度差異則與東海黑潮有關[21]。春秋測出的平均水溫值分別為16.63、22.48 ℃,這與余訓凱2023年調查三都澳大黃魚水產養殖區的結果相似[9],但其水溫都低于28 ℃,并非大黃魚生長養殖最佳溫度[22]。目前三都澳大黃魚養殖區以網箱養殖為主,海水溫度主要為自然溫度,人為干涉難以起效果,因此仍建議在夏季培育大黃魚。
3.1.2 "溶氧量特征
參照GB11607—89《漁業水質標準》,春秋兩季、養殖區內外測出平均的DO值均高于5 mg·L-1,遠超Ⅱ類海水水質標準[19]。三都澳大黃魚養殖區春秋DO差異極顯著(p <0.01),養殖區內外差異不顯著。這與處于同時期水樣采集有關,且采集深度差異不大,測得的養殖區內外溶氧量變化不大,而春秋季溶氧量差異則可能是臨近梅雨季、光照條件不一所致[23]。
3.1.3 "pH值和透明度特征
pH值是影響養殖區生物生長的重要理化因子之一,pH值為6.5 ~ 9.0魚類能夠安全生活。本研究中,三都澳大黃魚養殖區春秋兩季pH值變化不大,春季均值7.49,秋季均值6.93,與黃匡南等調查三都澳水產養殖海域水質結果相符[10]。秋季測出水體平均透明度為80.00 cm,與鄭欽華調查三都澳漁業水質結果相符,該階段海水較清澈,有利于大黃魚的培養[24]。
3.1.4 "春秋營養鹽特征
養殖區秋季測出的N-NO3與N-NH4的值均大于春季,不同于余訓凱的調查結果[9],這可能與浮游植物繁殖有關,入春使得浮游植物的繁殖與生長增加[25]。氮、磷元素被認為是造成海區富營養化的主要營養物質[26]。春季測出的總氮平均值高于秋季,養殖區內略高于養殖區外,可認為春季水體營養化高于秋季,養殖區內高于養殖區外,這與陸地負荷有關[27]。2007年福建三都澳漁業水域水環境調查顯示,無機氮已成為澳內主要污染物[24]。三都澳大黃魚養殖區春秋兩季N-NH4、TN存在顯著差異性(p<0.05),N-NO3差異性極顯著(p<0.01),而養殖區內外差異均不顯著,因此也可以認為三都澳大黃魚養殖區不同時間段資源差異較明顯,不同生境浮游植物資源差異不顯著。
3.2 "浮游植物群落特征及其與環境因子的相關性
3.2.1 "浮游植物群落特征
浮游植物的組成和數量在1年內的不同季節有規律地發生變化,秋季和春季養殖區內外生境基本一致,但兩季的優勢物種數量和種類有所不同[28]。三都澳大黃魚養殖區春秋兩季調查共檢出浮游植物32種屬,其中,秋季和春季浮游植物分別為21、27種屬,養殖區內外浮游植物分別為24、26種屬,且綠藻門種屬最多。優勢種均以藍藻門、綠藻門為主。三都澳大黃魚浮游植物春秋、養殖區內外以綠藻門為主,而東海海域以硅藻門為主,這與海域水體中營養物質流動有關[29]。秋春總浮游植物細胞密度測出分別為4.82×108 個·L-1、4.53×108 個·L-1,養殖區內外總浮游植物細胞密度分別為4.89×108 個·L-1、4.46×108 個·L-1。這與2014 年海洋生物多樣性環境監測結果相符,而所測H'、D值則均低于其結果[30]。多樣性指數、均勻度是生物群落特征數值的研究指標。種類豐富程度、種間數量分布均勻狀況、多樣性指數值高低與群落結構穩定性有關[31]。結果說明三都澳大黃魚養殖區浮游植物群落結果穩定性變差。多樣性指數常用來評價水質狀況,本試驗在春秋兩季、養殖區內外的H'、J'差異性不大,僅在春秋兩季浮游植物豐富度指數(D)存在顯著性差異(p<0.05),而養殖區外的植物細胞密度、H'、D、J'低于養殖區內,說明春秋兩季水質情況差異不大,而養殖區外的水質好于養殖區內[32]。
3.2.2 "浮游植物群落特征與環境因子的相關性
WT、pH、營養鹽等環境因子對浮游植物組成結構有較大的影響[33]。春季溫度較低,海水混合度較好,營養鹽豐富,為浮游植物生長提供了適宜的環境,因此春季綠藻門、藍藻門、裸藻門的生物量密度大于秋季[34]。經過鑒定分析,此次調查浮游植物綠藻門的數量最多,其中,小球藻為綠藻門優勢種,微囊藻為藍藻門優勢種,N-NO3、N-NH4、TN、電導率、透明度與小球藻的生長繁殖呈正相關,pH、DO、TP與微囊藻的生長繁殖呈正相關。營養鹽也是三都澳水體中重要的理化因子,三都澳養殖區的生物在養殖過程中產生的糞便可使營養鹽含量增加,促進浮游植物的生長繁殖,春季的N-NO3、N-NH4、TN低于秋季,養殖區內TP、N-NH4、TN高于養殖區外,因此春季的生物量密度高于秋季,養殖區內高于養殖區外[35]。根據營養鹽的相對限制法則,磷被優先利用的可能極大[36]。本研究也表明總磷是影響三都澳養殖區浮游植物生物量分布的關鍵因子(p<0.05)。
參考文獻:
[1] 王慧,蔡潤林,甘永亮,等.海洋浮游植物和藻際細菌的相互作用關系及其在海洋生態系統中的重要作用[J]. 汕頭大學學報(自然科學版),2022,37(3):3-21.
[2] 陳長平,高亞輝,林鵬. 深圳福田紅樹林保護區浮游植物群落的季節變化及其生態學研究[J]. 廈門大學學報(自然科學版),2005(S1):11-15.
[3] 徐良,馮平,孫冬梅,等. 水溫對藻類生長變化影響的數值模擬[J]. 安全與環境學報,2013,13(5):76-81.
[4] 顧兆俊,劉興國,吳娟,等. 養殖池塘水體溶解氧調控效果研究[J]. 水產科技情報,2009,36(6):297-299.
[5] 楊俊麗,李希磊,于瀟,等. 2016年萊州灣扇貝養殖區浮游植物群落生態特征[J]. 中國海洋大學學報(自然科學版),2019,49(8):27-36.
[6] 卜世勛. "河北撫寧海灣扇貝養殖區營養鹽及浮游植物變化特征及分析[D]. 天津:天津農學院,2023.
[7] 盧鈺博,于瀟,李希磊,等. 養馬島扇貝養殖區浮游藻類的生態特征及其與環境因子的關系[J]. 漁業研究,2019,41(3):207-216.
[8] 蘇家齊,朱長波,張博,等. 海陵灣網箱養殖區浮游植物群落結構及其與環境因子的關系[J]. 上海海洋大學學報,2022,31(4):929-939.
[9] 余訓凱. 三都澳水產養殖區水質調查[J]. 水產養殖,2023,44(2):29-32.
[10] 黃匡南,陳彩珍,陳佳,等. 三都澳水產養殖海域水質情況分析[J]. 河北漁業,2019(7):30-33,60.
[11] 吳衛平. 三都澳海域水質變化情況調查與評價[J]. 化學工程與裝備,2019(8):322-326.
[12] 雷廷波. 三都澳海域無機氮污染的分析及建議[J]. 皮革制作與環保科技,2020,1(Z1):28-33.
[13] 國家環境保護總局水和廢水監測分析方法編委會. 水和廢水監測分析方法[M]. 北京:中國環境科學出版社,2002.
[14] 中國孢子植物志編委會. "中國海藻志[M]. "北京: 科學出版社,2013.
[15] 劉海平,葉少文,楊雪峰,等. 西藏尼洋河水生生物群落時空動態及與環境因子的關系:1.浮游植物[J]. 湖泊科學,2013,25(5):695-706.
[16] SHANNON E E,WEAVER W. The mathematical theory of communication[M]. London: University Illinois Press,1949:125.
[17] PIELOU E C. Ecological diversity[M].New York: John Wiley,1975:165.
[18] 馬永紅,曾燏,任麗萍,等. 嘉陵江四川段藻類植物群落結構及水質評價[J]. 應用生態學報,2012,23(9):2573-2579.
[19] 中華人民共和國國家標準——漁業水質標準(GB11607—89)[J]. 環境保護,1989(12):25-27.
[20] 郭軍凱. 海水的溫度與鹽度[J]. 試題與研究,2011(27):41-42.
[21] 周立佳,黨振中,董慧超,等. 基于ROMS模式的東海黑潮季節變化特征模擬研究[J]. 艦船電子工程,2016,36(7):91-94,153.
[22] 阮成旭,吳德峰,袁重桂. 溫度對大黃魚(Pseudosciaena crocea)幼魚生長和養殖水質的影響[J]. 廣州大學學報(自然科學版),2013,12(2):36-39.
[23] 阮松樹. 夏季水體缺氧的原因[J]. 農家科技,2011(6):38.
[24] 鄭欽華. 福建三都澳漁業水域水環境監測與評價[J]. 寧德師專學報(自然科學版),2010,22(3):250-254,267.
[25] 韋欽勝,戰閏,臧家業,等. 南黃海春季海水化學要素的分布特征及其受控因素[J]. 海洋科學,2010,34(3):52-60.
[26] 鄭欽華,劉家富,陳洪清,等. 三都灣海水中無機氮和無機磷的分布特征及海水質量的評價[J]. 福建水產,2002(4):22-27.
[27] 樸香花. 大連灣海域氨氮、磷酸鹽及藻類生長的數值模擬研究[D]. 大連:大連理工大學,2006.
[28] 鄒雙燕. 平潭近岸海域春、夏季浮游植物特征及其與環境因子相關性分析[J]. 漁業研究,2022,44(3):266-274.
[29] 賴俊翔. "長江口及鄰近海域浮游植物色素與富營養化研究[D]. 青島:中國科學院研究生院(海洋研究所),2011.
[30] 謝宏英. "赤潮災害風險評估研究[D]. 上海:上海海洋大學,2018.
[31] 李冀剛,楊振雄,葛頌,等. 汕頭韓江榕江河口近岸海域浮游植物群落結構與環境因子的關系[J]. 海洋湖沼通報,2023,45(2):133-141.
[32] 朱為菊,龐婉婷,尤慶敏,等. 淮河流域春季浮游植物群落結構特征及其水質評價[J]. 湖泊科學,2017,29(3):637-645.
[33] REYNOLDS C S. "The ecology of phytoplankton [M]. "Cambridge: Cambridge University Press,2006.
[34] 江杉,王益澄,馬仁鋒. 中國東海葉綠素濃度變化分析及其海水溫度響應[J]. 測繪通報,2020(6):39-44.
[35] 林永添. 三都灣大黃魚網箱養殖區海水營養鹽狀況的初步研究[J]. 福建水產,2013,35(3):211-217.
[36] 楊東方,張經,陳豫等. 營養鹽限制的唯一性因子探究[J]. 海洋科學,2001(12):49-51.
(責任編輯:易 "婧)
