燈盞花3-O-葡萄糖基轉移酶基因的克隆、表達及生物信息分析
2024-09-03 00:00:00莫維淼王夢欽徐達何鳳明陳曉波張云峰
南方農業·上旬 2024年7期
關鍵詞:黃酮


摘 要 從燈盞花cDNA中克隆出燈盞花3-O-葡萄糖基轉移酶基因(EbUF3GT)并對其序列進行生物信息學分析以預測其功能。根據課題組前期的轉錄組分析數據,篩選出了燈盞花EbUF3GT基因的部分序列,并利用RACE基因克隆方法,獲得燈盞花EbUF3GT基因的全長cDNA,運用生物信息學分析該基因的相似性和同源性、編碼蛋白的理化性質,并通過熒光定量PCR檢測基因在植物不同部位中的表達情況。結果發現,克隆cDNA的ORF長1 395 bp,編碼464個氨基酸,其蛋白分子量51 850.48,理論等電點為5.48。EbUF3GT蛋白由24.78%的α-螺旋(Alpha helix)、22.20%的延伸(Extendedstand)、53.02%的隨機卷曲(Random coil)組成,屬于GT家族中的B類型且包含PSPG盒的保守序列。燈盞花的EbUF3GT基因與青蒿、艾草親緣關系較近,屬同一個分支,它們的UF3GT具有共同起源。EbUF3GT基因在根、莖、葉、花這幾個部位都能表達,尤其在花中的表達量最高,干旱脅迫也會增加它的表達。結論:通過對EbUF3GT研究,為進一步探索EbUF3GT基因在燈盞花類黃酮次生代謝產物合成和調控機制提供了重要的理論依據。
關鍵詞 燈盞花;糖基轉移酶;黃酮;基因克隆;生物信息學
中圖分類號:Q785 文獻標志碼:A DOI:10.19415/j.cnki.1673-890x.2024.13.001
燈盞花[Erigeron breviscapus(Vant.)Hand-Mazz.]又名短亭飛蓬,為菊科(Compositae)飛蓬屬(Erigeron)多年生植物,主要分布在我國西南地區,是云南省最具發展潛力的藥用植物之一,具有通經、活血、止痛的藥用功效[1]。燈盞花的野生資源和主要種植基地集中在云南紅河、大理等地[2],目前人工種植已成為燈盞花藥材的主要來源,但有效成分高、抗性強、適應范圍廣的優良品種選育一直是燈盞花產業發展的瓶頸[2-4]。因此,基于分子機制研究燈盞花關鍵活性成分的生物合成、生物及非生物抗性機制,對培育優良燈盞花品種尤為迫切。
燈盞花富含黃酮[5]。黃酮具有廣泛的生物活性和健康益處,是一種重要的天然營養素。糖基化修飾是一種常見的生物化學修飾過程,特別是在植物的次生代謝產物中。這一修飾過程通常由UDP-糖基轉移酶(UDP-glycosyltransferase,UGT)催化完成。UGT能夠將糖基從UDP-糖分子上轉移,并連接到不同的底物分子上,從而形成糖基化產物。類黃酮化合物的糖基化反應通常發生在相對容易接受糖基化修飾的羥基位置,如3-OH、5-OH和7-OH。其中UF3GT在類黃酮化合物的糖基化反應中發揮著重要作用,它催化將葡萄糖從尿苷二磷酸(UDP)-葡萄糖分子轉移至花青素或類黃酮的3-OH位置上,形成結構穩定的花色苷和類黃酮3-O-糖苷類化合物(見圖1)[6-7]。由此,苷類化合物可穩定存在并運輸到液泡中貯存,為后續修飾反應提供先決條件,在類黃酮生物合成途徑中發揮著重要作用[8-9]。目前,3GT作為催化類黃酮化合物最重要的修飾糖基化的酶,在植物研究中受到廣泛關注。例如,研究者通過基因克隆和蛋白質工程在甘藍中得到以UDP-葡萄糖,促進生成飛燕草素3-O-葡萄糖苷,從而增加黃酮含量[10];在甘薯中完成3GT基因的克隆,驗證其與其他植物的同源性,通過其的過表達可增加黃酮含量[11]。
盡管目前很多學者都對燈盞花的藥理活性及其活性物質合成途徑進行了研究,但燈盞花中類黃酮合成途徑合成的關鍵糖基轉移酶——3-O-葡萄糖基轉移酶(3GT)的研究還鮮有報導。為此,本研究根據課題組前期的轉錄組分析數據,篩選出了燈盞花3-O-葡萄糖基轉移酶基因(EbUF3GT)的部分序列,利用RACE基因克隆,獲得了燈盞花EbUF3GT基因全長的cDNA,再利用生物信息學對其序列進行分析以預測功能。實驗結果對后續培育高含量黃酮成分的燈盞花品種具有一定參考價值。
1" 材料與方法
1.1" 實驗材料
研究所用燈盞花種植于教育部可持續能源開發利用工程中心。新鮮葉片用于總RNA提取。總RNA用于合成cDNA第一鏈。
pMD18-T載體購自大連寶生物公司,大腸桿菌DH5α感受態細胞購買于北京Trans Gene公司。
1.2" 實驗方法
1.2.1" 擴增cDNA
克隆燈盞花EbUF3GT的cDNA部分片段。回收目的片段(膠回收試劑盒 TIANgel Midi Purification Kit 購自天根公司),轉化大腸桿菌DH5a,菌液PCR驗證陽性克隆,純化后的PCR產物送至上海生工直接測序。
1.2.2" EbUF3GT 3′-RACE和5′-RACE擴增
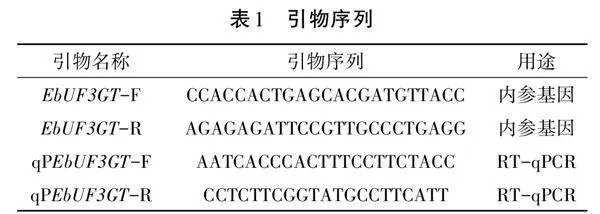
設計引物EbUF3GT基因的特異性引物A235-1(GSP1)F:AGGCACGTCGCGGTTTTCG;R:TCAAAG GGTGGCACTAGTG、A235-2 (GSP2)5′-ATCAGAGAA TAGGCCTTGGT-3′、A235-3 (GSP3)5′-TTGCAGTGCC AAAGAATGAA-3′(見表1)。
采用Invitrogen公司的5′RACE System for Rapid Amplification of cDNA Ends,Version 2.0試劑盒獲得該基因的5′端序列。操作過程中使用到的EP管和槍頭需經0.1% DECP水浸泡過夜處理,并嚴格按照試劑盒說明進行操作。
1.2.3" 生物信息學分析
對EbUF3GT基因cDNA全長進行生物信息學分析,根據信息預測基因的結構和功能,利用NCBI查找該基因的ORF序列并推測其氨基酸序列,采用NCBI Blastp蛋白質數據庫收集EbUF3GT的同源蛋白序列,利用DNAMAN和Jalview軟件對EbUF3GT同源氨基酸序列進行多重比對,通過MEGA軟件,采用鄰接法繪制EbUF3GT系統發育樹。利用ProtScale預測蛋白質的疏水性/親水性,通過Expasy的ProtParam tool推測蛋白質分子量及等電點。
1.2.4" 燈盞花的黃酮含量測定
燈盞花黃酮含量測定參照王夢欽[12]的方法,使用同一時期正常澆水的燈盞花根、莖、葉、花和干旱脅迫后燈盞花的葉片作為實驗材料,進行黃酮含量測定。
1.2.5" EbUF3GT基因qRT-PCR分析
將燈盞花根、莖、葉、花采集后分別放入液氮中,并在液氮中研磨樣品,用Trizol法提取總RNA。利用實時熒光定量PCR儀對EbUF3GT進行表達分析,生物學重復3次,技術重復3次。
2" 結果與分析
2.1" EbUF3GT的克隆及載體構建
根據已有的燈盞花轉錄組數據庫,設計EbUF3GT特異性引物,利用末端克隆技術分別擴增獲得EbUF3GT基因的5′和3′末端序列分別為259 bp、270 bp(見圖2 B-C),將EbUF3GT基因與pMD 18-T克隆載體連接,驗證后測序,利用Blast進行同源序列比對,確定擴增條帶為目的基因。
2.2" 燈盞花EbUF3GT的核酸序列信息分析
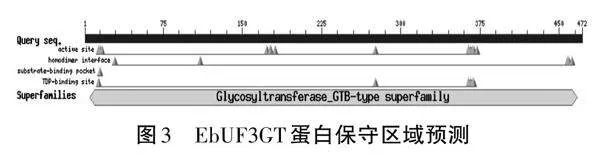
將克隆的EbUF3GT進行測序,獲得序列用ORF Finder查找EbUF3GT的最大開放閱讀框(ORF)。發現克隆cDNA的ORF長1 395 bp,編碼464個氨基酸。利用ExPASy數據庫ProtParam軟件在線分析的結果顯示,其蛋白分子量51 850.48,理論等電點為5.48。在NCBI網站上進行功能區域分析,結果表明EbUF3GT包含糖基轉移酶-GTB-超型家族的結構域,表明所克隆的基因屬于GT家族中的B類型(見圖3)。
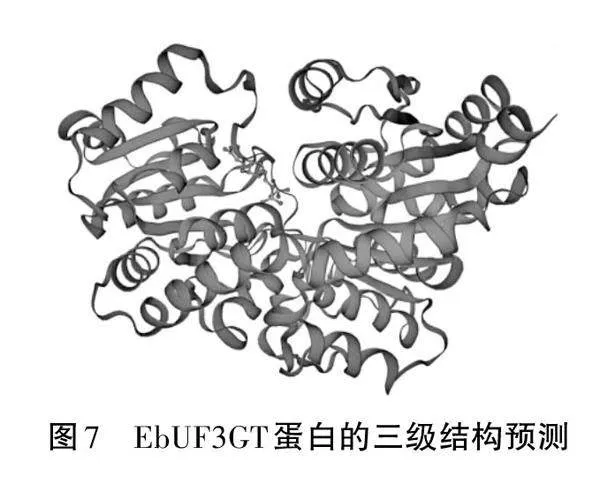
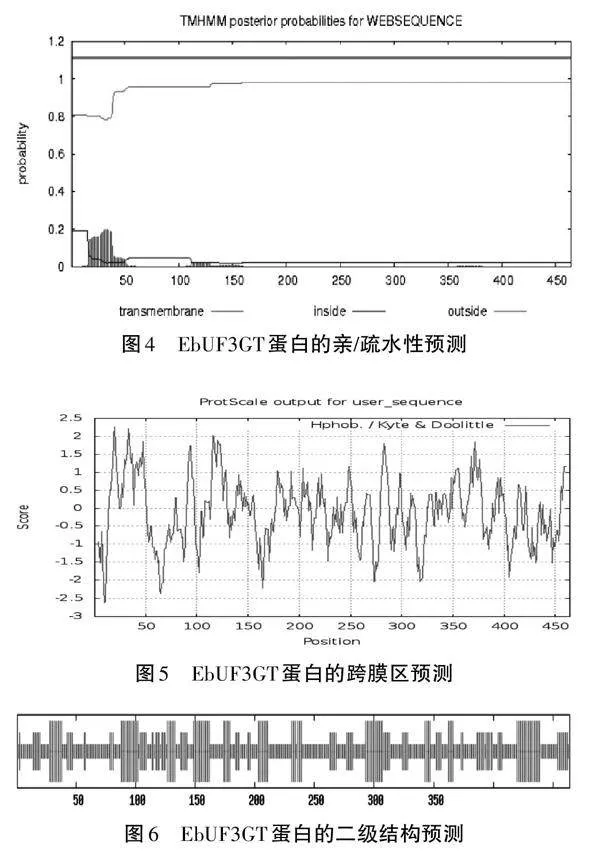
依據ProtScale疏水性/親水性的規律,預測結果表明在多肽鏈的第20位氨基酸最高分值為2.2,疏水性最強(見圖4)。在多肽鏈的第10位氨基酸最低分值為-2.6,親水性最強,于是可以推測此蛋白為親水性蛋白。TMHMM分析結果顯示基因EbUF3GT蛋白跨膜結構域分析,由圖可知該相關基因有半跨膜結構域,因此有半跨膜區,說明EbUF3GT的蛋白質是與細胞信號轉導有關的膜受體蛋白(見圖5)。利用GORIV程序對EbUF3GT基因的二級結構進行預測,結果表明EbUF3GT蛋白由24.78%的α-螺旋(Alpha helix )、22.20%的延伸(Extendedstand)、53.02%的隨機卷曲(Random coil)組成(見圖6)。利用SWISS-MODEL軟件用2acv為模板對EbUF3GT蛋白序列進行同源模建,獲得其三維結構(見圖7)。EbUF3GT蛋白的預測三維結構與2acv為模板空間結構類似,同源性為81.75%。EbUF3GT蛋白預測三維結構的分辨率:2.00 ?,Ramachandran Favoured數值為94.21%,表明這個三級結構是可靠的。
2.3" 燈盞花EbUF3GT蛋白序列信息分析
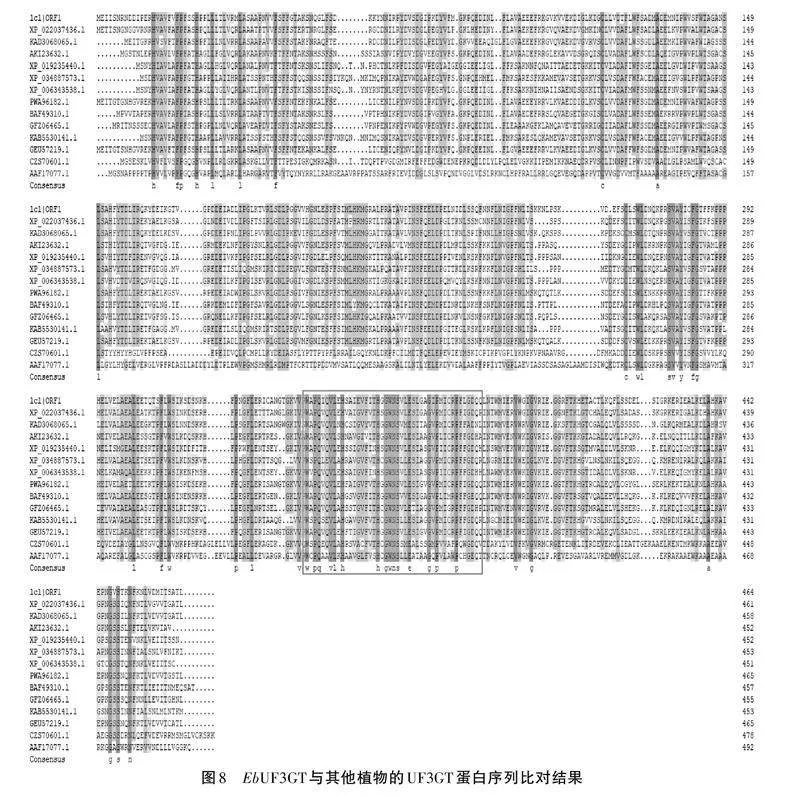
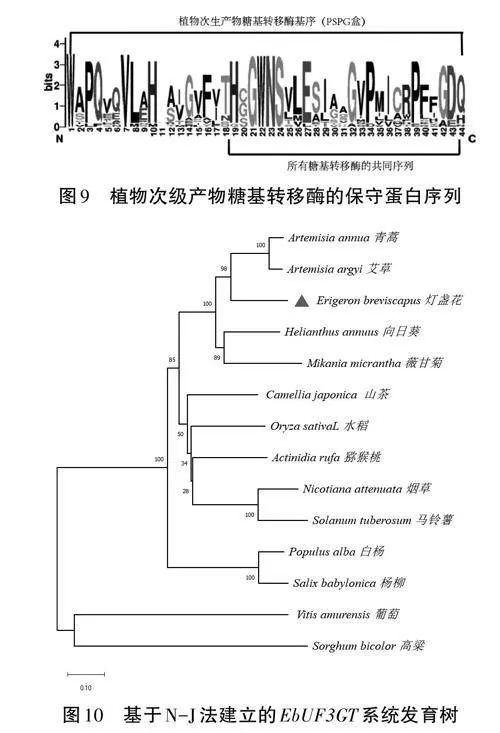
將EbUF3GT氨基酸序列與NCBI蛋白質數據庫進行Blastp比對,發現該EbUF3GT與NCBI數據庫中其他植物的基因同源性很高,與向日葵(登錄號:KAD30680651)、薇甘菊(登錄號:AK123632.1)、煙草(登錄號:XP-0192354401)、白楊(登錄號:XP-03487573)、馬鈴薯(登錄號:XP-00634358.1)、青蒿(登錄號:PWA96182.1)、山茶(登錄號:BAF49310.1)、獼猴桃(登錄號:GF206465.1)、柳樹(登錄號:KAB5530141.1)、艾草(登錄號:GEU57219.1)等親緣較近,與葡萄(登錄號:CZS70601.1)、高粱(登錄號:AAF17077.1)等親緣關系較遠,通過DNAMAN和Jalview軟件對該EbUF3GT基因與以上12個UF3GT基因的蛋白質序列進行多重比較(見圖8)。通過多重序列比對發現EbUF3GT蛋白含有糖基轉移酶中的一個高度保守序列PSPG盒。PSPG對結合糖基供體有關鍵作用(見圖9)。再利用MEGA對以上的同源性比對結果繪制系統發育樹,結果表明燈盞花的EbUF3GT基因與同為菊科的青蒿、艾草親緣關系較近,屬同一個分支,表明EbUF3GT在菊科中具有一定的保守型(見圖10)。
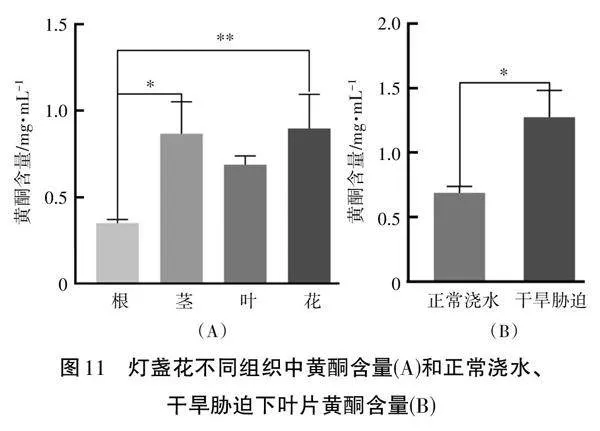
2.4" 燈盞花不同部位和不同處理的黃酮含量測定
對燈盞花不同組織中黃酮含量的測定表明,黃酮含量在花中的含量最高(0.95±0.11 mg·mL-1),葉和莖中黃酮含量分別為0.65±0.1 mg·mL-1和0.75±0.1 mg·mL-1,根中黃酮含量最少(見圖11-A)。將干旱脅迫和正常澆水處理的葉片相比較,發現干旱脅迫會使黃酮含量上升(見圖11-B) 。
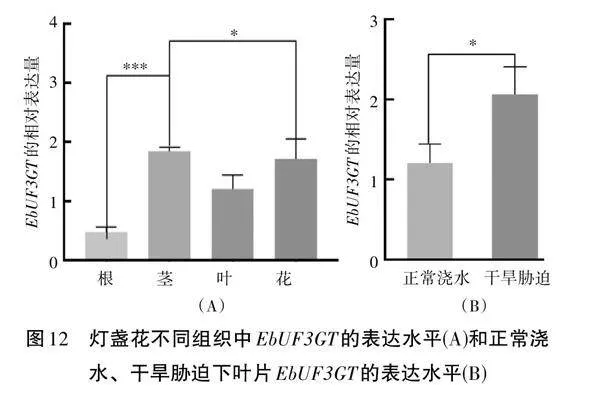
2.5" 燈盞花不同部位和不同處理中EbUF3GT基因表達水平
通過qRT-PCR分析燈盞花不同組織中EbUF3GT基因的表達水平,EbUF3GT在不同的組織中顯示出明顯的表達差異(見圖12),尤其在花中的表達量最高,根的表達量最小。干旱處理后葉片中EbUF3GT的表達量也顯著上升。上述結果表明不同組織和不同干旱處理中的黃酮含量變化一致,表明EbUF3GT在類黃酮的合成中具有重要作用。
3" 討論與結論
3.1" 討論
燈盞花作為云南歷史悠久的民間藥物,目前在云南已有較為完善的種植體系和臨床治療藥物研發體系,但品質改善和藥物創新技術依舊薄弱[12]。燈盞花產業面臨著野生資源不斷減少而人工栽培種存在自交衰退等問題,在藥效方面制劑適應癥范圍狹窄,也嚴重限制了燈盞花產業發展[2]。UF3GT是植物類黃酮-3-O-葡萄糖苷化合物生物合成途徑的關鍵酶之一[8,13],目前已有很多學者對類黃酮-3-O-糖基轉移酶進行深入研究,為挖掘類黃酮生物合成途徑和功能奠定了基礎[14-16]。隨著分子生物學和生物信息學的發展,燈盞花活性物質基因的研究得到了越來越多的關注,也為改良其品質提供了新的思路。
本文從生物信息學角度,對不同植物來源的13個UF3GT基因的核酸及蛋白質序列的組成、理化性質、結構特點、蛋白質保守功能域及系統進化關系等進行預測和分析發現,EbUF3GT的ORF全長1 395 bp,編碼464個氨基酸,其蛋白分子量51 850.48,理論等電點為5.48,大部分蛋白呈親水性,均存在明顯的疏水區和親水區。EbUF3GT基因有半跨膜結構域,因此有半跨膜區,說明EbUF3GT的蛋白質是與細胞信號轉導有關的膜受體蛋白。二級結構的預測表明EbUF3GT蛋白由24.78%的α-螺旋、22.20%的延伸鏈、53.02%的隨機卷曲組成。在氨基酸序列多重比對和系統進化樹構建分析過程中,多數蛋白進化過程中具有保守性和多樣性,不同物種間的進化可能存在一種定向趨勢,導致在某些分支上發生共同的變化,有物種內獨特的平行進化同源基因。
EbUF3GT在花的表達量最高,根的表達量最小。干旱處理后葉片中EbUF3GT的表達量也顯著上升。上述結果表明不同組織和不同處理中的黃酮含量變化一致。我們推測EbUF3GT在類黃酮合成過程中起著關鍵的轉化作用,參與了這些活性成分的糖基化過程。
3.2" 結論
本實驗為燈盞花3-O-糖基轉移酶的功能解析和生物合成研究提供理論基礎。通過生物信息學分析可望為本實驗室后期對燈盞花EbUF3GT在原核中的異源表達及黃烷酮糖的生物合成提供理論基礎。本研究為今后采用生物技術提高藥用植物燈盞花的黃酮含量,促進燈盞花高品質產業化生產提供了重要的理論依據。
參考文獻:
[1] 王澤華,丁美玲,張俊敏,等.云南燈盞花產業的潛力礎與發展路徑[J].社會主義論壇,2022(8):30-31.
[2] 蘇燦,普元柱,高勇,等.云南省燈盞花產業發展現狀及對策[J/OL].中藥材,2023(5):1067-1074.
[3]" 楊春德.燈盞花產業發展現狀及對策措施[J].云南農業,2014(2):56-57.
[4]" 紅河千山生物工程有限公司.以科技為依托打造燈盞花產業[J].云南科技管理,2009,22(2):53.
[5]" LIU R J, SONG J L, LIU S Q, et al. Genome-wide identification of the Capsicum bHLH transcription factor family: discovery of a candidate regulator involved in the regulation of species-specific bioactive metabolites[J]. BMC Plant Biology, 2021,21(1): 262.
[6]" GAO Q Q, SONG W L, LI X, et al. Genome-wide identification of bHLH transcription factors: discovery of a candidate regulator related to flavonoid biosynthesis in Erigeron breviscapus[J]. Frontiers in Plant Science, 2022, 13: 977649.
[7] 張廣輝,李翠婷,王建軍,等.燈盞花黃酮合成酶Ⅱ基因克隆及其生物信息學分析[J].中國中藥雜志,2013,38(14):2231-2236.
[8] LIANG W X, NI L, CARBALLAR-LEJARAZú R, et al. Comparative transcriptome among Euscaphis konishii Hayata tissues and analysis of genes involved in flavonoid biosynthesis and accumulation[J]. BMC Genomics, 2019, 20(1):24-37.
[9]" ZHAO Y, ZHANG G H, TANG Q Y, et al. EbMYBP1, a R2R3-MYB transcription factor, promotes flavonoid biosynthesis in Erigeron breviscapus[J]. Frontiers in Plant Science, 2022, 13: 946827.
[10]" 胡燈科.甘藍U-box家族全基因組鑒定及BoPUB8基因的克隆與分析[D].重慶:西南大學,2021.
[11]" 陸兆亮,郭晉雅,羅誼,等.紫心甘薯類黃酮-3-糖基轉移酶基因及啟動子的克隆與功能分析//廣東省遺傳學會.廣東省遺傳學會第九屆代表大會暨學術研討會論文及摘要匯編[C].廣州,2014:59.
[12]" 王夢欽.外源茉莉酸甲酯對燈盞花抗旱性的影響及其相關基因挖掘[D].昆明:云南師范大學,2023.
[13] 龍慶梅.強化產業鏈助推生物產業高質量發展[J].云南農業,2022(9):26-29.
[14]" WANG Q, LIAO X L, XIANG C, et al. A practical synthesis of the flavone, scutellarein[J]. Journal of Chemical Research, 2017, 41(3): 157-159.
[15] DU Y W, MA H Y, LIU Y Y, et al. Major quality regulation network of flavonoid synthesis governing the bioactivity of black wolfberry[J]. The New Phytologist, 2024, 242(2): 558-575.
[16]" GUO X, YAN X, WANG Y, et al. Integrated transcriptomics and metabolomics analysis reveals the effects of cutting on the synthesis of flavonoids and saponins in Chinese Herbal Medicine Astragalus mongholious[J]. Metabolites, 2024,14(2): 428-443.
(責任編輯:易" 婧)
收稿日期:2024-02-28
基金項目:國家自然科學基金項目(3196020302)。
作者簡介:莫維淼(1998—),女,在讀碩士,研究方向為植物生物化學與分子生物學。E-mail:mokvimmiu@163.com。
*為通信作者,E-mail:zhyun-feng001@163.com。
猜你喜歡
四川蠶業(2021年2期)2021-03-09 03:15:32
四川蠶業(2021年3期)2021-02-12 02:38:46
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
哈爾濱醫藥(2016年1期)2017-01-15 13:43:16
天然產物研究與開發(2016年11期)2016-06-15 20:29:17
湖南師范大學自然科學學報(2015年1期)2015-02-27 14:50:04
安徽醫藥(2014年12期)2014-03-20 13:15:15
