賀蘭山巖羊活動節律與家域特征
2024-09-11 00:00:00崔爽胡天華余夢琦梁詠亮李岳誠郭松濤劉振生
野生動物學報 2024年3期

摘 要:野生動物的活動節律和家域特征是其為適應內外環境變化而形成的生存模式,研究活動節律和家域特征有助于深入了解野生動物生態適應性,維護生態平衡,并有助于制定科學保護策略。因此,基于北斗衛星追蹤項圈監測技術,深入研究了賀蘭山巖羊(Pseudois nayaur)全年的活動節律和家域特征,以及不同季節、性別的個體差異。研究表明:(1)巖羊屬于晨昏性動物(06:00—10:00、16:00—20:00),4個季節均表現為雙峰模式的日活動節律;(2)巖羊全年的活動速率為(0. 17±0. 51) km/h,且不同季節的活動速率存在顯著差異(χ2=133. 48,df=3,plt;0. 05),冬季活動速率最高,為0. 23 km/h;雌雄個體間活動速率存在邊緣顯著差異(Z=-1. 913,p=0. 056),雄性個體的活動速率高于雌性個體;(3)巖羊的家域和核域范圍無明顯季節性差異(家域,χ2=1. 623,df=3,p=0. 654;核域,χ2=0. 727,df=3,p=0. 867),性別間無顯著差異。建議保護團隊在晨昏高峰期加強監測,避免人為干擾;在冬季采取補飼等保護措施確保食物充足;并在制定管理計劃時充分考慮家域特征。
關鍵詞:巖羊;衛星項圈;活動節律;家域
中圖分類號:Q958. 1
文獻標識碼:A
文章編號:2310 - 1490(2024)- 03 - 0480 - 10
DOI:10.12375/ysdwxb.20240303
活動節律是動物對環境中多種因素適應的結果,主要受溫度、光照強度和食物資源量等多種因素調控[1?2]。活動節律的研究主要是探究動物在不同時間和季節下的活動強度及變化規律[3],作為動物行為學與生態學中的重要內容,活動節律研究有助于揭示動物對環境所采取的時間分配策略[4],還能有效反映動物個體營養狀態和生存壓力等重要參數[5]。家域(home range)對于研究動物的資源利用、環境適應、種群密度和社會關系等生態學過程具有關鍵作用[6]。家域是動物取食、交配和育幼等行為所利用的區域[7],受到種群密度和捕食風險等生態因子的影響。核域(core range)則是家域中最為重要的組成部分,辨別該區域對于全面理解動物對家域的利用至關重要[8]。
巖羊(Pseudois nayaur),又名藍羊、青羊,屬鯨偶蹄目(Cetartiodactyla)牛科(Bovidae),被列為國家二級重點保護野生動物。賀蘭山是巖羊的重要分布區之一,分為寧夏和內蒙古兩部分[9]。巖羊是典型的高山動物,主要棲息于山地草原帶、山地疏林草原帶、山地針葉林帶、亞高山灌叢和草甸帶等生境中,取食針茅(Stipa spp.)、旱榆(Ulmus glaucescens)和早熟禾(Poa spp.)等[10]。目前學者對巖羊的種群數量[11]、集群特征[12]、生境選擇及適宜性評價[13]、食性選擇和覓食對策[14]、行為學[15]、生理生化[16]、腸道菌群和性別二態性[17?18],以及性別分離機制[19]等方面進行了深入研究。但對于季節與性別如何影響活動節律和家域特征的研究較少。
GPS項圈技術能夠在不干擾動物的情況下,遠程監測野生動物的活動,提供更多的連續活動位置數據,并具備更長的監測時間和更大的監測范圍。雖然目前國內運用GPS 項圈研究有蹄類動物活動節律和家域特征的研究較為匱乏,但GPS項圈對于解釋動物節律與家域特征的優勢是極為明顯的。例如:吳運佳等[20]利用GPS跟蹤項圈跟蹤技術和布朗橋模型分別對野駱駝(Camelus ferus)活動節律和家域特征進行分析,結果表明野駱駝日活動節律呈現明顯雙峰模式,屬晨昏性動物,且為核心家域利用類型,存在多個核心家域。李耀宇等[21]利用北斗衛星項圈對浙江清涼峰國家級自然保護區內野外放歸華東梅花鹿(Cervus nippon kopschi)的活動軌跡進行了追蹤,結果表明季節對放歸華東梅花鹿的活動節律和家域大小具有較大影響。可見,基于GPS項圈可以有效研究野生動物種群的活動節律和家域特征。因此,本研究基于巖羊的生態學研究進展,采用北斗衛星追蹤項圈的監測技術,重點監測賀蘭山10只野生巖羊在2023年2—12月的活動節律和家域,共傳回45 961條追蹤數據,以此重點研究(1)不同季節、性別的巖羊個體活動節律是否存在差異;(2)不同季節、性別的巖羊個體的家域和核域是否存在差異。本研究旨在探究賀蘭山巖羊的節律行為與家域、核域空間特征,從而為賀蘭山巖羊種群的保護提供更加細致的方向,也為其保護政策的制定提供科學依據。
1 研究區概況
研究區位于寧夏賀蘭山國家級自然保護區(38°21′—39°22′ N,105°49′—106°42′ E),賀蘭山東坡,總面積193 535. 68 hm2。賀蘭山地區的氣候屬中溫帶干旱氣候,冬季寒冷漫長,夏季炎熱短暫,春季氣溫回升快,伴隨頻繁的大風和沙塵天氣,秋季則相對涼爽。在保護區內,野生維管植物647種,呈現出垂直分布的植被類型。賀蘭山地區的脊椎動物豐富,共有24目56科218種,其中包括一些珍稀獸類,如雪豹(Panthera uncia)、馬麝(Moschus chrysogaster)、巖羊和馬鹿(Cervus elaphus alxaicus)等[22]。
2 研究方法
2. 1 北斗衛星追蹤項圈的安裝
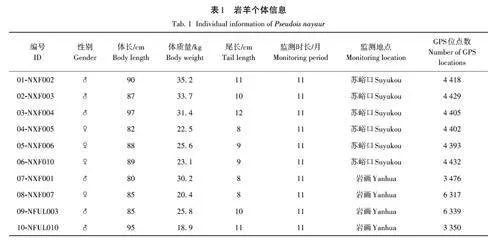
2022年2月17日,在寧夏賀蘭山國家級自然保護區的蘇峪口和賀蘭口捕獲10 只巖羊,雌雄比為2∶3。對捕獲的巖羊個體佩戴北斗衛星追蹤項圈(HQAB-M系列,湖南環球信士科技有限公司,項圈重量為1. 0~1. 6 kg)(圖1,表1)。佩戴北斗衛星追蹤項圈獲得寧夏賀蘭山國家級自然保護區管理局的許可。佩戴項圈的巖羊個體自放歸次日的00:00起每隔1 h 進行1 次位點的記錄,于2023 年4 月23 日(NXF002、NXF003、NXF004、NXF005、NXF006、NXF010、NFUL010)和24 日(NXF001)改為2 h 進行1次位點的記錄。項圈通過GSM網絡回傳時間、經緯度、海拔、活動量和運動速度等信息。
2. 2 活動節律分析
巖羊個體的活動強度是通過計算全天內的連續活動速率來反映,并對日活動節律、每個季節的活動速率和雌雄活動速率進行分析。利用ArcGIS 10. 8軟件,并結合Home Range Tools 2. 0(HRT 2. 0)插件計算每次活動位點之間的距離,獲得巖羊的活動速率(km/h)。同時,計算每個季節和雌雄個體的每小時平均活動速率。
2. 3 家域特征分析
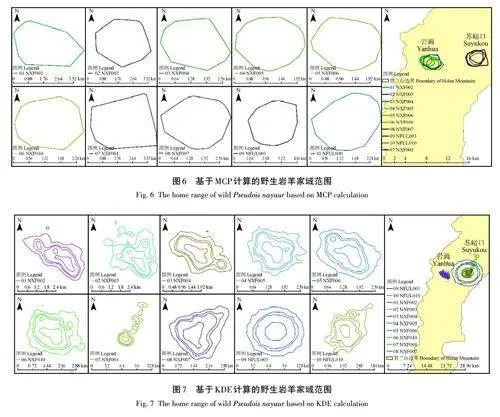
使用HRT 2. 0插件,由于最小凸多邊形法(minimumconvex polygon,MCP)計算的動物家域容易受異常點的影響,因此,每只巖羊的家域和核域采用MCP 和核心密度估算法(kernel density estimation,KDE)計算[23?25]。在KDE計算中,整體家域和核心家域分別采用99%和50%密度估計表示[20,26?27],并基于MCP和99% KDE分別計算不同季節、性別個體的平均家域范圍。
經單個樣本的Kolmogorov-Smirnov test檢驗,活動速率、家域面積和核域面積數據均不符合正態分布。因此,季節間對比采用非參數Kruskal-Wallis H檢驗不同組間是否存在顯著差異,并進行Mann-Whitney U 兩兩比較。不同性別間采用Mann-Whitney U 檢驗是否存在差異。所有數據結果采用平均值±標準誤(Mean±SE)表示,并使用SPSS 23. 0和Origin 2022進行數據分析。
3 結果
3. 1 項圈及個體情況
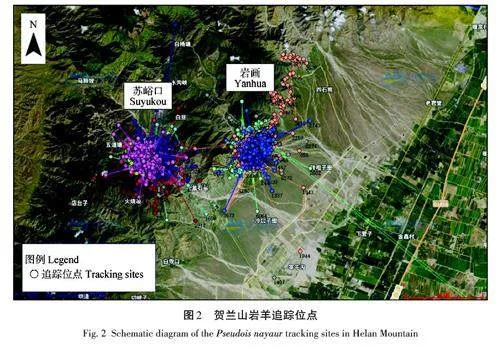
佩戴項圈的野生巖羊個體共10 只,無個體死亡,項圈均處于正常狀態,無脫落情況。項圈佩戴個體分別在蘇峪口和巖畫(賀蘭口)區域活動,其中,蘇峪口佩戴6枚項圈,巖畫佩戴4枚項圈。所有項圈均工作309 d以上,監測位點45 961個(圖2)。
3. 2 活動節律
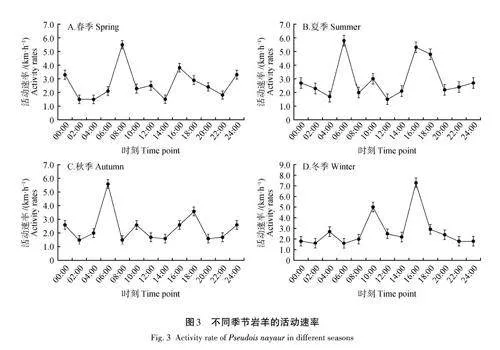
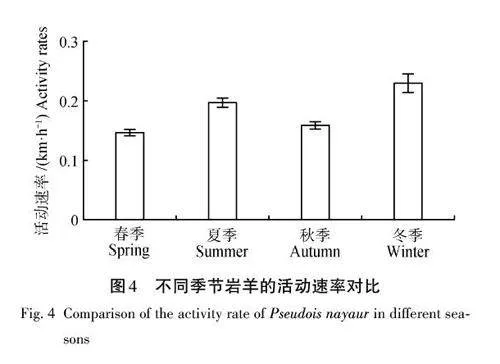
巖羊4個季節的日活動節律都表現為雙峰模式,屬于晨昏性動物(06:00—10:00、16:00—20:00)。4個季節活動高峰期不同,春夏秋季早晨活動高峰期相似(06:00—08:00),冬季早晨活動高峰期推遲至10:00—12:00。春夏冬季黃昏活動高峰期相同(16:00—18:00),秋季推遲至18:00—20:00(圖3)。此外,在春夏秋季24:00左右也有一個持續時間較短的高峰期,但低于晨昏高峰期活動速度。除冬季外,夜間的活動速率明顯低于早晨。
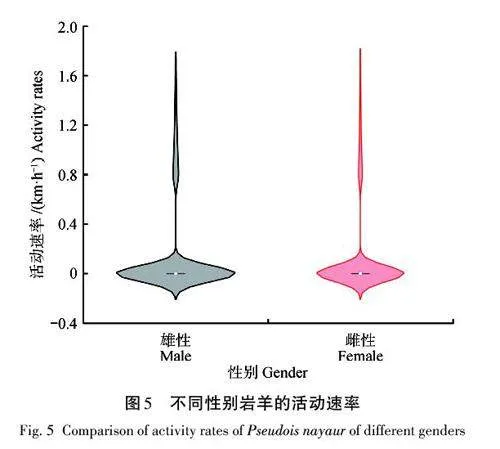
巖羊全年的活動速率為(0. 17±0. 51) km/h,且不同季節的活動速率存在顯著差異(χ2=133. 48,df=3,plt;0. 05),Mann-Whitney U 檢驗表明,季節間均存在顯著差異(春季×夏季,Z=-9. 427,plt;0. 05;春季×秋季,Z=-3. 380,plt;0. 05;春季×冬季,Z=-8. 878,plt;0. 05;夏季×秋季,Z=-5. 616,plt;0. 05;夏季×冬季,Z=-2. 808,plt;0. 05;秋季×冬季,Z=-6. 476,plt;0. 05)。其中,活動速率由高到低依次為冬季(0. 23 km/h)、夏季(0. 20 km/h)、秋季(0. 16 km/h)、春季(0. 15 km/h)(圖4)。不同性別巖羊個體的活動速率存在邊緣顯著差異(Z=-1. 913,p=0. 056),雌性個體平均活動速率為0. 17 km/h,雄性個體活動速率為0. 18 km/h( 圖5)。
3. 3 家域特征
3. 3. 1 家域面積
采用MCP計算4個季節巖羊的家域范圍,10只巖羊的MCP家域面積平均為(6. 93±3. 08) km2,平均周長為(9. 51±2. 32) km;核域面積平均為(0. 81±0. 28) km2,平均周長為(3. 18±0. 57) km(圖6)。采用KDE計算春夏秋冬4個季節巖羊的家域面積平均為(7. 20±4. 26) km2,平均周長為(16. 52±10. 94) km;核域面積平均為(0. 79±0. 37) km2,平均周長為(4. 96±3. 34) km(圖7)。
3. 3. 2 季節間的家域差異
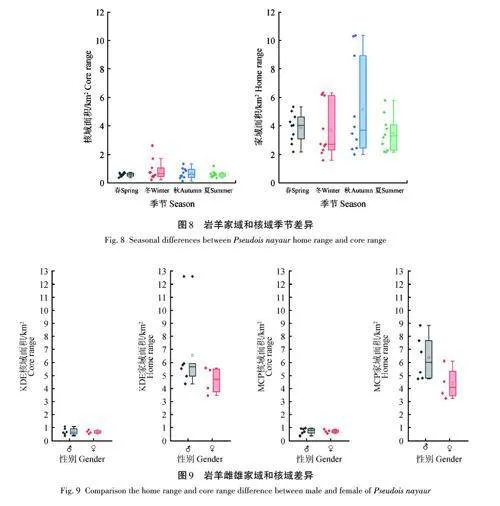
巖羊春季家域、核域面積分別為(3. 86±1. 02)、(0. 58±0. 12) km2,夏季分別為(3. 45±1. 22)、(0. 60±0. 24) km2,秋季分別為(5. 14±3. 35)、(0. 67±0. 37) km2,冬季分別為(3. 65±1. 86)、(0. 90±0. 73) km2。采用Kruskal-Wallis秩和檢驗表明,4個季節巖羊的家域和核域均不存在顯著性差異(家域,χ2=1. 623,df=3,p=0. 654;核域,χ2=0. 727,df=3,p=0. 867)(圖8)。
3. 3. 3 性別間的家域差異
采用MCP計算,雌性家域為(4. 39±1. 28) km2,核域為(0. 71±0. 14) km2;雄性家域為(6. 35±1. 69) km2,核域為(0. 73±0. 23) km2。采用KDE計算,雌性家域為(4. 63±1. 05) km2,核域為(0. 67±0. 13) km2;雄性家域為(6. 53±3. 02) km2,核域為(0. 69±0. 26) km2。雄性家域略大于雌性,但雌雄家域范圍不存在顯著性差異[家域,Z=-1. 229,p=0. 222;核域,Z=-0. 014,p=0. 989(MCP)。家域,Z=-1. 706,p=0. 171;核域,Z=-0. 107,p=0. 915(KDE)](圖9)。
4 討論
4. 1 巖羊活動節律特征
本研究通過GPS項圈對巖羊的活動節律進行分析,結果表明巖羊表現為晨昏活動,活動曲線在清晨和黃昏出現高峰期,這與崔多英[28]的研究結果一致,也與其他有蹄動物活動節律結果[21,29]相符。這種晨昏型活動方式是多種因素綜合作用的結果,對于巖羊等食草動物具有重要益處。在早晨氣溫相對凌晨有所回升,而在黃昏氣溫開始下降,這兩個時段都能避免高溫和強陽光,有利于動物的活動。晨昏時的露水能夠在一定程度上為動物提供水分補充[3]。考慮到賀蘭山部分區域是景區,存在一定的人為干擾[9],巖羊選擇晨昏活動時段也有助于規避這些潛在的人為干擾。
通過比較各季節活動速率,發現野生巖羊的活動強度具有季節性差異,冬季活動速率明顯高于其余3季,這可能是由于冬季的晝間時間變短、溫度下降較快,且食物資源匱乏,為獲得充足食物資源驅使巖羊活動速率增加。與其他野生動物相比,野駱駝活動強度高峰出現在夏季,這與夏季野駱駝天敵——狼(Canis lupus)的捕食活動時間高度重疊,為躲避天敵,活動強度增加[20,30]。賀蘭山地區巖羊種群缺少天敵[31],活動強度較低。雌雄個體間活動速率存在邊緣顯著差異,雄性個體的活動速率高于雌性個體,這可能與繁殖季節雄性較為活躍有關[32]。
4. 2 巖羊家域特征
本研究采用MCP和KDE兩種方法對野生巖羊的家域進行計算。與其他野生動物相比,巖羊的家域高于小麂(Muntiacus reevesi,1. 08 km2)[33],這是因為巖羊分布區域的植物種類相對較少,可利用的食物資源有限。因此,巖羊需要尋找更多的食物資源以滿足自身的需求,導致巖羊的家域相比其他一些有蹄類動物更大。
雖然基于MCP對比,季節間家域不存在顯著差異,但是從描述性統計值來看,家域面積均值仍有一定規律,家域面積由大到小依次是秋季、春季、冬季和夏季。這與梅花鹿、麋鹿(Elaphurus davidianus)家域面積季節間的變化規律[21,34]一致。秋季賀蘭山氣候適宜,食物豐富[35],且存在個體擴散行為[36],導致巖羊的家域面積增加。冬季巖羊的家域面積與春季接近,并均大于夏季,可能與保護區內食物豐富度和可獲得性差異有相關,在夏季,植物繁茂,種類多樣,食物的豐富度和可獲得性高;相比之下,春冬季由于大部分草本植物未長出或枯死,導致食物可利用性減少,豐富度和可獲得性相對較低[28]。同時,雄性家域略大于雌性,是由于在繁殖季節增加家域面積,可以提高其尋找雌性個體的機會,提高交配概率[32]。
本研究的雌雄個體家域面積比崔多英[28]研究所得家域[雄性家域(3. 74±0. 52) km2,雌性家域(2. 95±0. 40) km2]更大,可能存在三方面原因:一是本研究利用了巖羊所有時期的活動點數據,二是相比于崔多英[28]所使用的MOD-500 Telemetry Collar項圈精確度和靈敏度更高,三是相比于以前所使用的單一MCP算法易受特異位點影響的不足[37]。KDE利用核密度對家域進行計算,能夠排除一些特異位點的影響,更加準確地反映家域范圍特征,是對MCP計算結果的有效補充[24]。
綜上所述,季節對巖羊的活動速率和家域面積大小有不同程度的影響,在性別方面的差異仍然需要更大的樣本量和更加深入的研究。因而,未來的研究中可擴大研究區域和樣本量,進一步研究家域大小與性別之間的關系,特別是在繁殖等關鍵時期,深化對巖羊生態學的全面了解。還需要關注環境因素,如溫度和植被對巖羊活動模式的影響,并探討在風景區人為干擾的情況下,巖羊選擇晨昏活動時段的原因,有助于制定更科學的保護策略。
4. 3 保護和管理建議
巖羊表現出晨昏活動的特征,這一活動方式對于食草動物具有重要益處。建議保護管理團隊在晨昏高峰期加強對巖羊的巡邏和監測,以更好地了解其行為習慣和遷移路徑。考慮到季節變化,特別是食物匱乏的冬季,建議建立補飼點和增加救護巡邏次數等,以確保巖羊在冬季可獲得充足的食物資源。家域范圍的大小對于理解巖羊的空間利用至關重要。本研究結果中雄性家域略大,可能與繁殖期有關,建議加強保護措施以促進繁殖成功,在制定保護區域或管理計劃時,充分考慮巖羊家域的需求情況,以減少對其生態習性的干擾。
參考文獻:
[1] 尚玉昌. 動物的行為節律[J]. 生物學通報, 2006, 41(10):
8-10.
SHANG Y C. Behavior rhythm of animals[J]. Bulletin of Biology,
2006, 41(10): 8-10.
[2] DIBNER C, SCHIBLER U. Circadian timing of metabolism in
animal models and humans[J]. Journal of Internal Medicine,
2015, 277(5): 513-527.
[3] 原寶東, 孔繁繁. 哺乳動物活動節律研究進展[J]. 安徽農業
科學, 2011, 39(2): 1056-1058;1162.
YUAN B D, KONG F F. Research on mammal activity rhythm
[J]. Journal of Anhui Agricultural Sciences, 2011, 39(2):
1056-1058;1162.
[4] EVERS D C. Activity budgets of a marked common loon (Gavia
immer) nesting population[J]. Hydrobiologia, 1994, 279(1):
415-420.
[5] HALLE S, STENSETH N C. Activity patterns in small mammals:
an ecological approach[M]//CALDWELL M M, HELDMAIER G,
LANGE O L, et al. Ecological studies:Vol. 141: analysis and
synthesis. Berlin: Springer-Verlag, 2000.
[6] 張晉東, HULL V, 歐陽志云. 家域研究進展[J]. 生態學報,
2013, 33(11): 3269-3279.
ZHANG J D, HULL V, OUYANG Z Y. A review of home range
studies[J]. Acta Ecologica Sinica, 2013, 33(11): 3269-3279.
[7] BURT W H. Territoriality and home range concepts as applied to
mammals[J]. Journal of Mammalogy, 1943, 24(3): 346-352.
[8] SAMUEL M D, PIERCE D J, GARTON E O. Identifying areas of
concentrated use within the home range[J]. The Journal of Animal
Ecology, 1985, 54(3): 711-719.
[9] 張明春. 賀蘭山巖羊的種群動態及警戒行為研究[D]. 上海:
華東師范大學, 2012.
ZHANG M C. Population dynamics and flight responses of blue
sheep in Ningxia Heelan Mountain National Nature Reserve[D].
Shanghai: East China Normal University, 2012.
[10] 駱穎. 賀蘭山巖羊(Pseudois nayaur)和馬鹿(Cervus elaphus
alxaicus)的食性及生境選擇比較研究[D] 哈爾濱: 東北林業
大學, 2011.
LUO Y. Comparing the diet and habitat selection of sympatric
blue sheep( Pseudois nayaur) and red deer( Cervus elaphus alxa?
icus) in Helan Mountains, China[D]. Harbin: Northeast Forestry
University, 2011.
[11] THAPA K, JACKSON R, GURUNG L, et al. Applying the
double observer methodology for assessing blue sheep population
size in Nar Phu Valley, Annapurna Conservation Area, Nepal
[J]. Wildlife Biology, 2021, 2021(4): wlb. 00877.
[12] 曹麗榮. 巖羊(Pseudois nayaur)的社群結構和保護遺傳學研
究[D]. 上海: 華東師范大學, 2006.
CAO L R. Group behaviour and conservation genetics of bharal
(Pseudois nayaur) in Helan Mountain, China[D]. Shanghai:
East China Normal University, 2006.
[13] NAMGAIL T, FOX J L, BHATNAGAR Y V. Habitat segregation
between sympatric Tibetan argali Ovis ammon hodgsoni and
blue sheep Pseudois nayaur in the Indian Trans-Himalaya[J].
Journal of Zoology, 2004, 262(1): 57-63.
[14] LIU Z S, ZHU Z L, GAO H, et al. Comparative analysis of winter
diets and habitat use by the sympatric blue sheep (Pseudois
nayaur) and Alashan red deer (Cervus alashanicus) in the Helan
Mountains, China[J]. Folia Zoologica, 2018, 67(1): 43-53.
[15] LI Z Z, WANG J F, KHATTAK R H, et al. Coexistence mechanisms
of sympatric ungulates: behavioral and physiological adaptations
of blue sheep( Pseudois nayaur) and red deer( Cervus ela?
phus alxaicus) in Helan Mountains, China[J]. Frontiers in Ecology
and Evolution, 2022, 10: 925465.
[16] MENG D H, SI Y H, WANG J F, et al. The effect of sexes and
seasons on the morphological structures of the ruminant digestive
system of blue sheep (Pseudois nayaur)[J]. Animals, 2023, 13
(6): 1084.
[17] SUN G L, XIA T, WEI Q G, et al. Analysis of gut microbiota in
three species belonging to different genera (Hemitragus, Pseu?
dois, and Ovis) from the subfamily Caprinae in the absence of environmental
variance[J]. Ecology and Evolution, 2021, 11
(17): 12129-12140.
[18] ZHU Z L, SUN Y W, ZHU F, et al. Seasonal variation and
sexual dimorphism of the microbiota in wild blue sheep (Pseudois
nayaur)[J]. Frontiers in Microbiology, 2020, 11: 1260.
[19] 張明明. 賀蘭山巖羊(Pseudois nayaur)性別分離機制研究
[D]. 哈爾濱: 東北林業大學, 2013.
ZHANG M M. Research on the sexual segregation of blue sheep
(Pseudois nayaur) in the Helan Mountains, China[D]. Harbin:
Northeast Forestry University, 2013.
[20] 吳運佳, 程蕓, 袁磊, 等. 庫木塔格沙漠地區野駱駝活動節
律與家域特征[J]. 生物多樣性, 2021, 29(9): 1206-1214.
WU Y J, CHENG Y, YUAN L, et al. Quantitative study on the
activity rhythm and home range of wild camels (Camelus ferus)
in the Kumtag Desert[J]. Biodiversity Science, 2021, 29(9):
1206-1214.
[21] 李耀宇, 陳昕旸, 趙閃閃, 等. 野外放歸華南梅花鹿活動節
律與家域特征[J]. 獸類學報, 2023, 43(5): 501-512.
LI Y Y, CHEN X Y, ZHAO S S, et al. Activity rhythm and
home range of released sika deer (Cervus nippon kopschi)[J].
Acta Theriologica Sinica, 2023, 43(5): 501-512.
[22] 趙唱. 賀蘭山馬麝的營養采食策略和微生境選擇及評價研究
[D]. 哈爾濱: 東北林業大學, 2018.
ZHAO C. Nutrition diet strategy, micro-habitat selection and
evaluation of alpine musk deer (Moschus sifanicus) in Helan
Mountains, China[D]. Harbin: Northeast Forestry University,
2018.
[23] CAREY A B, REID J A, HORTON S P. Spotted owl home
range and habitat use in southern Oregon Coast Ranges[J]. The
Journal of Wildlife Management, 1990, 54(1): 11-17.
[24] WERDEL T J, JENKS J A, KANTA J T, et al. Space use and
movement patterns of translocated bighorn sheep[J]. Mammalian
Biology, 2021, 101(3): 329-344.
[25] HAVEMANN C P, RETIEF T A, COLLINS K, et al. Home
range and habitat use of roan antelope Hippotragus equinus in
northern Botswana[J]. Journal of Arid Environments, 2022,
196: 104648.
[26] DVO?áK S, BARTáK V, MACHá?EK Z, et al. Home range
size and spatio-temporal dynamics of male sika deer (Cervus nip?
pon; Cervidae, Artiodactyla) in an introduced population[J].
Folia Zoologica, 2014, 63(2): 103-115.
[27] 劉嘉輝, 王艷, 邊坤, 等. 重引入林麝的家域利用與個體遷
移[J]. 獸類學報, 2020, 40(2): 109-119.
LIU J H, WANG Y, BIAN K, et al. Home range utilization and
individual dispersal of re-introduced forest musk deer (Moschus
berezovskii) [J]. Acta Theriologica Sinica, 2020, 40(2):
109-119.
[28] 崔多英. 賀蘭山巖羊(Pseudois nayaur)的家域、活動規律和采
食生態學研究[D]. 上海: 華東師范大學, 2007.
CUI D Y. Home range, activity rhythm and foraging strategy of
bharal( Pseudois nayaur) in Helan Mountain, China[D]. Shanghai:
East China Normal University, 2007.
[29] 程建祥, 黃相相, 陳東東, 等. 清涼峰國家級自然保護區華
南梅花鹿分布及活動規律[J]. 生態學報, 2018, 38(22):
8213-8222.
CHENG J X, HUANG X X, CHEN D D, et al. Study on the distribution
and activity patterns of Cervus pseudaxis in the Qingliangfeng
National Nature Reserve[J]. Acta Ecologica Sinica,
2018, 38(22): 8213-8222.
[30] 袁國映. 野雙峰駝:沙漠戈壁里的“流浪漢”[J]. 生命世界,
2004(5): 24-33.
YUAN G Y. Wild bactrian camel: a“ tramp” in the gobi desert
[J]. Life World, 2004(5): 24-33.
[31] 劉振生, 王小明, 李志剛, 等. 賀蘭山巖羊冬春季取食生境
的比較[J]. 動物學研究, 2005, 26(6): 580-589.
LIU Z S, WANG X M, LI Z G, et al. Comparison of seasonal
feeding habitats by blue sheep (Pseudois nayaur) during winter
and spring in Helan Mountain, China[J]. Zoological Research,
2005, 26(6): 580-589.
[32] 謝建沖. 寧夏賀蘭山巖羊(Pseudois nayaur alashanicus)種群
數量及種群結構研究[D]. 哈爾濱: 東北林業大學, 2022.
XIE J C. Population size and structure of blue sheep in Helan
Mountains National Nature Reserve, Ningxia[D]. Harbin:
Northeast Forestry University, 2022.
[33] MCCULLOUGH D R, PEI K C J, WANG Y. Home range, activity
patterns, and habitat relations of Reeves’ muntjacs in Taiwan
[J]. Journal of Wildlife Management, 2000, 64(2): 430-441.
[34] DING J J, CHANG Q, DING Y H, et al. Seasonal home range
patterns of the reintroduced and rewild female Père David’s deer
Elaphurus davidianus[J]. Biological Rhythm Research, 2017,
48(3): 485-497.
[35] 丁彥珂. 賀蘭山地區赤狐(Vulpes vulpes)與流浪狗(Canis fa?
miliaris)生境選擇及生境適宜性比較研究[D]. 哈爾濱: 東北
林業大學, 2023.
DING Y K. A study on comparing the habitat selection and habitat
suitability assessment of red fox (Vulpes vulpes) and stray dog
(Canis familiaris) in Helan Mountains[D]. Harbin: Northeast
Forestry University, 2023.
[36] 劉旭. 基于分子生物學的巖羊個體遷移擴散研究[D]. 上海:
華東師范大學, 2014.
LIU X. The individual dispersal mechanism of blue sheep based
on molecular biology[D]. Shanghai: East China Normal University,
2014.
[37] HARRIS S, CRESSWELL W J, FORDE P G, et al. Homerange
analysis using radio-tracking data: a review of problems
and techniques particularly as applied to the study of mammals
[J]. Mammal Review, 1990, 20(2/3): 97-123.
