基于DNA 宏條形碼技術(shù)的斑海豹食性分析
2024-09-11 00:00:00高祥剛夏瑩王震邢衍闊鹿志創(chuàng)田甲申
野生動(dòng)物學(xué)報(bào) 2024年3期
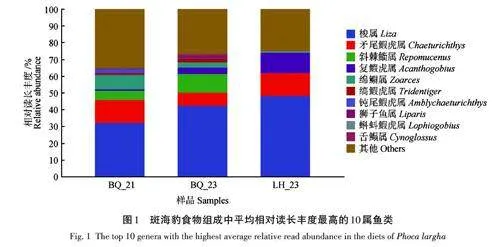

摘 要:野生動(dòng)物食性研究是掌握動(dòng)物生境需求的核心內(nèi)容,對野生動(dòng)物的保護(hù)和管理具有重要意義。在我國遼東灣斑海豹(Phoca largha)傳統(tǒng)繁殖地和盤錦棲息地海域采集其糞便,選用12S rRNA作為分子標(biāo)記進(jìn)行糞便DNA擴(kuò)增,利用高通量測序鑒定其食物組成。結(jié)果發(fā)現(xiàn):在斑海豹糞便中共鑒定出魚類16種,隸屬5目8科13屬。食物組成的相對豐度顯示:梭魚(Liza haematocheilus)為絕對優(yōu)勢的餌料食物(40. 72%),其次為鰕虎科(Gobiidae)種類(23. 18%)。屬水平不同采樣群體的相對豐度顯示:2021年、2023年遼東灣北部冰區(qū)和2023年盤錦遼河口棲息地排在前3位的種類分別是梭屬(Liza)31. 91%、矛尾鰕虎屬(Chaeturichthys)14. 06%和縞鰕虎屬(Tri?dentiger)8. 39%,梭屬42. 37%、復(fù)鰕虎屬(Acanthogobius)14. 06% 和矛尾鰕虎屬11. 17%,及梭屬47. 93%、矛尾鰕虎屬13. 93%和綿鳚屬(Zoarces)12. 18%,分別合計(jì)占斑海豹3個(gè)群體食物組成相對豐度的54. 36%、67. 60%和74. 04%。研究結(jié)果與遼東灣北部的漁業(yè)資源優(yōu)勢物種一致,表明斑海豹為廣食性物種,其食物組成主要取決于棲息海域、季節(jié)及主要獵物種類的豐度。
關(guān)鍵詞:斑海豹;DNA宏條形碼;12S rRNA;食性分析;繁育期
中圖分類號:Q958.1
文獻(xiàn)標(biāo)識碼:A
文章編號:2310 - 1490(2024)- 03 - 0498 - 06
DOI:10.12375/ysdwxb.20240305
野生動(dòng)物食性研究是掌握動(dòng)物生境需求的核心內(nèi)容,這對加強(qiáng)野生動(dòng)物的保護(hù)和管理有非常重要的意義[1]。傳統(tǒng)的食性分析方法如直接觀察法、利用法、胃容物分析法和糞便顯微鑒定法等,以及現(xiàn)代的食性分析方法如穩(wěn)定同位素技術(shù)法、脂肪酸法等均各有利弊,一些涉及解剖或?qū)?dòng)物造成傷害的操作可能會(huì)對動(dòng)物個(gè)體造成損傷,破壞野生動(dòng)物資源,不利于野生動(dòng)物保護(hù)[2?3]。隨著現(xiàn)代高通量測序技術(shù)的不斷發(fā)展,DNA宏條形碼技術(shù)成為現(xiàn)代食性研究的一種重要方法,該技術(shù)基于二代測序技術(shù)獲得生物特異性基因識別DNA 條形碼序列,通過對糞便[4?5]、胃容物[6]等樣本測序,借助已有數(shù)據(jù)庫序列比對,實(shí)現(xiàn)食物種類的分類鑒定;該技術(shù)還可通過比較所得物種糞便序列的相對豐度來定量描述該物種的食物組成,進(jìn)而實(shí)現(xiàn)食性研究的定性及定量分析,尤其適用于野生保護(hù)動(dòng)物的食性研究[7?8]。
斑海豹(Phoca largha)屬國家一級重點(diǎn)保護(hù)野生動(dòng)物,在我國主要分布于渤海和黃海海域,偶見于東海和南海,是渤海和黃海海洋生態(tài)系統(tǒng)的旗艦物種。斑海豹是唯一能在中國海域進(jìn)行繁殖的鰭足類海洋哺乳動(dòng)物,渤海遼東灣結(jié)冰區(qū)是世界上斑海豹8個(gè)繁殖區(qū)中最南端的一個(gè)[9]。每年11月以后,斑海豹陸續(xù)進(jìn)入渤海的遼東灣繁育后代,翌年5月上中旬后陸續(xù)離開渤海。目前分布于我國海域的斑海豹群體數(shù)量僅約2 000頭,因而受到國內(nèi)外學(xué)者的普遍重視[9?10]。在寒冷的冬春季節(jié),渤海漁業(yè)資源較匱乏,在此繁育和棲息的斑海豹攝取食物的種類是科研工作者關(guān)注的重點(diǎn)之一。然而目前關(guān)于野外斑海豹的食物來源報(bào)道僅見胃容物法和同位素法的研究結(jié)果,且研究時(shí)間和海域并未完全覆蓋斑海豹在我國海域分布的冬春季節(jié)[11?13]。利用DNA宏條形碼技術(shù)分析野外斑海豹食性的研究尚無報(bào)道。本研究首次利用DNA宏條形碼技術(shù)對遼東灣北部冰區(qū)和盤錦遼河口棲息海域的繁育期斑海豹的糞便樣本進(jìn)行采集與分析,提取DNA后選用12S rRNA為分子標(biāo)記分析斑海豹的食物組成及攝食選擇性,解析DNA宏條形碼技術(shù)在其食性研究中的應(yīng)用潛能,為斑海豹保護(hù)提供科學(xué)依據(jù),也為其他野生瀕危保護(hù)物種的攝食研究提供借鑒。
1 材料與方法
1. 1 樣品采集
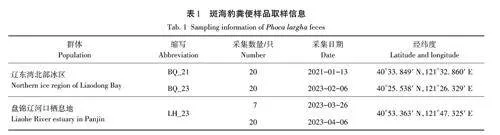
2021年1月13日和2023年2月6日在遼東灣北部冰區(qū)采集斑海豹糞便各20份。2023年3月26日和4月6日在斑海豹重要棲息地——盤錦遼河口海域分別采集斑海豹糞便7份和20份。用一次性PE手套拾取單個(gè)糞便樣品分別裝入5 mL的無菌凍存管中,在低溫保溫箱中保存運(yùn)至實(shí)驗(yàn)室,超低溫冰箱中-80 ℃保存?zhèn)溆谩>唧w采樣信息見表1。
1. 2 DNA 提取、PCR 擴(kuò)增及測序
采用常規(guī)酚氯仿法提取新鮮糞便的總DNA,溶于TE緩沖液中,-40 ℃保存?zhèn)溆谩NA濃度和質(zhì)量使用NanoDrop 2000核酸蛋白分析儀測量,經(jīng)檢驗(yàn)合格后,進(jìn)行PCR擴(kuò)增。使用線粒體12S rRNA(魚類)通用引物對總DNA 進(jìn)行PCR 擴(kuò)增。引物序列為:MiFish-U-F: 5′-GTCGGTAAAACTCGTGCCAGC-3′,MiFish-U-R: 5′-CATAGTGGGGTATCTAATCCCAGTTTG-3′[14]。PCR擴(kuò)增反應(yīng)體系為50. 0 μL,包括總DNA2. 0 μL(200 ng),rTaq DNA聚合酶1. 0 μL(5 U/μL),dNTPs 4. 0 μL(各2. 5 mmol/L),上、下游引物各2. 0 μL,10×PCR buffer 5. 0 μL,雙蒸水34. 0 μL。PCR 擴(kuò)增反應(yīng)程序?yàn)?8 ℃預(yù)變性1 min;98 ℃變性10 s,57 ℃退火30 s,72 ℃延伸30 s,35個(gè)循環(huán);最后72 ℃延伸10 min。PCR產(chǎn)物經(jīng)1. 5% 瓊脂糖凝膠電泳純化后選擇主帶大小約200 bp的序列,割膠回收目的條帶。PCR擴(kuò)增產(chǎn)物建庫檢測,合格的文庫送上海派森諾生物科技股份有限公司進(jìn)行高通量測序。
1. 3 數(shù)據(jù)處理與分析
使用Illumina PE250 測序平臺,采用雙端測序法,得到下機(jī)數(shù)據(jù),去除引物接頭序列,根據(jù)PEreads之間的重疊關(guān)系,將成對的讀長拼接成一條序列,然后按照標(biāo)簽序列識別并區(qū)分樣品得到各樣本數(shù)據(jù),最后經(jīng)質(zhì)控、過濾和拼接得到有效數(shù)據(jù)。
通過Excel統(tǒng)計(jì)分析OTUs在種、屬分類水平下的相對豐度;選取OTUs相對豐度值最高的種類與美國NCBI NT(https://www. ncbi. nlm. nih. gov/)數(shù)據(jù)庫比對,進(jìn)行物種注釋并人工校對,篩選出序列的最佳比對結(jié)果。序列的物種鑒定原則:12S rDNA序列在相同的基因序列區(qū)域內(nèi),當(dāng)覆蓋度(coverage)不低于95. 0%且一致度達(dá)到99. 5%及以上時(shí),判定樣品來自匹配序列對應(yīng)的物種;最高一致度lt;99. 5% 且≥95. 0%,一致度最高序列僅有1條,且與得分次之的序列一致度差距≥2%時(shí),記為一致度最高序列的上一級分類單元,最高一致度對應(yīng)有多個(gè)物種時(shí),則記為能夠涵蓋所有一致度最高的當(dāng)?shù)匚锓N的最低分類單元[4,15]。通過OTU分析鑒定糞便中主要獵物物種的分類和不同物種的相對豐度。
2 結(jié)果與分析
2. 1 分類地位鑒定
糞便樣本共獲得原始序列9 055 417條,剔除斑海豹的自身序列以及豐度較低的序列后,共獲得有效序列692 167 條。按照97% 相似性聚類后得到390條OTU代表序列,注釋得到16種餌料生物,分屬于5目8科13屬(表2)。
2. 2 食物組成
斑海豹糞便中食物組成的相對豐度顯示,梭魚(Liza haematocheilus)為絕對優(yōu)勢的餌料食物,相對豐度為40. 72%;其次為鰕虎科(Gobiidae)的種類,包含5個(gè)屬(表2),占相對豐度的23. 18%。從屬水平不同群體的平均相對讀長豐度看,2021年遼東灣北部冰區(qū)群體(BQ_21)、2023年遼東灣北部冰區(qū)群體(BQ_23)和2023年盤錦遼河口棲息地群體(LH_23)排在前3位的食物種類分別是梭屬31. 91%、矛尾鰕虎屬(Chaeturichthys)14. 06% 和縞鰕虎屬(Triden?tiger)8. 39%,梭屬42. 37%、復(fù)鰕虎屬(Acan?thogobius)14. 06% 和矛尾鰕虎屬11. 17%,及梭屬47. 93%、矛尾鰕虎屬13. 93% 和綿鳚屬(Zoarces)12. 18%;分別合計(jì)占斑海豹3個(gè)群體食物組成相對豐度的54. 36%、67. 60%和74. 04%(圖1)。研究結(jié)果表明,DNA宏條形碼技術(shù)在獲取食物種類信息和食物豐度分析上應(yīng)用的可行性,證明了DNA宏條形碼技術(shù)在動(dòng)物食性研究中具有較高的應(yīng)用潛能。
3 討論
近年來,DNA宏條形碼技術(shù)憑借明顯的技術(shù)優(yōu)勢,在野生動(dòng)物的食性研究中得到較廣泛應(yīng)用[2,16]。邵昕寧等[4]利用宏條形碼技術(shù)從不同糞便樣本中分析出我國西南山地四川甘孜藏族自治州的7種食肉動(dòng)物的食性組成,包括棕熊(Ursus arctos)、豹(Pan?thera pardus)、雪豹(P. uncia)、豹貓(Prionailurus ben?galensis)、狼(Canis lupus)、狗以及赤狐(Vulpesvulpes)。金森龍等[5]基于DNA宏條形碼技術(shù)在臥龍國家級自然保護(hù)區(qū)食肉動(dòng)物的食性分析中發(fā)現(xiàn)赤狐的食物主要來自3個(gè)目,豹貓的食物來自5個(gè)目,初步弄清了臥龍保護(hù)區(qū)內(nèi)野生食肉動(dòng)物的食物組成。宮爽等[7]利用DNA宏條形碼技術(shù)獲得我國東北地區(qū)野豬(Sus scrofa)食物組成,包含60屬植物,比例大于4%的主要食物有6個(gè)屬。滕揚(yáng)等[8]通過DNA宏條形碼技術(shù)在豹貓食物構(gòu)成分析中,鑒別出來自10目22科的36種獵物,表明北京市4個(gè)保護(hù)區(qū)的豹貓食性具有顯著差異,發(fā)現(xiàn)基于DNA宏條形碼技術(shù)鑒定出動(dòng)物性獵物種類較為齊全。本研究從不同糞便樣本中分析出我國遼東灣斑海豹的食物主要來自13個(gè)屬;同時(shí),在斑海豹糞便中鑒定出兩個(gè)相對低豐度的淡水種類:鯉(Cyprinus carpio)和北鰍(Lefua costata),分別占相對豐度的0. 13%和0. 05%,但查找其取樣群體發(fā)現(xiàn),兩個(gè)淡水種類僅出現(xiàn)在盤錦遼河口棲息地的群體中,分析這可能是斑海豹到遼河口附近取食的原因。
盡管DNA宏條形碼技術(shù)因其明顯的優(yōu)勢在動(dòng)物食性研究領(lǐng)域得到應(yīng)用,但也存在一些不足,即在PCR擴(kuò)增反應(yīng)中,效率高的模板很可能會(huì)對效率低的產(chǎn)生抑制作用,導(dǎo)致在PCR擴(kuò)增中受到抑制的食物類群被低估甚至無法被鑒別[17]。田甲申等[12]利用同位素方法研究發(fā)現(xiàn)野外斑海豹的食物來源以魚類為主,同時(shí)也攝食一些頭足類和蝦類。但本研究使用無脊椎動(dòng)物COI通用引物(LCO1490/HCO2198)及宏條形碼引物(mlCOIintF/jgHCO2198R)對糞便樣品進(jìn)行分析后,擴(kuò)增出大量無效序列、污染序列,未發(fā)現(xiàn)頭足類和蝦類的序列。但隨著DNA條形碼數(shù)據(jù)庫的不斷完善和更為適宜的分子標(biāo)記的優(yōu)化和被發(fā)現(xiàn),DNA宏條形碼技術(shù)的檢出準(zhǔn)確度將會(huì)大幅提高[18]。未來,DNA宏條形碼技術(shù)聯(lián)合其他技術(shù)手段,將為野生動(dòng)物食性分析的應(yīng)用開啟更廣闊的空間[17]。
關(guān)于野外斑海豹的食性研究國內(nèi)外學(xué)者開展了一些工作,Bukhtiyarov等[19]利用胃容物方法研究發(fā)現(xiàn)斑海豹食物的多樣性以及食物的地域和季節(jié)差異與其主要獵物種類的季節(jié)分布和豐度有關(guān),斑海豹的食物存在區(qū)域差異性;同時(shí)發(fā)現(xiàn),斑海豹食物以魚類為主,無脊椎動(dòng)物獵物在其食物中被發(fā)現(xiàn)的頻率很低。Bukhtiyarov等[19]在春季白令海北部的斑海豹胃中發(fā)現(xiàn)魚類占比超過95. 0%,俄羅斯阿納迪厄海灣斑海豹胃中魚類占比超過52. 2%;同時(shí)發(fā)現(xiàn),斑海豹對食物的選擇也與年齡有關(guān),1~4齡主要捕食魚、體型較大的蝦類,偶爾也捕食頭足類,而4齡以上主要捕食魚類。Bukhtiyarov等[19]發(fā)現(xiàn)在白令海北部,斑海豹主要捕食北極鱈(Boreogadus saida);在白令海中部、鄂霍茨克海,主要捕食黃線狹鱈(Theragrachalcogramma);在白令海東南部,主要捕食毛鱗魚(Mallotus villosus);在楚科奇海東南部,主要捕食鯡魚(Clupea pallasii);在日本海、阿拉斯加的近海等地,捕食鱈、鮭等魚類。王者茂等[11]研究發(fā)現(xiàn)在黃、渤海海域斑海豹春季捕食鯡魚、太平洋玉筋魚(Am?modytes personatus)和小黃魚(Larimichthys polyactis)等,秋冬季節(jié)則多以梭魚為食。劉修澤等[20]在遼東灣北部海域魚類群落結(jié)構(gòu)及多樣性研究中,指出遼東灣北部冬春季節(jié)魚類資源以梭魚、鰕虎魚為主。因此,本研究結(jié)果與上述研究相似,為在繁育季節(jié)加強(qiáng)對斑海豹的保護(hù)和漁業(yè)資源養(yǎng)護(hù)提供了科學(xué)管理依據(jù)。
4 結(jié)論
本研究利用糞便DNA宏條形碼技術(shù)對繁育季節(jié)遼東灣斑海豹的食性進(jìn)行了分析,結(jié)果顯示,斑海豹糞便中共鑒定出魚類16種,隸屬5目8科13屬。根據(jù)食物組成的相對豐度,梭魚為絕對優(yōu)勢的餌料食物,相對豐度為40. 72%,其次為鰕虎科的種類,相對豐度為23. 18%。研究結(jié)果與遼東灣北部的漁業(yè)資源優(yōu)勢物種相一致,表明我國遼東灣斑海豹為廣食性物種,其食物組成主要取決于棲息海域、季節(jié)及主要獵物種類的豐度。遼東灣斑海豹在冬春繁育季節(jié)以梭魚、鰕虎魚等為食,本研究對掌握野生動(dòng)物生境需求、加強(qiáng)野生動(dòng)物的管理和保護(hù)有著非常重要的意義。
參考文獻(xiàn):
[1] 司曉艷, 趙天飆, 毛永強(qiáng), 等. 食草動(dòng)物食性研究方法的評價(jià)
[J]. 醫(yī)學(xué)動(dòng)物防制, 2007, 23(12): 957-959.
SI X Y, ZHAO T B, MAO Y Q, et al. The review and evaluation
on the study method for herbivore food-habit[J]. Journal of Medical
Pest Control, 2007, 23(12): 957-959.
[2] 黃乃瑩, 劉艷華. 分子糞便學(xué)在野生動(dòng)物食性研究中的應(yīng)用
[J]. 野生動(dòng)物學(xué)報(bào), 2022, 43(1): 266-269.
HUANG N Y, LIU Y H. Application of molecular scatology in the
study of wild animal feeding habits[J]. Chinese Journal of Wild‐
life, 2022, 43(1): 266-269.
[3] 張健, 王佚兮, 馮慧敏, 等. 基于脂肪酸標(biāo)記法和穩(wěn)定同位素
技術(shù)的通州灣養(yǎng)殖水域海蜇食性分析[J]. 南方水產(chǎn)科學(xué),
2021, 17(1): 25-31.
ZHANG J, WANG Y X, FENG H M, et al. Analysis of feeding
habits of cultured jellyfish (Rhopilema esculentum) in Tongzhou
Bay based on fatty acid and stable carbon and nitrogen isotopic
analysis[J]. South China Fisheries Science, 2021, 17(1): 25-31.
[4] 邵昕寧, 宋大昭, 黃巧雯, 等. 基于糞便DNA及宏條形碼技術(shù)
的食肉動(dòng)物快速調(diào)查及食性分析[J]. 生物多樣性, 2019, 27
(5): 543-556.
SHAO X N, SONG D Z, HUANG Q W, et al. Fast surveys and
molecular diet analysis of carnivores based on fecal DNA and metabarcoding
[J]. Biodiversity Science, 2019, 27(5): 543-556.
[5] 金森龍, 瞿春茂, 施小剛, 等. 臥龍國家級自然保護(hù)區(qū)食肉動(dòng)
物多樣性及部分物種的食性分析[J]. 野生動(dòng)物學(xué)報(bào), 2021,
42(4): 958-964.
JIN S L, QU C M, SHI X G, et al. The species diversity and diet
of some carnivores in Wolong National Nature Reserve, Sichuan
Province[J]. Chinese Journal of Wildlife, 2021, 42(4): 958-964.
[6] 陳曉雷, 李敏, 陳作志, 等. 基于宏條形碼技術(shù)的南海亮眶燈
魚食性初步分析[J]. 南方水產(chǎn)科學(xué), 2022, 18(3): 22-29.
CHEN X L, LI M, CHEN Z Z, et al. Preliminary metabacording
dietary analysis of Diaphus splendidus in South China Sea[J].
South China Fisheries Science, 2022, 18(3): 22-29.
[7] 宮爽, 張萍, 欒雪, 等. 糞便顯微分析和DNA宏條形碼技術(shù)在
野豬食性分析中的應(yīng)用和比較[J]. 野生動(dòng)物學(xué)報(bào), 2022, 43
(3): 645-653.
GONG S, ZHANG P, LUAN X, et al. Application and comparison
of fecal microscopic analysis and DNA metabarcoding technology
for dietary analysis of wild boar[J]. Chinese Journal of Wildlife,
2022, 43(3): 645-653.
[8] 滕揚(yáng), 蓋立新, 李建, 等. 兩種分析方法在豹貓食物構(gòu)成中的
應(yīng)用比較[J]. 獸類學(xué)報(bào), 2023, 43(1): 50-58.
TENG Y, GAI L X, LI J, et al. A comparison of two methods of
leopard cat food content analysis[J]. Acta Theriologica Sinica,
2023, 43(1): 50-58.
[9] 中華人民共和國農(nóng)業(yè)部. 農(nóng)業(yè)部關(guān)于印發(fā)《斑海豹保護(hù)行動(dòng)計(jì)
劃(2017—2026年)》的通知[J]. 中華人民共和國農(nóng)業(yè)部公報(bào),
2017(9): 56-62.
Ministry of Agriculture of the People’s Republic of China. Notice
of the Ministry of Agriculture on printing and distributing The Ac?
tion Plan for The Protection of Spotted Seals( 2017-2026)[J]. Gazette
of the Ministry of Agriculture of the People’s Republic of
China, 2017(9): 56-62.
[10] RUGH D J, SHELDEN K E W, WITHROW D E. Spotted
seals, Phoca largha, in Alaska[J]. Marine Fisheries Review,
1997, 59(1): 1-18.
[11] 王者茂, 王丕烈. 斑海豹[M]. 北京: 海洋出版社, 1990: 68-75.
WANG Z M, WANG P L. Spotted seal[M]. Beijing: China
Ocean Press, 1990: 68-75.
[12] 田甲申, 鹿志創(chuàng), 羅耀明, 等. 應(yīng)用碳氮穩(wěn)定同位素研究斑
海豹(Phoca largha)的食性[J]. 生態(tài)學(xué)雜志, 2015, 34(12):
3453-3458.
TIAN J S, LU Z C, LUO Y M, et al. Feeding habits of the spotted
seal Phoca largha by stable carbon and nitrogen isotopes
analysis[J]. Chinese Journal of Ecology, 2015, 34(12): 3453-
3458.
[13] 鄧景耀, 金顯仕. 渤海越冬場漁業(yè)生物資源量和群落結(jié)構(gòu)的
動(dòng)態(tài)特征[J]. 自然資源學(xué)報(bào), 2001, 16(1): 42-46.
DENG J Y, JIN X S. Dynamic characteristics of abundance and
community structure of fishery species in the overwintering
ground of the Bohai Sea[J]. Journal of Natural Resources,
2001, 16(1): 42-46.
[14] MIYA M, SATO Y, FUKUNAGA T, et al. MiFish, a set of universal
PCR primers for metabarcoding environmental DNA from
fishes: detection of more than 230 subtropical marine species
[J]. Royal Society Open Science, 2015, 2(7): 150088.
[15] 李玉龍, 陳百靈, 鮑相渤, 等. 基于DNA宏條形碼技術(shù)的沙氏下
鱵幼魚食性分析[J]. 中國水產(chǎn)科學(xué), 2023, 30(4): 393-405.
LI Y L, CHEN B L, BAO X B, et al. Preliminary dietary analysis
of Hyporhamphus sajori juveniles based on DNA metabarcoding
[J]. Journal of Fishery Sciences of China, 2023, 30(4):
393-405.
[16] LERAY M, MEYER C P, MILLS S C. Metabarcoding dietary
analysis of coral dwelling predatory fish demonstrates the minor
contribution of coral mutualists to their highly partitioned, generalist
diet[J]. PeerJ, 2015, 3: e1047.
[17] 劉剛, 寧宇, 夏曉飛, 等. 高通量測序技術(shù)在野生動(dòng)物食性
分析中的應(yīng)用[J]. 生態(tài)學(xué)報(bào), 2018, 38(9): 3347-3356.
LIU G, NING Y, XIA X F, et al. The application of highthroughput
sequencing technologies to wildlife diet analysis[J].
Acta Ecologica Sinica, 2018, 38(9): 3347-3356.
[18] 李晗溪, 黃雪娜, 李世國, 等. 基于環(huán)境DNA-宏條形碼技術(shù)
的水生生態(tài)系統(tǒng)入侵生物的早期監(jiān)測與預(yù)警[J]. 生物多樣
性, 2019, 27(5): 491-504.
LI H X, HUANG X N, LI S G, et al. Environmental DNA
(eDNA)-metabarcoding-based early monitoring and warning for
invasive species in aquatic ecosystems[J]. Biodiversity Science,
2019, 27(5): 491-504.
[19] BUKHTIYAROV Y A, FROST K J, LOWRY L F. New information
on foods of the spotted seal, Phoca largha, in the Bering
Sea in spring[R]//FAY F H, FEDOSEEV G A. Soviet-American
cooperative research on marine mammal: Vol. 1: pinnipeds.
Washington: NOAA Technical Report NMFS, 1984: 55-59.
[20] 劉修澤, 李玉龍, 王文波, 等. 遼東灣北部海域魚類群落結(jié)
構(gòu)及多樣性[J]. 水產(chǎn)學(xué)報(bào), 2015, 39(8): 1155-1165.
LIU X Z, LI Y L, WANG W B, et al. The fish community structure
and diversity in the north of Liaodong Bay[J]. Journal of
Fisheries of China, 2015, 39(8): 1155-1165.
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
