金釵石斛種子直播幼苗不同月齡共生真菌群落多樣性變化
2024-10-09 00:00:00高世南張文杰王夢雪梁玉瓊邵士成
熱帶作物學報 2024年9期

關鍵詞:金釵石斛;幼苗;ITS 擴增子測序;菌根真菌;種群重建
中圖分類號:S567.239 文獻標志碼:A
蘭科(Orchidaceae)是被子植物中物種數量較為豐富的類群之一,全世界約有870 屬29 524種[1],我國約有181 屬1708 種[2]。蘭科植物的生存和繁衍依賴于專性的共生真菌和傳粉昆蟲[3-4],因此更容易受到氣候變化和人為采集的影響[5-6]。目前所有野生蘭科植物均列入《瀕危野生動植物種國際貿易公約》(CITES),并嚴禁在國際上交易流通[7]。蘭花中不乏有一些具有重要觀賞價值和藥用價值的種類,如石斛屬(Dendrobium)、開唇蘭屬(Anoectochilus)、獨蒜蘭屬(Pleione)等。蘭科植物不僅是植物多樣性保護的旗艦類群,也是科學研究的熱點類群。
蘭科植物的種子大多細若粉塵,缺乏胚乳儲存營養,需要共生真菌為其提供碳源、氮源、水和無機鹽等才能成功萌發[8-9]。共生真菌的菌絲在種子萌發形成的原球莖基部細胞中或成年植株的根皮層細胞內部會纏繞形成菌絲團結構(peloton),這是蘭科菌根的典型特征[10]。蘭科植物在生命伊始就與共生真菌建立了特殊的營養作用關系,二者無論是在生理上還是生態上均存在密切的聯系。與蘭科植物共生的典型真菌類群俗稱“ 蘭科絲核菌” 包括: 蠟殼耳科( Sebacinaceae)、無孢蠟殼科(Serendipitaceae)、膠膜菌科(Tulasnellaceae)和角擔菌科(Ceratobasidiaceae)等[10]。此外,一些外生菌根真菌和腐生真菌也可以作為蘭科植物的共生真菌而存在[11]。蘭科植物在整個生活史中總是部分或完全依賴共生真菌獲取必要的營養,因此探究蘭科植物與共生真菌關系的穩定性及其變化規律,對蘭花的人工栽培和野外回歸具有重要意義。
多數蘭科植物從種子至成年植株的發育過程中,需要先經歷依賴真菌獲取營養的異養階段,然后才逐步開始光合自養。由于營養需求的轉變,共生真菌類群可能會發生變化。VENTRE 等[12]綜合分析了蘭科菌根真菌種類隨宿主發育的轉變模式包括:不轉變、嵌套性增加或減少、部分替換以及完全替換4 種類型。某些蘭科植物終生偏好于某一特定真菌類群,如杏黃兜蘭(Paphiopedilumarmeniacum)從幼苗到開花的發育過程中,僅有膠膜菌科真菌是優勢類群[13];歐洲對葉蘭(Neottia ovata)從原球莖到成年植株階段的主要真菌類群均為無孢蠟殼科[14]。共生真菌在蘭花生命歷程中發生轉變也是常見的模式,藥用植物天麻(Gastrodia elata)種子的萌發依賴紫萁小菇(Mycena osmundicola),而原球莖后續發育需要與蜜環菌(Armillaria mellea)共生[15]。同樣在地生蘭和附生蘭中發現了真菌類群轉變,杜鵑蘭(Cremastra appendiculata)在原球莖階段的優勢真菌屬于鬼傘科(Psathyrellaceae),在成年后增加了角擔菌科和蠟殼耳目(Sebacinales)[16];而風蘭(Vanda falcata)的種子萌發由角擔菌科真菌誘導,成年植株還與膠膜菌科真菌共生[17]。由此可見,蘭科植物在生長發育過程中的共生真菌轉變模式不盡相同,對于特定的蘭花物種往往需要野外驗證,才能深入了解其自然狀態下對共生真菌的真實需求。
依賴有效的促萌發真菌對蘭科植物進行野外種苗擴繁是瀕危物種恢復的有效手段。目前基于種子和共生真菌的人工擴繁主要集中在地生蘭,JIANG 等[18] 針對藥用植物手參( Gymnadeniaconopsea)的野外原地共生萌發試驗證實了角擔菌屬(Ceratobasidium)GS2 菌株能夠在自然條件下促使手參種子成功萌發并形成幼苗;對于具有觀賞價值的紋蝶蘭(Anacamptis papilionacea),膠膜菌科AP1 菌株能夠有效促進其種子在原地條件發育為幼苗[19]。本實驗室前期也利用一株蠟殼耳科JC-01 菌株對瀕危附生蘭金釵石斛(Dendrobiumnobile)開展了適宜自然生境下的原地共生萌發,成功建立了人工種群[20]。然而,這些在自然條件下形成的人工幼苗,其根系真菌的變化模式有待進一步探究,這將有助于理解個體在發育過程中對共生真菌的動態需求以及共生真菌對人工種群維持和變化的影響。
石斛(D. nobile Lindl.),又稱金釵石斛,是石斛屬多年生附生草本植物,因干燥后形態酷似我國古代婦女的發釵而得名[21]。作為我國傳統中藥材,金釵石斛被《中華人民共和國藥典》(2020年版)所收錄。金釵石斛莖中富含石斛堿、多糖和酚類等有機化合物等,經藥理研究證明有降血糖血脂、抗氧化衰老以及抗腫瘤等功能[22]。然而,對金釵石斛的過度利用以及自身繁殖率低等原因導致野生資源瀕危[23],我國2021 年頒布的《國家重點保護野生植物名錄》將其列為國家二級保護植物。本研究所用的金釵石斛幼苗基于“種子-真菌混合播種技術”在茶樹上獲得種苗,并在采集根系的基礎上對比人工擴繁的幼苗與自然形成的幼苗在共生真菌群落組成和多樣性方面的差異,同時也對人工幼苗種群的共生真菌的菌群轉變模式進行探究,以期為金釵石斛的種苗繁育和人工種群生態恢復提供理論依據。
1 材料與方法
1.1 材料
利用種子原地共生萌發技術,于2023年4月在云南省勐臘縣象明鄉茶園(22°13′N,101°20′E,海拔約為1500 m)播種獲得金釵石斛人工幼苗,播種方法參照本實驗室已有的成熟技術[24]進行,利用2 株膠膜菌科真菌(編號為FDL-01、JC-03)、2 株蠟殼耳科真菌(編號為JC-01、YB-01)和4株無孢蠟殼科真菌(編號為LP-01、RJC-131、YB-03 和YDLXB)作為真菌處理組,不添加真菌的作為空白對照組(CK)。于當年8 月、9 月和10 月分別采集真菌處理組120d(4 月齡)、150d(5 月齡)、180d(6 月齡)及CK 的幼苗根樣,每個處理采集3個重復樣本。將收集的根系樣本分裝至無菌采樣袋中并做好標記,置于生物樣品采樣箱中低溫運回實驗室。
直播茶園所在的象明鄉是典型熱帶季風氣候區, 年平均溫度21.9 ℃ , 年降雨量為1500~1900 mm,森林覆蓋率達到64.96%,主要植被類型為熱帶雨林、亞熱帶常綠闊葉林、落葉闊葉林等[25]。茶園中的主要樹種是普洱茶(Camelliasinensis var. assamica),周圍伴有一定的遮蔭喬木。茶園溫暖、濕潤以及一定遮蔭的生態條件適于金釵石斛種子的野外共生萌發。
1.2 方法
1.2.1 根樣處理參考 PANDEY 等[26]的方法對金釵石斛幼苗根樣進行處理,先用流水清洗干凈根上附著的苔蘚等雜質,然后用無菌濾紙吸干水分。隨后用75%乙醇消毒1 min,再用質量體積分數為3%的次氯酸鈉溶液消毒30 s,重復75%乙醇消毒1 min,最后用無菌純水清洗5 次。消毒的根樣用無菌濾紙吸干水分后,刮去根被。將處理的根樣放入無菌離心管并編號,置于液氮中速凍,于–80 ℃冰箱低溫保存。
1.2.2 真菌總 DNA的提取和rDNA-ITS 擴增子測序 金釵石斛幼苗根樣品經冷鏈運輸至上海派森諾生物科技股份有限公司進行真菌總DNA 的提取和rDNA-ITS 擴增子測序。以真菌核糖體DNA的內部轉錄間隔區ITS2 序列為目標,用蘭科菌根真菌特異性引物ITS3(5?–GCATCGATGAAGAACGCAGC–3?)和ITS4OF(5?–GTTACTAGGGGAATCCTTGTT–3?),并添加特異性標簽序列(AAGCACTG)對ITS2 區域進行PCR 擴增[27]。回收后的PCR 產物進行定量與測序文庫的構建,對質量合格的文庫,在NovaSeq-PE250(Illumina,USA)平臺上進行2×250 bp 的雙端測序。
1.3 數據處理
對得到的下機原始序列文件,首先利用QIIME2 軟件( Quantitative Insights Into MicrobialEcology, v2023-7, http://qiime.org/)的cutadapt 插件去除序列的引物片段,然后在R 軟件中用DADA 2 方法對序列進行后續的質量過濾、去噪、拼接和去嵌合體等質控步驟,將序列以100%相似度聚類,得到ASVs(amplicon sequence variants)特征序列,去除序列總數僅為1 的ASVs 后得到ASVs 特征表。使用QIIME 2 軟件以最低樣本量的95%對各樣本進行抽平,預測同一測序深度下觀測到的ASVs 數量及其相對豐度,計算α 多樣性指數并繪制稀疏曲線;使用R 軟件中的Venndiagram 包繪制不同樣本間的Venn 圖,分析不同分組間獨有和共有的ASVs 數量; 基于真菌UNITE 數據庫,利用QIIME 2 軟件的classifysklearn插件對每個ASVs 特征序列進行物種注釋,繪制物種堆積柱狀圖;使用R 軟件中的vegan包和ggplot2 包分別計算Bray-Curtis 距離和繪制主坐標分析圖,結合ANOSIM 相似性分析,分析樣本間群落結構的差異。
不同播種方式比較采用獨立樣本t 檢驗,不同齡期比較采用非參數Kruskal-Wallis 檢驗。
2 結果與分析
2.1 金釵石斛幼苗根樣測序結果
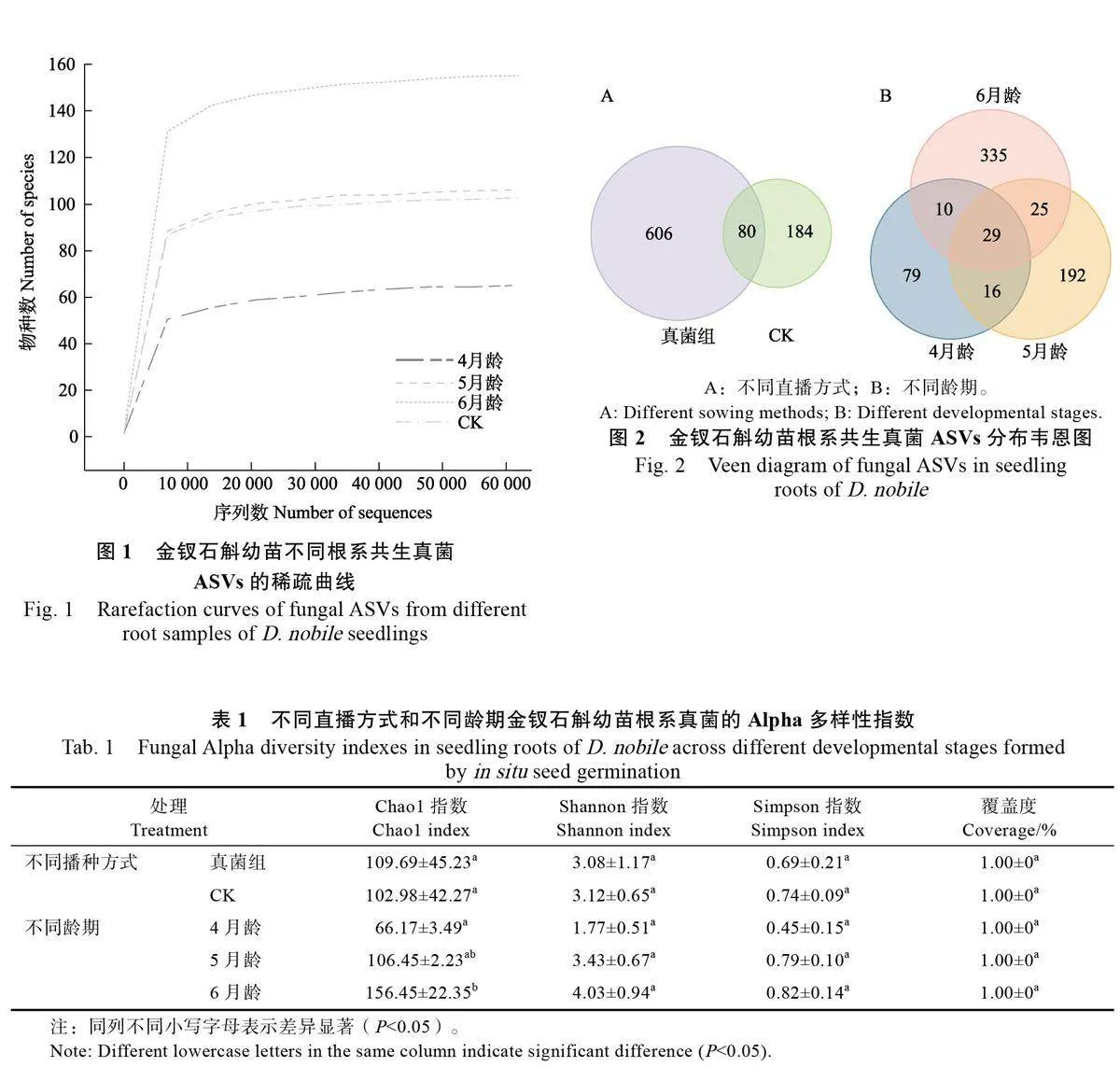
基于Illumina NovaSeq-PE250高通量測序結果,不同金釵石斛幼苗根樣品中的序列長度主要分布在350~400 bp 之間,共獲得1029977條高質量的ITS2序列,金釵石斛4 月齡、5 月齡、6月齡和CK 根中分別獲得287969、232556、274084、235368條有效序列(圖1)。各樣品的ASVs 曲線隨著測序深度的增加而趨于平緩,覆蓋度接近100%,表明本次測序量達到飽和狀態,能夠全面覆蓋幼苗根系的真菌類群。
通過剔除非真菌序列、嵌合體和singleton 序列等質控步驟并結合聚類分析,共得到876 個真菌ASVs。真菌處理組的金釵石斛幼苗根系的真菌ASVs 為686 個,多于CK(264個),二者共有80 個真菌ASVs(圖2A)。金釵石斛4月齡、5月齡和6 月齡幼苗根系的真菌ASVs 總量和獨特的真菌ASVs 數量隨齡期的增長而增加,不同月齡幼苗根系的真菌ASVs 總數分別為134、262和399個,三者共有29個真菌ASVs。金釵石斛6月齡幼苗根系獨特的真菌數量最多,達335 個ASVs(圖2B)。
2.2金釵石斛幼苗根系真菌多樣性
對于不同處理的金釵石斛幼苗根樣,真菌處理組的菌群豐度指數Chao1 指數(109.69±45.23)高于CK(102.98±42.27),即真菌處理組幼苗根系的物種數目多于CK;而真菌處理組的菌群多樣性指數Shannon 指數和Simpson 指數均低于CK,真菌處理組與CK 間3 個多樣性指數均無顯著差異。在不同齡期的金釵石斛幼苗根系中,6月齡幼苗的Chao1 指數(156.45±22.35)顯著高于4月齡幼苗(66.17±3.49)和5 月齡幼苗(106.45±2.23),6 月齡幼苗的Shannon 指數和Simpson 指數同樣也高于其他齡期,但無顯著性差異(表1)。
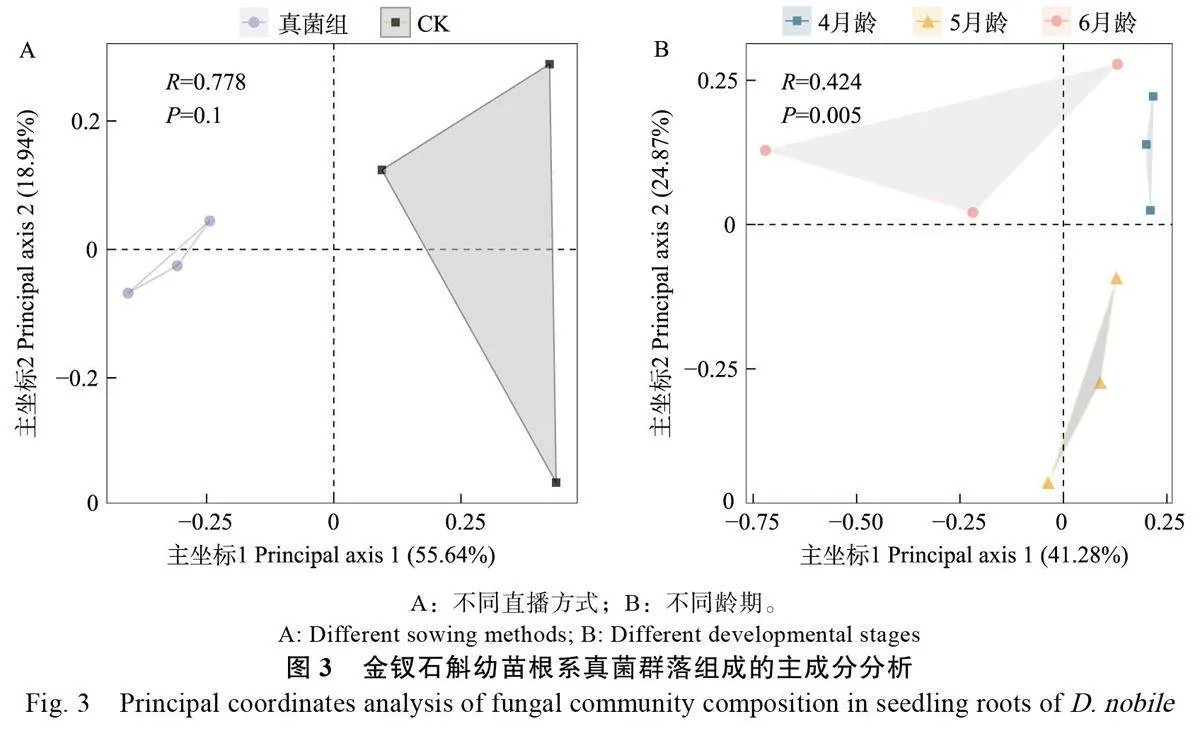
基于Bray-Curtis 距離的主坐標分析對不同處理的金釵石斛幼苗根系真菌群落組成信息進行降維和排序后投射于二維平面上,結果顯示真菌處理組和CK 各聚為一類(圖3A)。根據樣本點的分布情況,真菌處理組各樣本點比CK 更加聚集,結合ANOSIM 非參數檢驗結果,雖然兩組間菌落組成差異較大(R2=0.778),但并無顯著差異。而在不同齡期的幼苗樣品中,4 月齡、5 月齡和6 月齡也各聚為一類,其中6月齡樣本點的分布比其他月齡更加分散(圖3B)。對不同齡期幼苗根系真菌群落結構進行ANOSIM 檢驗表明,金釵石斛幼苗不同齡期之間的根系真菌群落組成具有一定差異(R2=0.424),且差異顯著(P=0.005)。
2.3 金釵石斛幼苗根系真菌物種組成
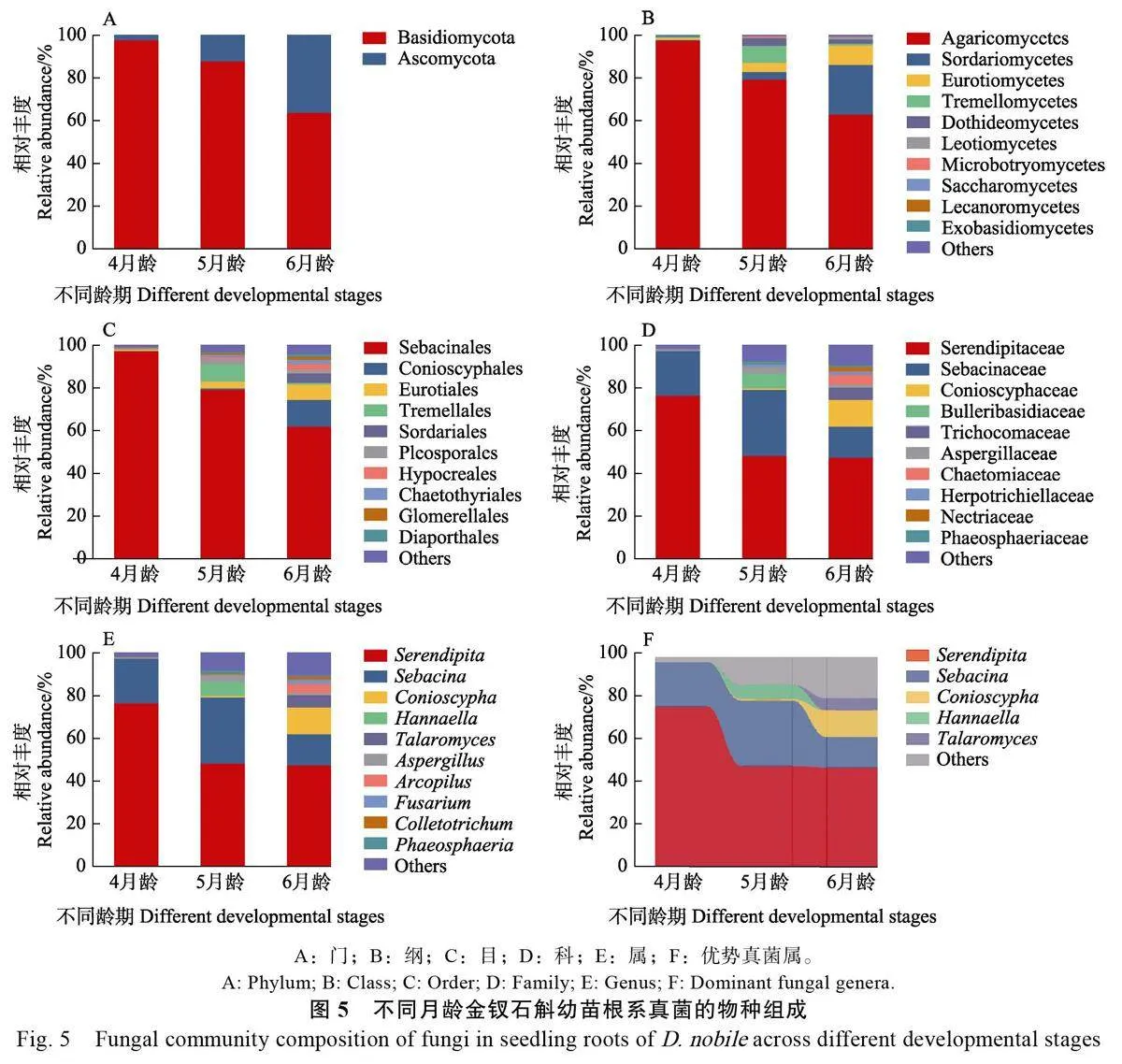
2.3.1 不同直播方式 從真菌處理組和CK 的幼苗根樣品中獲得870 個真菌ASVs,隸屬于2 門11 綱28 目46 科56 屬,兩組之間根系真菌群落的種類組成和相對豐度有一定差異。在門水平,真菌處理組和CK 的根系真菌均由擔子菌門(Basidiomycota)和子囊菌門(Ascomycota)組成,兩類真菌的相對豐度類似,擔子菌門分別為83.29%和83.93%,子囊菌門分別為16.71%和16.07%(圖4A)。
在綱水平,真菌的主要綱(相對豐度大于1%)包括: 傘菌綱( Agaricomycetes ) 、糞殼菌綱(Sordariomycetes)、座囊菌綱(Dothideomycetes)、散囊菌綱(Eurotiomycetes)、銀耳綱(Tremellomycetes)和錘舌菌綱(Leotiomycetes)。傘菌綱是真菌處理組和CK 的優勢綱,相對豐度分別達80.04%和83.75%。其余各綱的相對豐度均低于10%,其中糞殼菌綱在真菌處理組的相對豐度較高,達9.15%,而座囊菌綱在CK 的豐度達7.80%(圖4B)。
在目水平,主要類群(相對豐度大于2%)有:蠟殼耳目、雞油菌目(Cantharellales)、格孢腔菌目(Pleosporales)、Conioscyphales、刺盾炱目(Chaetothyriales)、曲霉目(Eurotiales)和銀耳目(Tremellales)。蠟殼耳目是真菌處理組和CK的優勢目,相對豐度分別為79.71%和67.52%。對于真菌處理組而言,其余各目的相對豐度均低于5%,而在CK 中,雞油菌目和格孢腔菌目的相對豐度分別為16.22%和6.74%(圖4C)。
在科水平,主要類群(相對豐度大于2%)包括: 無孢蠟殼科、蠟殼耳科、膠膜菌科、Conioscyphaceae、Herpotrichiellaceae 和大皰擔子科(Bulleribasidiaceae)。無孢蠟殼科是真菌處理組和CK 的優勢科,相對豐度分別達57.52%和55.55%。蠟殼耳科在兩組中的相對豐度占比均較高,分別為22.19%和11.98%。膠膜菌科僅出現在CK 中,相對豐度達16.22%。其余各科的相對豐度在兩組中均低于5%(圖4D)。
在屬水平,真菌的主要屬(相對豐度大于2%)有: 無孢蠟殼屬( Serendipita ) 、蠟殼耳屬(Sebacina)、膠膜菌屬(Tulasnella)、Conioscypha、Pyrenochaetopsis、Hannaella 和Medicopsis。無孢蠟殼屬和蠟殼耳屬是真菌處理組的優勢屬,相對豐度分別達57.52%和22.19%,而CK 的優勢屬不僅包括無孢蠟殼屬和蠟殼耳屬,還包括膠膜菌屬,相對豐度分別為55.55%、11.98%和16.22%。其余各屬的相對豐度均低于5%(圖4E)。此外,針對相對豐度大于5%的不同真菌屬進行組間差異分析,結果顯示無孢蠟殼屬在兩組間的差異不大,而真菌處理組的蠟殼耳屬相對豐度比CK 高10.21%。膠膜菌屬僅出現在CK,相對豐度比蠟殼耳屬高4.24%,是CK 幼苗根系的優勢屬之一(圖4F)。
2.3.2 不同齡期 從真菌處理組不同齡期幼苗根樣品中獲得686 個真菌ASVs,隸屬于2 門11 綱28 目46 科56 屬,不同齡期幼苗根系真菌群落的種類組成和相對豐度有一定差異。在門水平,各齡期幼苗根系真菌均由擔子菌科和子囊菌科組成,隨著培養齡期的增加,擔子菌門的相對豐度逐漸下降,而子囊菌門則逐漸增加(圖5A)。
在綱水平,傘菌綱(97.62%)是4 月齡幼苗的主要真菌類群,其相對豐度隨幼苗齡期的增長而減少,其余綱的相對豐度均小于1%;5月齡幼苗根中的糞殼菌綱、散囊菌綱和銀耳綱的相對豐度比4 月齡分別增加至3.56%、4.35%和7.93%;6月齡幼苗根中的糞殼菌綱和散囊菌綱的相對豐度則分別增至23.38%和9.09%,而銀耳綱的相對豐度降至0.72%(圖5B)。
在目水平,4月齡幼苗根系的主要目是蠟殼耳目(相對豐度為97.61%),其余目的相對豐度均小于1%。除蠟殼耳目外,5月齡幼苗根中相對豐度較高的真菌類群為銀耳目(7.94%),但6月齡幼苗根中的銀耳目相對豐度卻降為0.72%,而Conioscyphales 和曲霉目的相對豐度則分別增至12.65%和7.12%(圖5C)。
在科水平,無孢蠟殼科和蠟殼耳科是4 月齡幼苗根系的主要真菌類群, 相對豐度分別為76.55%和21.06%,其余真菌科的相對豐度低于1%。5 月齡幼苗的主要真菌科為無孢蠟殼科、蠟殼耳科和大皰擔子科,而6 月齡幼苗的主要真菌科基于上一階段還增加了發菌科(Trichocomaceae,5.70%)和毛殼科(Chaetomiaceae, 4.49%)(圖5D)。
在屬水平,無孢蠟殼屬和蠟殼耳屬占據4 月齡幼苗根系主要地位,總體相對豐度高達97.61%。5 月齡幼苗根系的主要真菌類群在4 月齡幼苗的基礎上增加了Hannaella ( 6.75% ) 和曲霉屬(Aspergillus, 3.11%),而6 月齡幼苗根中優勢真菌轉變為Conioscypha ( 12.65%) 、籃狀菌屬(Talaromyces, 5.71%)和Arcopilus(4.15%)(圖5E)。此外,針對相對豐度大于5%的真菌屬進行組間差異分析,結果顯示無孢蠟殼屬從4 齡期幼苗轉變成5 齡期幼苗過程中相對豐度減少了28.11%,而在后續發育中趨于穩定;蠟殼耳屬從4 月齡到6 月齡的相對豐度呈先增后降的趨勢;Hannaella 是5 月齡幼苗的獨特優勢真菌,相對豐度達6.75%,該真菌類群在其他齡期幼苗根中的相對豐度小于1%;Conioscypha 和籃狀菌屬的相對豐度隨幼苗的生長而增加,在6 月齡幼苗中的相對豐度分別達到12.65%和5.71%(圖5F)。
3討論
本研究表明金釵石斛種子原地共生萌發不同處理方式形成的幼苗根系在真菌群落組成和相對豐度方面具有一定差異,真菌處理組和CK 的優勢真菌類群有重疊,但CK 能夠招募到環境中的膠膜菌科真菌并作為優勢類群而存在。不同齡期幼苗根系的菌群結構差異顯著,真菌的豐富度和多樣性隨幼苗生長而增加,典型蘭科共生真菌的相對豐度隨著月齡的增加而下降,同時非共生真菌的占比增加,且種類和豐度具有齡期特異性。
金釵石斛原地共生萌發形成的幼苗和自然條件(CK)形成的幼苗中,二者的真菌ASVs 數量雖然差別較大,但真菌群落的豐富度和多樣性無顯著差異,表明在同質的環境下,金釵石斛幼苗偏好性地選擇相似的共生真菌作為主要類群,如蠟殼耳目。某些蘭科植物也表現出在相似的生境下選擇相同的真菌類群的現象。在大陸尺度上,分布于亞洲的不同手參種群偏好的共生真菌均屬于角擔菌科[28];而生長在同一個島嶼上的琉球石斛(D. okinawense)均傾向于選擇同一株膠膜菌科真菌TU11 作為主要共生菌[29];分布于相似沼澤生境的5 個幽靈蘭(D. lindenii)種群,其根系的優勢共生真菌均為角擔菌科真菌[30]。雖然蘭科共生真菌是廣泛分布的,但某些真菌在局部特殊生境中相對較高的豐度可能使同一環境內的蘭花個體對共生真菌的選擇更加局限和集中。
在本研究中,金釵石斛幼苗根系真菌的多樣性和豐度隨著齡期的增長而逐漸增加,同時獨特的ASVs 數量也隨之增加,從4 月齡幼苗的79 個ASVs 增加到6 月齡幼苗的355 個ASVs。當幼苗逐漸適應環境而存活下來,環境中的機會性真菌(opportunistic fungi)勢必會占據根中的一席之地。這一現象也常見于其他蘭花類群,如黃花白及(Bletilla ochracea)從生長期到花期的發育過程中,真菌OTUs 從119 個增加到270 個[31];在哥斯達黎加熱帶雨林共存的4 種附生蘭,從幼年階段到成年階段的發育過程中其根系的核心真菌類群會保留,同時增加了新的真菌類群[32]。但這種規律不能一概而論,對杏黃兜蘭生長過程中的真菌組成研究表明,杏黃兜蘭從幼苗的營養生長轉變到成年植株的生殖生長期間,根系的真菌OTUs 數量從幼苗時期的126 個降至開花時期的67 個[13];針對風蘭的菌群變化的研究則顯示,幼苗和成年植株的真菌群落組成保持相對穩定,子囊菌門占比超過一半,其次為擔子菌門,其中典型的蘭科共生菌僅占少部分[33]。不同生活型的蘭花在生長期間復雜的營養需求可能會導致真菌類群組成的變化,從而影響特定時期的真菌豐度差異。
在本研究的金釵石斛幼苗根系的真菌組成中,典型的蘭科植物共生真菌占據大部分,在真菌處理組種子原地共生萌發形成的幼苗中主要由無孢蠟殼屬和蠟殼耳屬組成,而在CK 組形成的幼苗中,膠膜菌屬同樣成為了優勢真菌。金釵石斛的共生真菌種類豐富,早期從金釵石斛根中分離到角擔菌屬和蘭小菇(Mycena orchidicola),被認為是潛在的共生真菌[34],這已在鐵皮石斛(D.officinale)的種子共生萌發試驗中得到證實[35]。本研究中,CK 的幼苗能夠招募到膠膜菌屬真菌,暗示周圍環境中有該真菌類群的存在和分布,但真菌處理組幼苗根系中該類群的相對豐度僅為0.02%,低于CK(16.22%),這可能是因為人工添加的真菌在野外播種點建立了局部優勢并與環境中的真菌彼此具有拮抗作用,導致真菌處理組根系中膠膜菌科真菌的相對豐度極低。然而蘭科共生真菌之間的拮抗關系驗證試驗主要集中在實驗室條件下,與單獨菌株的種子共生萌發相比,不同共生真菌的疊加培養會導致金釵石斛和紋蝶蘭的種子萌發率下降[36-37],本研究結果也初步表明共生真菌的拮抗作用可能在金釵石斛種子原地共生萌發中存在。
金釵石斛幼苗根系真菌組成并非一成不變,隨著齡期的增長(4 月齡至6 月齡),優勢真菌的占比逐漸下降,而機會性真菌的相對豐度逐漸增加。在優勢真菌屬中,無孢蠟殼屬的相對豐度先下降后趨于穩定,而蠟殼耳屬的相對豐度則先增后降,這反映出金釵石斛幼苗隨著個體發育變化對共生真菌的偏好性發生了改變。這種偏好性變化可能與蘭科植物在不同生長時期的營養需求有關,一項針對不同營養型鳥巢蘭屬(Neottiaspp.)蘭花的研究發現,能夠進行光合作用的類群通常在發育早期與無孢蠟殼科真菌建立共生關系,而腐生的類群則經常與蠟殼耳科真菌共生[14]。推測其原因可能是無孢蠟殼科真菌僅能夠支持蘭科植物早期真菌異養萌發時期的營養需求,在幼苗逐漸生根的過程中對營養需求逐漸增大,不足以支持后續的大量營養需求[38]。而蠟殼耳科是腐生蘭[如鳥巢蘭(Neottia nidus-avis)]的主要共生真菌類群,因其同時具有形成外生菌根的能力,很可能從周圍樹木中獲取碳源,從而支持腐生蘭完成其生活史[39],據此推測5 月齡幼苗根系中蠟殼耳屬真菌增加的原因是該類群具有更加高效的營養供應能力。此外,5 月齡幼苗根系特有的優勢真菌屬是Hannaella,據報道是禾本科(Poaceae)植物內生酵母菌能夠產生植物激素吲哚乙酸(IAA),促進植物的生長發育,是潛在的金釵石斛內生真菌資源[40]。6 月齡幼苗的優勢真菌屬為Conioscypha 和籃狀菌屬,前者主要從沉水腐爛的木屑中分離得到[41],可能是茶園環境中廣泛存在的機會性真菌;后者則是青霉菌屬(Penicillium)的有性世代,是典型的植物內生真菌,在幫助幼苗抵抗病原微生物方面可能具有重要作用[42]。
本研究明確了金釵石斛人工形成的幼苗和自然形成的幼苗根系在共生真菌和內生真菌的多樣性和真菌群落組成上的差異,以及不同齡期人工幼苗根系的共生真菌及內生真菌相對豐度的變化模式。在后期有必要對金釵石斛幼苗根系中潛在的有功能性的共生真菌和內生真菌進行分離和驗證,將具有生態功能的菌株應用到金釵石斛的野外回歸中,維持人工幼苗群落穩定,實現該瀕危物種的生態恢復。
致謝 感謝云南省林業和草原科學院研究員蔣宏和深圳市蘭科植物保護和研究中心資源管理部副部長孔德敏惠贈金釵石斛種子。感謝曲靖師范學院Don NalinNilusha Wijayawardene 教授對英文摘要的修改。感謝普洱學院實習生龍蘭和王汶倩以及租車師傅王玉明在野外直播試驗中的幫助。
