忽地笑的花部特征與蝴蝶翅膀傳粉研究
2024-10-17 00:00:00廖瑞凌鑫煜吳遠會朱興福
廣西植物 2024年9期


摘 要: 忽地笑(Lycoris aurea)為石蒜科石蒜屬多年生草本植物,具有重要的藥用和觀賞價值,其花呈蜘蛛花型,具有高度外露的柱頭和花藥,可能代表了一種獨特的傳粉方式。為了探究這種傳粉方式,該文對忽地笑的花部特征、繁育系統和傳粉昆蟲及其行為進行了研究。結果表明:(1)忽地笑的單花持續時間為 (5.9±0.1) d,花粉胚珠比(P/O)約為2.3萬;單花展示存在雌雄異位現象,在開花前期花柱和花絲是分開的,兩者之間的角度約為30°,在開花的中后期花柱逐漸向花絲移動,最終兩者在同一條線上,但柱頭比花藥高2~3 cm;柱頭在整個花期都具有可授性。(2)繁育系統試驗表明,忽地笑具有一定的自交親和能力,但不具有主動自交能力,需要訪花昆蟲來傳粉;補充花粉顯著提高了坐果率,證明存在花粉限制。(3)4種鳳蝶是有效傳粉者,由于忽地笑存在雌雄異位,在開花前期,鳳蝶訪花后翅可能接觸到扁平的花藥,在開花中后期鳳蝶訪花時后翅才可能碰觸到柱頭從而完成傳粉過程。總體而言,該研究證明了忽地笑獨特的花部特征可能與蝴蝶翅膀傳粉相適應,同時也為忽地笑的保護和利用奠定了基礎。
關鍵詞: 忽地笑, 繁育系統, 傳粉限制, 訪花昆蟲, 自交親和性
中圖分類號: Q945.5
文獻標識碼: A
文章編號: 1000-3142(2024)09-1746-09
Floral characteristics and butterfly wing
pollination of Lycoris aurea
Abstract: Lycoris aurea is a perennial herbaceous plant with important medicinal and ornamental values in the genus Lycoris Herb, the family Amaryllidaceae. The flower has a spider shape with highly exserted stigmas and anthers, which may represent a unique pollination type. In order to explore this type of pollination, we studied its floral characteristics, breeding system, and floral visitors and their behaviors. The results were as follows: (1) The single flower lasted (5.9±0.1) d and the pollen-ovule ratio (P/O) reached about 23 000. The flowers showed reverse herkogamy. On the beginning of flowering, the style and filaments were separated and the angle between them was about 30°. The style gradually moved towards the filaments in the middle and later stages and were finally on the same line with the stigma 2-3 cm higher than the anthers. The stigmas were receptive throughout the flowering period. (2) The breeding system test was self-compatible and bagged flowers cannot set fruit, indicating that pollinators were necessary for pollen transfer. Supplemental hand pollination significantly increased fruit production, indicating the existence of pollen limitation. (3) Four species of swallowtail butterfly were the effective pollinators. Due to the existence of herkogamy in L. aurea, on the beginning of flowering, their hindwings were likely to come into contact with the flattened anthers. Their hindwings might touch the stigma in the middle and later stages and thus the process of pollen transfer was accomplished. In conclusion, this study shows that the uqique floral characteristics of L. aurea are adapted for butterfly wing pollination and also will lay the foundation for its conservation and sustainable utilization.
Key words: Lycoris aurea, breeding system, pollination limitation, flower-visiting insect, self-compatibility
在現存的 35 萬種被子植物中,大約 78%的溫帶物種和 94%的熱帶物種需要動物傳粉(Paton et al., 2008;Ollerton et al., 2011),尤其是昆蟲。最為常見和高效的傳粉昆蟲是蜂類,對很多蜂類來說,花蜜和花粉是它們唯一的食物來源,只有高頻率的拜訪花朵,它們才能獲取足夠的營養用于給自身提供動力和撫育后代,在長期的協同進化后,蜂類進化出和傳粉相適應的一系列特征,比如體表有大量的絨毛(Harder, 1990;Pauw, 2006;Huang & Shi, 2013)。蝴蝶也是常見的訪花昆蟲,訪花主要是為了獲取少量的能量用于生存,因為它們用于生長和繁殖所需的營養物質在幼蟲階段都已經積累完成(周堯,1998;廖宇,2009),蝴蝶有著長長的口器,可以吸食多種植物的花蜜,這些植物不僅可以是開放式的花朵,還可以是花蜜比較隱藏的花朵,如花冠管較長且為蜂類傳粉的植物,因此蝴蝶被認為是一個傳粉效率低下的機會主義者(Jennersten, 1984;Massinga et al., 2005)。但是部分研究發現蝴蝶也是十分高效的傳粉昆蟲,而且植物也能很好地適應蝴蝶傳粉(Butler & Johnson, 2020;Liu et al., 2022)。考慮到蝴蝶訪花的普遍性,我們需要更多的研究來闡明蝴蝶在傳粉系統中的作用。
大部分由蝴蝶傳粉的植物是由蝴蝶的口器或者頭部來傳播花粉,這些植物往往具高腳碟狀花冠,花冠的下部為狹圓筒狀,上部突然擴大成平面的裂片,花藥和柱頭一般內藏于花冠筒中或者和冠口齊平(Massinga et al., 2005;Ferrero et al., 2009)。為了減少同域分布的其他植物花粉帶來的干擾,不同植物趨向于使用傳粉動物的各部位來傳粉(Dressler, 1968;Armbruster et al., 1994;Muchhala & Potts, 2007;Johnson, 2010)。雖然蝴蝶翅膀的表面積很大,但是相對而言,利用蝴蝶翅膀傳粉的植物較少,這些植物主要分布在8個不同的科中,包括石蒜科、百合科、秋水仙科、鳶尾科、唇形科、豆科、杜鵑花科和山柑科,在這些科中,植物的花通常具有高度外露的花藥和柱頭,花冠成蜘蛛花型或者碗狀(Cruden & Hermann-Parker, 1979;Johnson & Bond, 1994;Butler & Johnson, 2020;Liu et al., 2022)。因此,該種花部特征可能代表了蝴蝶翅膀傳粉綜合征。
忽地笑(Lycoris aurea)為石蒜科(Amaryllidaceae)石蒜屬(Lycoris Herb)多年生草本植物(裴鑒,1985)。忽地笑鱗莖內富含石蒜堿、加蘭他敏、力克拉敏等10余種生物堿,可用于治療輕中度老年性癡呆癥、小兒麻痹后遺癥等疾病,同時具有抗腫瘤、抗病毒和抗瘧的作用,對癌癥具有一定療效,不僅能抑制癌細胞的生長,還能殺死癌細胞,具有重要的藥用價值(孫志峰等,2013;季宇彬等,2016;Masi et al., 2020);忽地笑花色艷麗、色彩繽紛,是一種具有觀賞價值的野生花卉資源,深受人們的喜愛,在日本和中國臺灣地區的忽地笑切花市場已經初具規模(袁菊紅和胡綿好,2009)。近年來,由于經濟利益驅動,國內的忽地笑被大量采挖,野生資源破壞較為嚴重(潘春屏,2023)。為減緩忽地笑這一特有花卉物種資源的流失和破壞,保護其資源多樣性,有必要對忽地笑的野外繁殖狀況進行研究。在石蒜科中發現蝴蝶翅膀傳粉的植物,如網球花(Scadoxus multiflorus)、美麗花盞(Brunsvigia marginata)、君子蘭(Clivia miniata)、紫根蘭(Cyrtanthus elatus)等都是分布在南非(Butler & Johnson, 2020),它們都具有典型蝴蝶翅膀傳粉的花部特征,而分布于中國的忽地笑有類似的特征,呈蜘蛛花型,具有高度外露的柱頭和花藥,我們預測其也是由蝴蝶翅膀傳粉。為此,本文對該物種花部特征、繁育系統、傳粉昆蟲及其行為進行了研究,旨在探討以下兩個問題:(1)蝴蝶是否為忽地笑的有效傳粉動物和如何為其傳粉;(2)忽地笑的自然居群是否存在花粉限制。這些問題的答案不僅可以讓我們更深入認識蝴蝶翅膀傳粉綜合征的進化,也可為忽地笑的資源保護和利用奠定基礎。
1 材料與方法
1.1 試驗地點和試驗材料
觀察試驗于2022年8—10月在貴州省貴陽市花溪區黨武鎮思丫村附近的喀斯特落葉林下進行,地理位置為106° 38′ E、26° 23′ N,海拔1 130~1 220 m,屬中亞熱帶溫潤季風氣候,全年平均氣溫15.7 ℃,植被類型為落葉闊葉林。忽地笑主要分布在中國西南的省份,貴陽市處于該物種的分布中心(裴鑒,1985),此處是開展忽地笑的傳粉生物學的理想試驗區。忽地笑鱗莖卵形;葉劍形;傘形花序,有花4~8朵,黃色;蒴果具3棱;種子少數,近球形,黑色;花葉不同期,葉期10月至次年5月,花期8—9月,果期10月。忽地笑在此處生長在腐殖土和石縫中,研究的居群大約有3 000個個體,400~600個花序。
1.2 方法
1.2.1 花部特征觀察與花粉胚珠比(P/O)測定 隨機選擇20朵來自不同植株上的花蕾,用號牌進行標記。于每天早上9:00進行觀察并記錄花的狀態,即花瓣開放狀態、花藥是否開裂散粉。我們觀察到忽地笑的花柱存在運動現象后,記錄了花柱的運動方向并測量了花絲和花柱兩者之間的角度。從開花第一天到花朵枯萎凋謝,我們用量角器測量并記錄花柱與和花柱夾角最小的一束花絲之間的度數,用游標卡尺測量了花藥與柱頭之間的距離。隨機選擇20個處于盛花期的植株,用號牌在每個植株上隨機標記1朵開放的花,用游標卡尺測量花冠長、花冠寬和花冠筒長。
隨機選取35朵來源于不同植株上即將開放的花蕾,用鑷子剖開花蕾后取出全部花藥分別置于1.5 mL離心管中,用鑷子夾取子房置于另外一個離心管中,同一花蕾的花藥和子房標號對應,加入75%酒精,帶回實驗室。在室內用鑷子分別將每朵花的花藥全部夾碎,置于50 mL EP管中,加蒸餾水定容至50 mL,充分搖勻后用移液槍取出10 μL,置于載玻片上,于顯微鏡下計數單花花粉數,重復統計10次。用鑷子分別將每朵花的子房取出后剖開,統計單花胚珠數目。計算花粉胚珠比(單花花粉數/單花胚珠數,即P/O)。
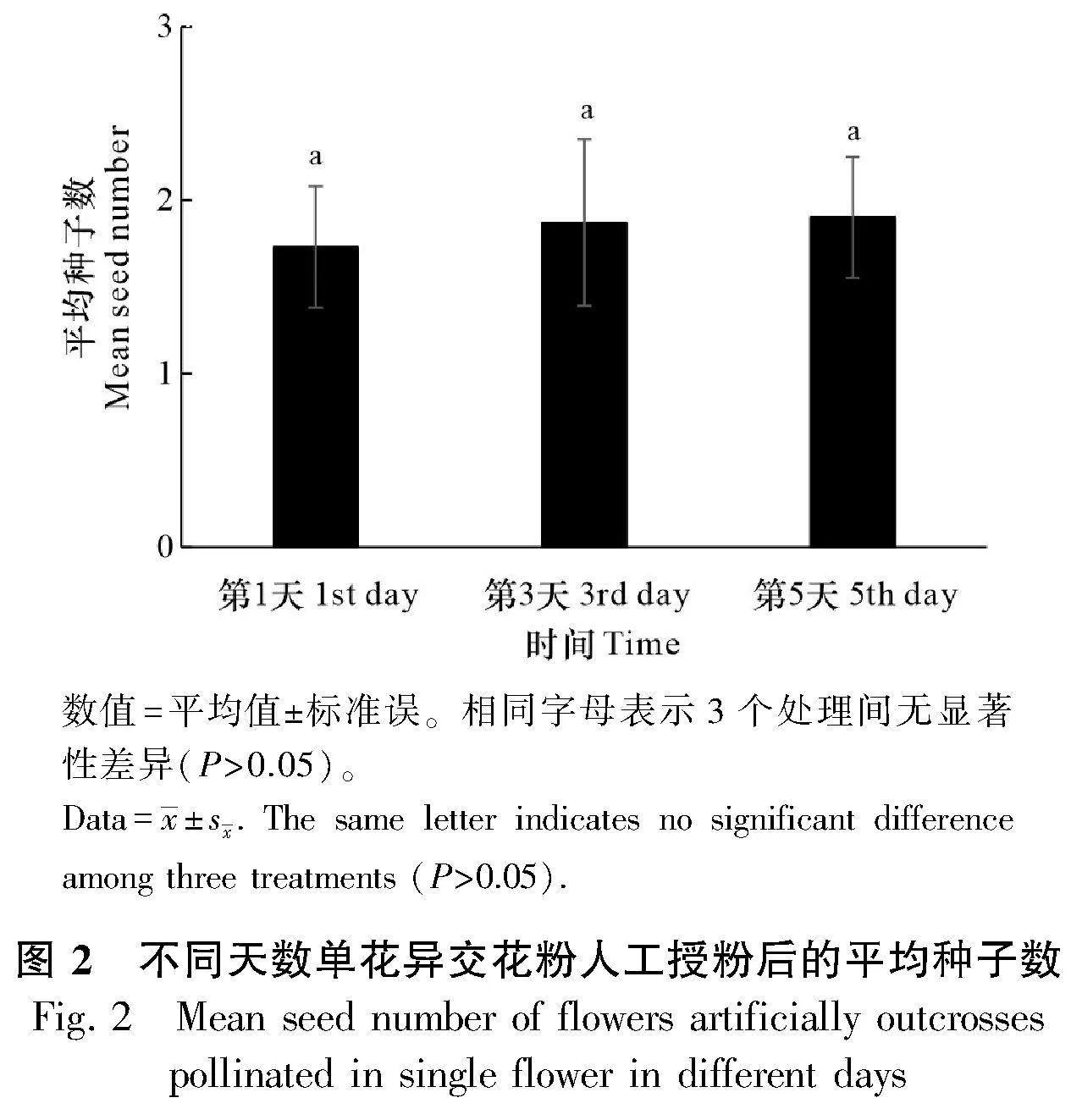
1.2.2 柱頭可授性 隨機選取90朵尚未開放的花蕾套袋,分成3個處理,每個處理30朵花,分別在單花開花第1天、第3天、第5天后取同種異株上剛散粉的花藥對柱頭進行人工授粉。一個月后收取所有處理的花,統計每種處理的平均種子數。實驗結果用SPSS 26.0軟件進行單因素方差分析(one-way ANOVA)。
1.2.3 花蜜量和花蜜糖濃度測定 隨機選取60朵尚未開放的花蕾套袋,分成3個處理,每個處理20朵花,分別在單花開花第1天、第3天、第5天后取下花朵,使用內徑為0.5 mm,長度為100 mm的玻璃毛細管吸取每朵花的花蜜,用游標卡尺測量花蜜在毛細管內的長度,計算花蜜體積公式:V=πr2h(式中:r為毛細管內半徑;h為毛細管內花蜜長度)。將測完體積的花蜜采用手持式折光測糖儀(0~50%、45%~80%)測定糖濃度。花蜜量和花蜜糖濃度分別用SPSS 26.0軟件進行單因素方差分析(one-way ANOVA)。
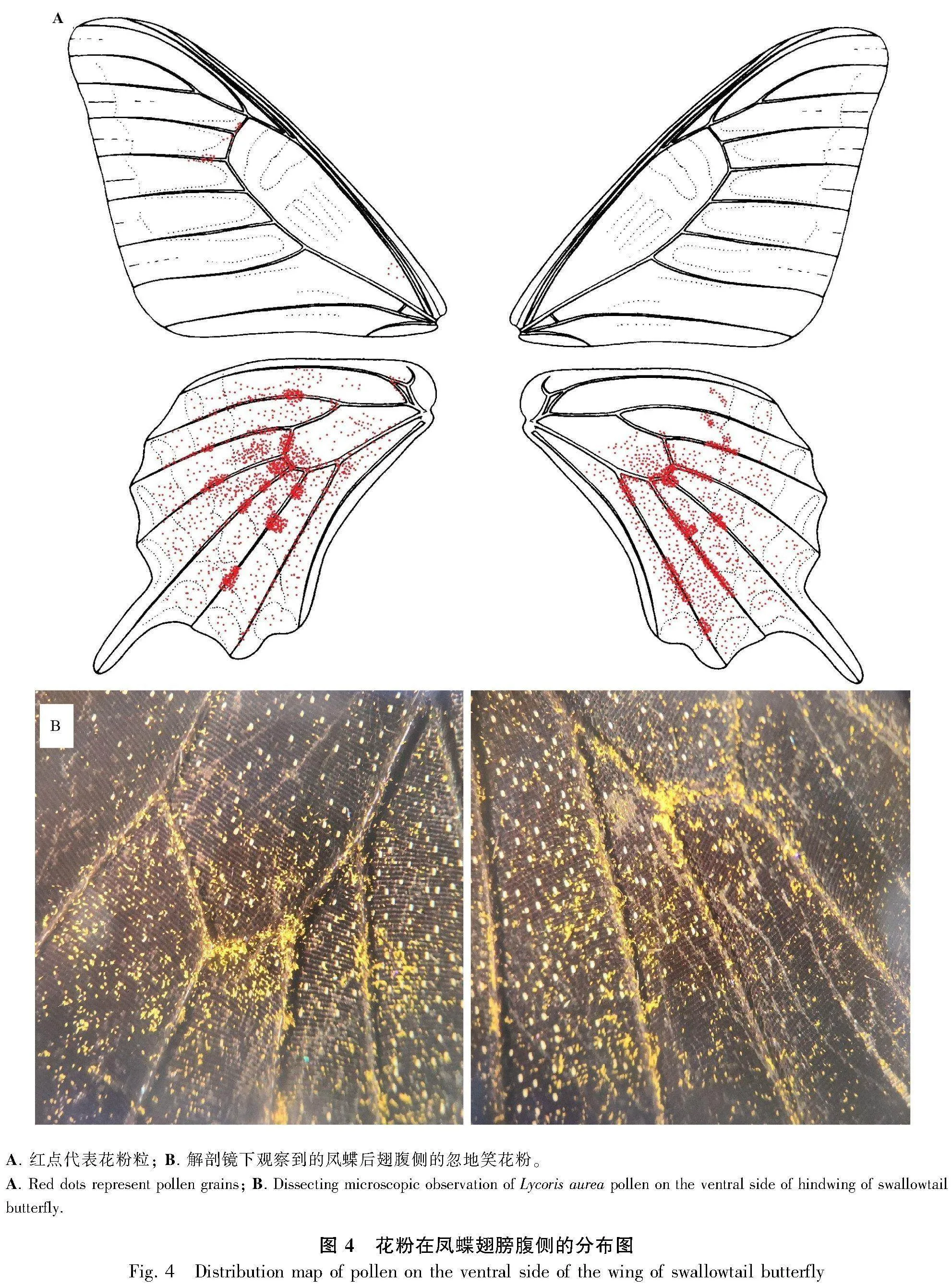
1.2.4 訪花和傳粉昆蟲種類和行為的觀察 于忽地笑盛花期(8月中旬),選擇開花集中的地方,選取200朵花進行訪花昆蟲觀察,連續觀察4 d,時間為9:00—17:00。每隔 1 h 觀察記錄訪花昆蟲種類、數目和訪問頻率,并記錄昆蟲在單花上的訪問時間,對每種昆蟲的訪花行為進行拍照和視頻記錄。用捕蟲網捕捉訪花昆蟲帶回室內鑒定,在解剖鏡下觀察記錄昆蟲攜帶花粉情況,同時在蝴蝶翅脈圖上繪制出花粉所分布的區域。
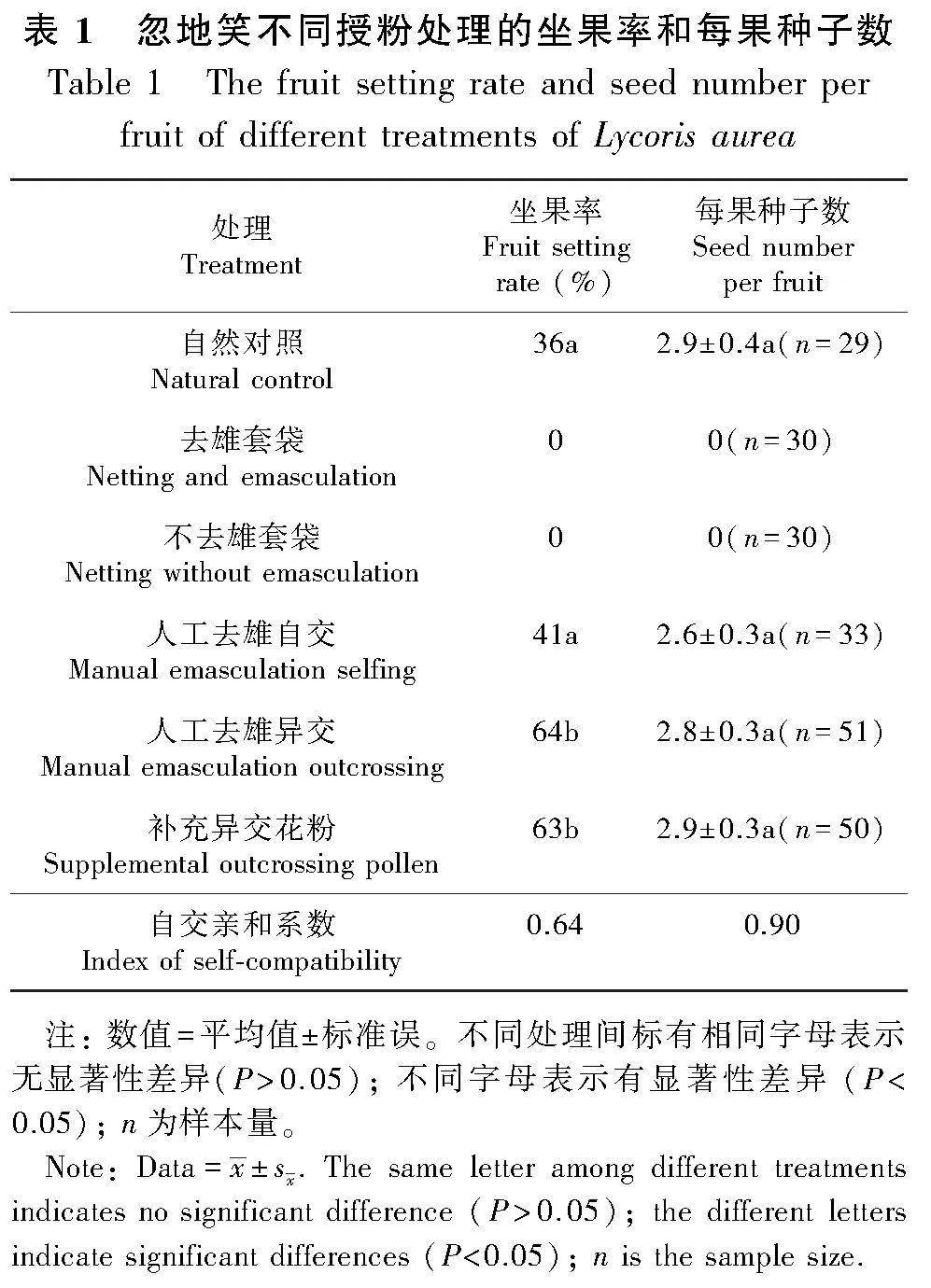
1.2.5 授粉試驗 為檢測忽地笑的繁育系統類型,在該居群實施以下6組處理:(1)自然對照,使用號牌標記80朵即將開放的花蕾,以檢測自然狀態下結實情況;(2)去雄套袋,標記30朵即將開放的花蕾,用鑷子去掉雄蕊后套上尼龍袋,不做任何處理,以檢測是否存在孤雌生殖現象;(3)不去雄套袋,標記30朵即將開放的花蕾并套袋,以檢測是否具備主動自交能力;(4)人工去雄自交,標記80朵即將開放的花蕾去雄并套袋,花開放后用同一植株上其他花朵上的花粉人工授粉;(5)人工去雄異交,標記80朵即將開放的花蕾去雄并套袋,花蕾散開后用其他個體的花粉人工授粉;(6)補充異交花粉,標記80朵花做補充異交花粉處理。一個月后收取所有處理的花,統計坐果率和每果種子數。用人工去雄自交的坐果率和每果種子數分別除以人工去雄異交的坐果率和每果種子數,得出忽地笑的自交親和系數(Becerra & Lloyd, 1992)。比較自然對照和補充授粉處理的坐果率和每果種子數,檢驗該居群是否存在花粉限制。利用卡平方(χ2)和單因素方差分析分別檢驗各種處理的坐果率和每果種子數。
2 結果與分析
2.1 花部特征與P/O 值
忽地笑的單花持續時間為(5.9±0.1) d (平均值±標準誤,下同)。由圖1可知,忽地笑最初花藥和柱頭聚集在花朵中央,花蕾上的花瓣散開后雄蕊稍微散開,此時花絲和花柱集中成一束,隨后花柱很快向下移動,同時花藥開始縱裂散粉,開花第2天后花柱逐漸向上移動,第3天時基本與花絲在同一條線上,柱頭比花藥高29.4 mm,第3天后花柱和花絲位置無明顯變化。我們在開花第1天早上9:00觀察到花絲和花柱兩者之間的角度為(30.3±3.2)°,第2天為(19.9±3.1)°,第3天為(3.1±0.3)°,第3天后無明顯變化。花冠長為(50.5±0.9) mm,花冠寬為(86.3±2.5) mm,花冠筒長為(15.0±0.6) mm。單花花粉數目為(310 000.0±19 725.2),胚珠數目為(13.5±0.7),P/O值為(23 488.6±1 306.1)。
2.2 柱頭可授性
進行人工授粉處理后,柱頭在開花后的第1天、第3天和第5天都有一定的可授性,三者產生的種子數不存在顯著差異(df=2, F=0.049, P=0.952)(圖2),這表明忽地笑在整個花期柱頭都具有一定程度的可授性。
2.3 花蜜量和花蜜糖濃度
忽地笑開花第1天的花蜜量為(9.1±0.5) μL,與第3天(19.9±1.6) μL和第5天(21.4±1.1) μL存在顯著性差異(df=2, F=34.768, P<0.05),第3天(19.9±1.6) μL與第5天(21.4±1.1) μL不存在顯著性差異(P=0.351)。第5天的糖濃度為(42.3±2.2)%,與第1天(31.7±1.2)%和第3天(32.0±1.3)%存在顯著性差異(df =2, F=13.464, P<0.05),第1天(31.7±1.2)%與第3天(32.0±1.3)%不存在顯著性差異(P=0.907)(圖3)。
2.4 訪花和傳粉昆蟲種類和行為
對忽地笑的訪花昆蟲連續觀察4 d后發現,訪花昆蟲主要有鳳蝶、熊蜂和螞蟻(圖1)。其中,鳳蝶為最常見的訪花昆蟲和有效的傳粉動物,它們訪花主要是吸取花蜜,訪花時腹面后翅攜帶大量花粉(肉眼能分辨),訪花頻率為每花每小時0.098 4次。熊蜂訪花時最初停留在花冠上,然后沿著花冠邊緣進入花冠基部取食花蜜,由于忽地笑花絲和花柱較長,熊蜂體型相對小,它們一般不會碰到花藥和柱頭。螞蟻取食花蜜時,在花冠上和花冠基部頻繁活動,其體型很小,身體通常也不會接觸到花藥和柱頭。
鳳蝶在9:00 后開始出現訪花,訪花頻率隨著氣溫升高逐漸增多,14:00—16:00為鳳蝶出現高峰期;隨著氣溫降低和光照減少,鳳蝶出現頻率減少。鳳蝶在單個花序上平均拜訪(1.2±0.1)朵花。我們一共捕捉了14只鳳蝶 ,其中8只為玉斑鳳蝶(Papilio helenus)、3只為藍鳳蝶(P. protenor)、2只為碧鳳蝶(P. bianor)、1只為寬帶鳳蝶(P. nephelus),在解剖鏡下觀察發現花粉均集中在這4種鳳蝶的后翅腹側翅脈上,在前翅腹側觀察到少量花粉,其他部位沒有觀察到(圖4)。而在捕捉到的3只熊蜂和12只螞蟻身體的各個部位中,解剖鏡下觀察并未發現忽地笑的花粉。
2.5 授粉試驗
去雄套袋和不去雄套袋處理均不結實,說明忽地笑不存在孤雌生殖現象和主動自交能力。由表1可知,人工去雄異交坐果率顯著高于人工去雄自交的坐果率(χ2=7.243 1, df=1, P=0.007),但是每果種子數沒有顯著差異(t=-0.686, P=0.494)。基于坐果率和每果種子數得到的自交親和系數分別為0.64 和0.90(表1),表明忽地笑自交后也很有可能結實。補充授粉顯著提高了忽地笑的坐果率(χ2=10.002, df=1, P=0.002),坐果率從36% 增加到 63%(表1),但是每果種子數沒有顯著差異(t=-0.051, P=0.96),說明忽地笑在自然狀態下存在一定程度的花粉限制。
3 討論與結論
忽地笑的花呈蜘蛛花型,具有高度外露的柱頭和花藥,根據前人的研究結果,該種花部特征的植物由蝴蝶翅膀傳粉(Cruden & Hermann-Parker, 1979;Johnson & Bond, 1994;Kiepiel & Johnson, 2014;Epps et al., 2015;Liu et al., 2022),因此我們預測忽地笑的傳粉方式為蝴蝶翅膀傳粉,經過野外定點試驗和觀察,可以確定鳳蝶是忽地笑的有效傳粉動物且是利用其翅膀為忽地笑傳粉。雖然熊蜂和螞蟻是忽地笑的訪花昆蟲,它們沿著花冠邊緣到達花冠筒口并吸取花蜜,但是由于忽地笑的柱頭和花藥離花冠筒口較遠,熊蜂和螞蟻在訪花過程中不能碰觸到它們,在采集到的熊蜂和螞蟻標本身體表面,我們也沒有觀察到忽地笑的花粉,因此熊蜂和螞蟻最有可能是忽地笑的竊蜜者。4種鳳蝶被觀察到拜訪忽地笑的花,在它們的后翅腹側沉積了大量忽地笑的花粉,在拜訪過程中,我們發現蝴蝶的翅膀會接觸到忽地笑外露的柱頭和花藥,因此蝴蝶應該是忽地笑的有效傳粉動物。南非是石蒜科植物的起源中心和多樣性中心(Ito et al., 1999),前人已經發現多種石蒜科植物由蝴蝶翅膀傳粉,它們都是分布在南非,并且發現攜帶花粉的蝴蝶種類也主要是鳳蝶(Johnson & Bond, 1994;Kiepiel & Johnson, 2014;Butler & Johnson, 2020)。本研究證實了在石蒜科中該種傳粉方式在中國也存在,這說明蝴蝶(鳳蝶)翅膀傳粉在石蒜科植物中可能是一個重要的傳粉方式,未來可通過研究石蒜科的系統發育和傳粉方式,來探討石蒜科植物在分化和擴散過程中傳粉模式的演變具有重要科學意義。
忽地笑的花部特征與蝴蝶翅膀傳粉相適應,具體表現在以下三點特征。第一,忽地笑的花和鳳蝶的體型是匹配的,忽地笑的花為兩側對稱,蝴蝶訪花時,抓住花被片和花絲維持身體平衡,口器朝向花冠口以吸取花蜜,此時蝴蝶后翅腹側就可以接觸到高度暴露的花藥和柱頭,這種完美的匹配反映了傳粉動物對植物的花部特征有很強的塑造作用(Fenster et al., 2004)。第二,忽地笑具有雌雄異位的特點,在花剛開放的時候,花柱和花絲是在同一個平面,但是很快花柱向下移動,這樣花柱和花絲就有一個約30°的夾角,蝴蝶來拜訪花的時候,一般停留在花絲上方,后翅腹側可以和花藥接觸,而不能和花絲下方的柱頭接觸。在開花的中后期,花柱向上運動且變得更長,最終花柱和花絲呈平行狀態,但是柱頭往往比花藥高2~3 cm,此時蝴蝶訪花時其后翅往往只接觸到柱頭。雖然柱頭可授性試驗表明柱頭一直都有可授性且忽地笑是自交親和的,但是蝴蝶訪花很難導致花內傳粉,這樣可避免自交和雌雄干擾。第三,忽地笑花藥以縱裂方式裂開,裂開后的形狀呈平口狀,這個特征利于蝴蝶訪花時翅膀觸碰到花藥時大量花粉附著其上。Shwe(2020)就發現在蝴蝶翅膀傳粉的匍莖百合(Lilium lankongense)具有類似的雌雄異位機制以避免花粉折損,同時也具有類似利于花粉附著在蝴蝶翅膀上的花藥裂開方式及裂開形狀。
人工自交授粉試驗證明忽地笑具有一定的自交親和能力,該結果與李雪等(2023)對中國石蒜交配系統的研究結果一致,即以異交為主,有一定的自交能力,這說明石蒜屬植物可能普遍具有一定的自交能力,但這不等同于說明它們有較高的自交率。石蒜屬植物為多年生具鱗莖植物,多年生植物由于反復繁殖,如果有一定的自交率,自交衰退效應會持續累積,會強烈降低個體的繁殖適合度,因此多年生植物往往趨向于異交(任明迅等,2012)。忽地笑雖然有一定自交能力,但是套袋試驗證明其不存在主動自交的能力。忽地笑的雌雄異位基本可以避免同株自花傳粉且鳳蝶每次訪花一般也只訪問1朵花,這樣就可以避免同株異花傳粉。我們推測忽地笑雖然有一定的自交親和性,但是通過特定的花部機制和傳粉動物的互作,使得其自交率維持在一個非常低的水平,這需要通過分子標記來檢驗交配系統加以驗證。
在補充花粉后,忽地笑的坐果率有顯著提高,說明該物種在自然狀態下存在花粉限制,這可能是由于傳粉動物鳳蝶訪問次數少導致,而每果種子數的變化不是很大,可能是由于忽地笑的子房中胚珠數量有限且往往只有1~2個胚珠發育成種子,一次有效拜訪后落置的花粉就足夠忽地笑結實。在石蒜科中許多物種也存在一定的花粉限制,如Haemanthus rotundifolius、Cyrtanthus ventricosus、C. guthrieae(Johnson & Bond, 1997)。有研究發現蝴蝶可適應低糖濃度花蜜(Kingsolver & Daniel, 1979;Goldblatt & Manning, 2002;Martins, 2015),如Johnson和Bond(1994)就發現在蝴蝶傳粉的石蒜科植物中,糖濃度一直較低。但是本研究發現,在忽地笑分泌的花蜜糖濃度在開花后期達到40%以上。這可能反映傳粉動物活動的不足,糖濃度的升高是由于花蜜持續分泌,鳳蝶拜訪較少,而水分通過花冠筒口不斷蒸發,使得糖濃度增加。棲息地喪失和全球氣候變化會使得蝴蝶數量變少(Bonebrake et al., 2016;Jain et al., 2017;Miao et al., 2021),全球氣候變化還會使得蝴蝶的生存空間向高緯度和高海拔移動,隨著氣溫上升,成蟲出現時間也會提前(趙彩云等,2010),這可能導致蝴蝶和其傳粉植物的物候不同步和分布不重疊。在我們研究的居群中,忽地笑的個體以成年植株為主,小個體植株較少(未發表數據),忽地笑的自然更新可能存在障礙,我們需要更多的研究來評估這一重要野生資源的生存繁育狀況。
參考文獻:
ARMBRUSTER WS, EDWARDS ME, DEBEVEC EM, 1994. Floral character displacement generates assemblage structure of Western Australian triggerplants (Stylidium) [J]. Ecology, 75(2): 315-329.
BECERRA JX, LLOYD DG, 1992.Competition-dependent abscission of self-pollinated flowers of Phormium tenax (Agavaceae): a second action of self-incompatibility at the whole flower level? [J]. Evolution, 46(2): 458-469.
BONEBRAKE TC, PICKETT EJ, TSANG TPN, et al., 2016. Warming threat compounds habitat degradation impacts on a tropical butterfly community in Vietnam [J]. Glob Ecol Conserv, 8: 203-211.
BUTLER HC, JOHNSON SD, 2020. Butterfly-wing pollination in Scadoxus and other South African Amaryllidaceae [J]. Bot J Linn Soc, 193(3): 363-374.
CRUDEN RW, HERMANN-PARKER SM, 1979. Butterfly pollination of Caesalpinia pulcherrima, with observations on a psychophilous syndrome [J]. J Ecol, 67(1): 155-168.
DRESSLER RL, 1968. Pollination by euglossine bees [J]. Evolution, 22(1): 202-210.
EPPS MJ, ALLISON SE, WOLFE LM, 2015. Reproduction in flame azalea ( Rhododendron calendulaceum, Ericaceae): a rare case of insect wing pollination [J]. Am Nat, 186(2): 294-301.
FENSTER CB, ARMBRUSTER WS, WILSON P, et al., 2004. Pollination syndromes and floral specialization [J]. Ann Rev Ecol Evol Syst, 35(1): 375-403.
FERRERO V, DEVEGA C, STAFFORD GI, et al., 2009. Heterostyly and pollinators in Plumbago auriculata (Plumbaginaceae) [J]. S Afr J Bot, 75(4): 778-784.
GOLDBLATT P, MANNING JC, 2002. Evidence for moth and butterfly pollination in Gladiolus (Iridaceae-Crocoideae) [J]. Ann Mo Bot Gard, 89(1): 110-124.
HARDER LD, 1990. Pollen removal by bumble bees and its implications for pollen dispersal [J]. Ecology, 71(3): 1110-1125.
HUANG SQ, SHI XQ, 2013. Floral isolation in Pedicularis: how do congeners with shared pollinators minimize reproductive interference? [J]. New Phytol, 199(3): 858-865.
JAIN A, LIM FKS, WEBB EL, 2017. Species-habitat relationships and ecological correlates of butterfly abundance in a transformed tropical landscape [J]. Biotropica, 49(3): 355-364.
JENNERSTEN O, 1984. Flower visitation and pollination efficiency of some North European butterflies [J]. Oecologia, 63(1): 80-89.
JI YB, XIN GS, QU ZY, et al., 2016. Research progress on chemical constituents and pharmacological effects of alkaloids from plants of Lycoris Herb [J]. Chin Tradit Herbal Drugs, 47(1): 157-164. [季宇彬, 辛國松, 曲中原, 等, 2016. 石蒜屬植物生物堿類化學成分和藥理作用研究進展 [J]. 中草藥, 47(1): 157-164.]
JOHNSON SD, 2010. The pollination niche and its role in the diversification and maintenance of the southern African flora [J]. Philos T R Soc B, 365(1539): 499-516.
JOHNSON SD, BOND WJ, 1994. Red flowers and butterfly pollination in the fynbos of South Africa [M]. Bloomberg: Kluwer Academic Publishers: 137-148.
JOHNSON SD, BOND WJ, 1997. Evidence for widespread pollen limitation of fruiting success in Cape wildflowers [J]. Oecologia, 109(4): 530-534.
KIEPIEL I, JOHNSON SD, 2014. Shift from bird to butterfly pollination in Clivia (Amaryllidaceae) [J]. Am J Bot, 101(1): 190-200.
KINGSOLVER JG, DANIEL TL, 1979. On the mechanics and energetics of nectar feeding in butterflies [J]. J Theor Biol, 76(2): 167-179.
LI X, PENG Y, LI FX, 2023. Temporal dynamics of mating systems in Lycoris chinensis [J/OL]. Mol Plant Breed: 1-7 [2024-01-14]. http://kns.cnki.net/kcms/detail/46.1068.S.20230202. 1447. 007.html. [李雪, 彭冶, 李豐霞, 2023. 中國石蒜交配系統的實時動態變化 [J/OL]. 分子植物育種: 1-7 [2024-01-14]. http://kns.cnki.net/kcms/detail/46.1068.S.20230202.1447.007. html.]
LIAO Y, 2009. The study on biological characteristics of Papilio polytes(Linnaeu) [D]. Fuzhou: Fujian Agriculture and Forestry University: 26-27. [廖宇, 2009. 玉帶鳳蝶生物學特性研究 [D]. 福州: 福建農林大學: 26-27.]
[JP3]LIU CQ, NIU Y, LU QB, et al., 2022. Papilio butterfly vs. hawkmoth pollination explains floral syndrome dichotomy in a clade of Lilium [J]. Bot J Linn Soc, 199(3): 678-693.[JP2]
ITO M, KITA Y, YUKAWA T, et al., 1999. Phylogenetic relationships of Amaryllidaceae based on matK sequence data [J]. J Plant Res, 112: 207-216.
MARTINS DJ, 2015. Butterfly pollination of the dryland wildflower Gloriosa minor [J]. J East Afr Nat Hist, 103(1): 25-30.
MASI M, DI LR, CIMMINO A, et al., 2020. Advances in the chemical and biological characterization of Amaryllidaceae alkaloids and natural analogues Isolated in the last decade [J]. Molecules, 25(23): 5621.
MASSINGA PH, JOHNSON SD, HARDER LD, 2005. Heteromorphic incompatibility and efficiency of pollination in two distylous Pentanisia species (Rubiaceae) [J]. Ann Bot, 95(3): 389-399.
MIAO BG, PENG YQ, YANG DR, et al., 2021. Climate and land-use interactively shape butterfly diversity in tropical rainforest and savanna ecosystems of southwestern China [J]. Insect Sci, 28(4): 1109-1120.
MUCHHALA N, POTTS MD, 2007. Character displacement among bat-pollinated flowers of the genus Burmeistera: analysis of mechanism, process and pattern [J]. Proc R Soc B, 274(1626): 2731-2737.
OLLERTON J, WINFERR R, TARRANT S, 2011. How many flowering plants are pollinated by animals? [J]. Oikos, 120(3): 321-326.
PAN CP, 2023. Chinese “Tulipa” — Lycoris [J]. China Flowers Hortic (6): 32-36. [潘春屏, 2023. 中國“郁金香”——石蒜 [J]. 中國花卉園藝 (6): 32-36.]
PATON AJ, BRUMMITT N, GOVAERTS R, et al., 2008. Toward starget 1 of the global strategy for plant conservation: a working list of all known plant species — progress and prospects [J]. Taxon, 57(2): 602-611.
PAUW A, 2006. Floral syndromes accurately predict pollination by a specialized oil-collecting bee (Rediviva peringueyi, Melittidae) in a guild of South African orchids (Coryciinae) [J]. Amer J Bot, 93(6): 917-926.
PEI J, 1985. Lycoris aurea [M]// PEI J, DING ZZ. Flora Republicae Popularis Sinicae: Vol. 16(1). Beijing: Science Press: 20 [裴鑒, 1985. 忽地笑 [M]// 裴鑒, 丁志遵. 中國植物志: 第16(1)卷. 北京: 科學出版社: 20.]
REN MX, JIANG XH, ZHANG DY, 2012. SIwdLdILl+/1mphPTcwxSpw==ome important questions in plant reproductive ecology [J]. Biodivers Sci, 20(3): 241-249. [任明迅, 姜新華, 張大勇, 2012. 植物繁殖生態學的若干重要問題 [J]. 生物多樣性, 20(3): 241-249.]
SHWE EE, 2020. Butterfly pollination in a lily flower with plastic sexual systems [D]. Wuhan: Central China Normal University: 37-54.
SUN ZF, MU SZ, ZHANG JX, et al., 2013. Chemical constituents of Lycoris aurea from Guizhou Province [J]. J N Pharm, 10(9): 1-2. [孫志峰, 穆淑珍, 張建新, 等, 2013. 黔產藥用植物忽地笑中的化學成分研究 [J]. 北方藥學, 10(9): 1-2.]
YUAN JH, HU MH, 2009. A review on germplasm resources of Lycoris and its exploitation and utilization [J]. Subtrop Plant Sci, 38(2): 79-84. [袁菊紅, 胡綿好, 2009. 石蒜屬種質資源及其開發利用研究 (綜述) [J]. 亞熱帶植物科學, 38(2): 79-84.]
ZHAO CY, LI JS, LUO JW, et al., 2010. A review on responses of butterflies to global climate change [J]. Acta Ecol Sin, 30(4): 1050-1057. [趙彩云, 李俊生, 羅建武, 等, 2010. 蝴蝶對全球氣候變化響應的研究綜述 [J]. 生態學報, 30(4): 1050-1057.]
ZHOU Y, 1998. Classification and identification of Chinese butterflies [M]. Zhengzhou: Henan Science and Technology Press. [周堯, 1998. 中國蝴蝶分類與鑒定 [M]. 鄭州: 河南科學技術出版社.]
