黃瓜FtsZs基因家族鑒定及分析
2024-10-29 00:00:00陳琳杰常婧姝張宇顏爽任仲海王麗娜
山東農業大學學報(自然科學版) 2024年5期



關鍵詞:黃瓜;FtsZs基因;生物信息學分析;非生物脅迫反應;生物脅迫反應
黃瓜(CucumissativusL.)是世界性的十大蔬菜作物之一,而我國是黃瓜的主產國。2022年,我國黃瓜種植面積為131.15萬hm2,總產量為7730.73萬噸,分別占全球的60.32%和81.62%,均居世界第一位。
FtsZ(FilamentingtemperaturesensitiveZ)蛋白作為一種可溶性的胞質蛋白,是微管蛋白的結構同系物。FtsZ在離體條件下經誘導可通過多聚化作用組裝成線形排列的有序結構,而這些由FtsZ蛋白組成的結構則與tubulin形成的原纖絲非常類似[1],暗示了其功能的相似性。FtsZ蛋白多定位于質體或葉綠體,并作為質體分裂復合體的基本組成部分,組裝成FtsZ環并招募至少二十多種蛋白質介導胞質分裂[2]。FtsZ可直接或間接地調控葉綠體分裂環中蛋白質的組裝和定位,并驅動葉綠體的分裂。有研究表明,擬南芥FtsZ1、FtsZ2-1和FtsZ2-2定位于葉綠體的基質和類囊體上,可以通過影響葉綠體分裂,使葉綠體變大[3-5]。水稻(Oryzasativa)OsFtsZ1和OsFtsZ2-1過表達和敲低都會抑制淀粉體分裂,產生多形性淀粉顆粒[6]。與野生型相比,馬鈴薯ftsz1敲除突變體塊莖中淀粉顆粒增大或淀粉粒黏度增加[7]。在小立碗蘚中,利用基因編輯技術獲得了ftsz2-1突變體,研究發現,ftsz2-1突變體的葉綠體分裂受到顯著影響,甚至在其少數細胞中能觀察到單個不分裂的絲狀葉綠體[8]。在煙草中敲低FtsZ基因會干擾葉綠體的正常分裂過程,引起葉綠體發育的異常,并導致葉綠體數目的減少和體積的增加[9]。
目前黃瓜FtsZs家族的研究還未見報道,其功能尚不清楚。為了解析黃瓜FtsZs基因的功能,本文對黃瓜FtsZs家族成員進行了鑒定,分析了其基因結構及在染色體上的分布,研究了黃瓜FtsZs與擬南芥、番茄、甜瓜FtsZs的親緣關系,確定了其組織表達特性及對生物和非生物脅迫的響應情況,研究結果可為探明黃瓜FtsZs作用機制提供數據支撐。
1 材料與方法
1.1 FtsZs家族基因的鑒定
分別從葫蘆科基因組網站(http://CucurbitGenomicsDatabase(CuGenDB)、擬南芥基因組網站(https://www.arabidopsis.org)和番茄基因組網站(https://solgenomics.net)下載黃瓜、甜瓜、擬南芥和番茄gff、cds、pep、fasta文件,獲得黃瓜、甜瓜、擬南芥和番茄所有注釋基因信息。隨后從Pfam數據庫(http://www.example.com)下載FtsZ保守域(PF12327)的HMM配置文件,在所下載注釋基因信息中篩選黃瓜、甜瓜、擬南芥和番茄的FtsZs基因(默認參數和截止值為0.01)。使用Pfam工具(http://www.example.com)和SMART(http://www.example.com)對篩選獲得的黃瓜、甜瓜、擬南芥和番茄FtsZ保守域進行鑒定,并根據其與擬南芥FtsZs的親緣關系進行命名;使用ExPasy(htps:/web.ExPasy.org/computepI/)計算FtsZs的相對分子量(Mw)、氨基酸數量(AA)和等電點(pI);使用Plant-mPLoc(http://www.csbio.situ.edu.cn/bioinf/plant-multi/)預測FtsZs的亞細胞定位情況。
1.2 黃瓜、甜瓜、擬南芥和番茄FtsZs進化關系分析
利用MEGA7軟件中的ClustalX對所有FtsZs蛋白序列進行對比,隨后采用鄰接法(Neighbor-Joiningmethod,NJ)構建系統進化樹,并用Bootstrap法(重復次數設定為1000)評估系統進化樹,最終使用MEGA7呈現進化樹。
1.3 黃瓜FtsZs基因及蛋白結構的分析
從黃瓜基因組網站(http://CucurbitGenomicsDatabase(CuGenDB))獲得FtsZs的內含子和外顯子信息,并利用TBtools生成基因結構;使用MEME(htps://www.omicsclass.com/article/432)在線網站分析黃瓜FtsZs的保守motif(設置基序數為10,基序的最小長度為6個氨基酸,基序的最大長度為10個氨基酸),并用TBtools可視化[10];從黃瓜基因組網站(http://CucurbitGenomicsDatabase(CuGenDB))獲得FtsZs的蛋白序列,從SMART網站獲得Tubulindomain和FtsZ-Cdomain兩個保守結構域的蛋白序列,利用DNAMAN生成FtsZs蛋白序列并分析蛋白序列的差異。
1.4 黃瓜FtsZs組織表達特性分析
基于已有的RNA-seq數據(SRA046916)[11],分析了黃瓜FtsZs在根、莖、葉、雌花、雄花、子房、未受精子房、已受精子房和卷須等九個組織的表達情況。并基于FPKM值使用TBtools軟件(v2.086)可視化。
1.5 實時熒光定量PCR分析黃瓜FtsZs表達特性
將華北型黃瓜“9930”種于山東農業大學園藝實驗站9號溫室,并按照常規栽培方法進行管理。在結果期取根、莖、葉、雌花、雄花、卷須和果實等組織,采用TRIzol(湖南艾科瑞生物工程有限公司)提取RNA,并用EvoM-MLV反轉錄預混型試劑盒Ver.2(t湖南艾科瑞生物工程有限公司)逆轉錄成cDNA。使用PrimerPremier5.0設計引物,并以黃瓜肌動蛋白基因作為內參(引物序列見附表S1)。采用SYBRGreenProTaqHS預混型qPCR試劑盒(湖南艾科瑞生物工程有限公司)在CFXOpus96實時PCR檢測系統(Bio-Rad,Hercules,CA,USA)進行實時熒光定量PCR檢測。采用2-ΔΔCt方法計算所選基因的相對表達水平[12]。對每個基因進行3次生物學和3次技術重復。
1.6 黃瓜FtsZs響應非生物和生物脅迫分析
從NCBI(https://www.ncbi.nlm.nih.gov/)下載了鹽脅迫(GSE116265)[13]、高溫脅迫(GSE151055)[14]、冷處理(GSE210703)[15]和白粉病菌侵染(GSE81234)[16]的轉錄組數據,以分析黃瓜FtsZs的響應情況,并用TBtools可視化。
2 結果與分析
2.1 黃瓜、甜瓜、擬南芥和番茄FtsZs的鑒定
本文在黃瓜、甜瓜、擬南芥和番茄中分別鑒定到了3個、3個、3個和4個FtsZs基因,并按親緣關系對其進行命名(見表1)。通過比對基因結構發現,基因間的長度相差很大,介于2050和17560bp之間,其中最短的基因為AtFtsZ1(2050bp),最長的基因為SlFtsZ2-1(17560bp);它們的外顯子數目介于6和8之間,黃瓜FtsZs基因的外顯子多為7個,擬南芥FtsZs基因的外顯子多為6個,番茄FtsZs基因的外顯子多為7個。蛋白結構分析發現,氨基酸長度介于419和491AA之間;大部分基因偏酸性(AtFtsZ1,AtFtsZ2-1,AtFtsZ2-2,CsFtsZ1,CsFtsZ2-1,CsFtsZ2-2,SlFtsZ1-1,SlFtsZ2-1,SlFtsZ2-2,CmFtsZ1,CmFtsZ2-1,CmFtsZ2-2),只有1個基因偏堿性(SlFtsZ1-2)。黃瓜FtsZs基因長度與其他物種的差異較大,暗示FtsZs在不同物種中可能發揮不同的功能。
亞細胞定位預測結果顯示,所有的FtsZ1s基因都在葉綠體和細胞質中有定位信號(AtFtsZ1,CsFtsZ1,SlFtsZ1-1,SlFtsZ1-2,CmFtsZ1),黃瓜和甜瓜的FtsZ2s基因都有細胞質定位信號(CsFtsZ2-1,CsFtsZ2-2,CmFtsZ2-1,CmFtsZ2-2),而擬南芥和番茄的FtsZ2s基因都有葉綠體定位信號(AtFtsZ2-2,SlFtsZ2-1,SlFtsZ2-2)。
2.2 黃瓜、甜瓜、擬南芥和番茄FtsZs進化關系分析
根據親緣關系的遠近,我們可以將FtsZs基因分為2個組。第一組包括1個黃瓜FtsZ基因(CsFtsZ1),1個甜瓜FtsZ基因(CmFtsZ1),1個擬南芥FtsZ基因(AtFtsZ1)和2個番茄FtsZs基因(SlFtsZ1-1,SlFtsZ1-2),其中黃瓜FtsZ基因和甜瓜FtsZ基因的親緣關系最近,其次是擬南芥和番茄。第二個組包括2個黃瓜FtsZs基因(CsFtsZ2-1,CsFtsZ2-2),2個番茄FtsZs基因(SlFtsZ2-1,SlFtsZ2-2),2個擬南芥FtsZs基因(AtFtsZ2-1,AtFtsZ2-2),2個甜FtsZs基因瓜(CmFtsZ2-1,CmFtsZ2-2),其中黃瓜FtsZs基因和甜瓜FtsZs基因的親緣關系最近,其次是擬南芥FtsZs基因和番茄FtsZs基因(圖1)。
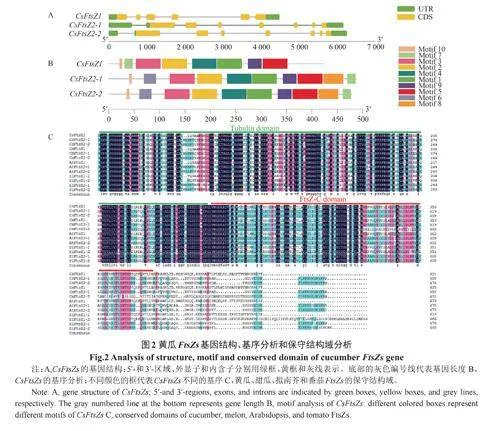
2.3 黃瓜FtsZs基因結構分析
研究發現,CsFtsZ1有6個外顯子,氨基酸長度為421AA;CsFtsZ2-1有7個外顯子,氨基酸長度為488AA;CsFtsZ2-2有7個外顯子,氨基酸長度為479AA。CsFtsZ1與CsFtsZ2-1、CsFtsZ2-2在外顯子數目和氨基酸長度上存在差異,CsFtsZ2-1與CsFtsZ2-2在基因結構上差異不大。
保守motif基序分析發現,黃瓜3個FtsZs基因均含有M1、M2、M3、M4、M5、M7、M9、M10這8個基序,而M6和M8這2個基序只存在CsFtsZ2-1和CsFtsZ2-2中。
保守結構域分析發現,黃瓜、甜瓜、擬南芥和番茄的FtsZs都含有Tubulindomain和FtsZ-Cdomain結構域,暗示了基因進化上的保守性。
2.4 黃瓜FtsZs基因的組織表達模式
RNA-seq數據顯示(圖3),3個黃瓜FtsZs基因在葉、莖、子房和雌花中表達量較高,推測其參與了葉、莖、子房和雌花的生長發育。其中,CsFtsZ2-1和CsFtsZ2-2在雌花和未受精子房中的表達量差異較大,暗示其在雌花和子房生長發育中發揮不同的功能。
熒光定量PCR結果顯示,黃瓜CsFtsZ2-1基因在華北型黃瓜“9930”葉片、雄花和雌花中的表達量較高。
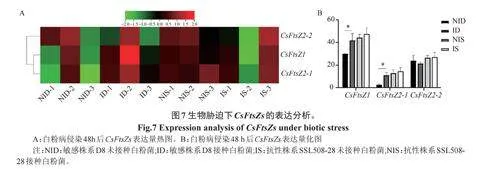
2.5 黃瓜FtsZs基因在生物脅迫和非生物脅迫下的表達譜
利用鹽脅迫、高溫處理、冷脅迫和白粉病菌侵染RNA-seq數據,分析了黃瓜FtsZs基因在脅迫下的響應情況。首先,我們分析了黃瓜FtsZs基因在鹽脅迫下的表達情況,結果顯示,NaCl處理后,黃瓜CsFtsZ2-1基因下調,表明該基因可響應鹽脅迫;硅(Si)被認為是植物的第四大基本元素,可增強抗逆性并促進植物的生長發育。在僅用Si處理的條件下和外源鹽加硅(NaSi)處理下,黃瓜FtsZs的表達量差異不大(圖4)。
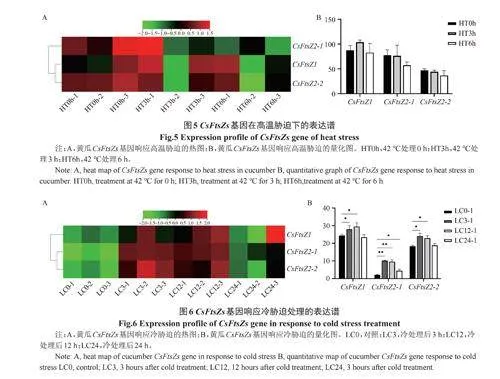
黃瓜FtsZs基因對高溫處理響應結果表明(圖5),高溫處理3h和高溫處理6h后,黃瓜中的3個FtsZs基因表達水平沒有明顯變化,這表明黃瓜中的3個FtsZs基因不響應高溫脅迫。
黃瓜FtsZs基因對冷脅迫響應結果表明(圖6),冷處理3h和12h后,黃瓜中3個FtsZs基因表達均上調,而冷處理24h后只有CsFtsZ2-1基因上調,這表明黃瓜中的3個FtsZs基因均可響應冷脅迫。
分析了黃瓜FtsZs基因對白粉病的響應情況,結果顯示,在白粉病敏感株系D8中,接種白粉病后,CsFtsZ1和CsFtsZ2-1的表達量顯著提高,表明兩者可響應白粉病病原菌侵染;在白粉病抗性株系SSL508-28中,接種白粉菌前后,黃瓜FtsZs的表達量差異不大(圖7)。
3 討論
迄今為止,在大多數的原核生物中均發現了FtsZ基因,并對其功能進行了深入的研究。相較于原核生物,真核生物中FtsZs基因的報道較少。本文在黃瓜、甜瓜、擬南芥和番茄中分別鑒定到了3~4個FtsZs基因,暗示了FtsZs基因數目在不同物種間具有高度的保守性。擬南芥3個FtsZs基因中最長的為3269bp,最短的為2050bp;黃瓜3個FtsZs基因中最長的為6264bp,最短的為4493bp;甜瓜3個FtsZs基因中最長的為13038bp,最短的為4410bp;番茄4個FtsZs基因中最長的為17560bp,最短的為6365bp,暗示著在不同物種之間FtsZs基因的結構存在差異性(表1)。本文鑒定到的13個FtsZs基因都含有Tubulindomain和FtsZ-Cdomain兩個保守的結構域,但是不同物種之間保守結構域的基序是有差異的,即使是同屬于葫蘆科的黃瓜和甜瓜,它們保守結構域的基序也不完全相同(圖2),這就暗示了該基因在進化上既有保守性又存在差異性,進一步說明不同物種之間FtsZs基因功能可能不盡相同。
有研究發現,木薯中的MeFtsZ2-1和MeFtsZ2-2定位于葉綠體內[17]。將MeFtsZ2-1和MeFtsZ2-2在擬南芥中異源表達,可影響擬南芥葉肉細胞中的葉綠體分裂進而使葉綠體數量減少[17];在擬南芥中抑制AtFtsZ1-1或AtFtsZ2-1的表達也會通過影響葉綠體分裂使成熟葉細胞中葉綠體的數量減少[18]。我們利用在線網站預測了黃瓜中3個FtsZs基因的亞細胞定位情況,發現它們在葉綠體或細胞質中有定位信號,推測它們在黃瓜的葉綠體中起作用(表1)。
已有的研究表明,在木薯中過表達MeFtsZ2-1基因會使木薯植株早開花,頂端分支明顯增多,葉片變小且數量增多[17]。我們利用RNS-seq數據和熒光定量PCR技術分析發現,黃瓜中3個FtsZs基因在葉中都有較高表達,推測其在黃瓜葉中發揮重要功能(圖3)。我們利用熒光定量PCR技術研究發現,CsFtsZ2-1在雄花和雌花中表達量也較高,推測CsFtsZ2-1也會影響花的發育(圖3),但其具體作用機制還需要做深入的研究。
前人研究表明,在300mMNaCl脅迫下,木薯FtsZ2-1的轉錄水平先降低后升高,在NaCl脅迫4h時達到最低水平,在NaCl脅迫48h時達到最高水平[19]。本文結果顯示,黃瓜FtsZ2-1基因在75mM鹽處理下表達下調。除此之外,我們還發現黃瓜CsFtsZ1、CsFtsZ2-1和CsFtsZ2-1基因可響應冷脅迫,CsFtsZ1和CsFtsZ2-1可響應白粉病菌侵染,暗示著黃瓜FtsZs基因在鹽脅迫、冷脅迫和白粉病抗性中發揮作用。
4 結論
本文在黃瓜中鑒定到3個FtsZs基因,基因結構分析發現黃瓜FtsZs基因長度為1266~1467bp,外顯子數目為6~7個,氨基酸長度為421~488AA;保守結構域分析發現它們都含有Tubulindomain和FtsZ-Cdomain兩個保守的結構域。組織表達特異性分析發現,FtsZs在黃瓜葉、莖、子房和雌花組織中表達量較高;CsFtsZ2-1基因可響應鹽脅迫,CsFtsZ1、CsFtsZ2-1和CsFtsZ2-2可響應冷脅迫,CsFtsZ1和CsFtsZ2-1可響應白粉病病原菌侵染。
