草地貪夜蛾與東方黏蟲的種內(nèi)及種間斗爭行為差異研究
2024-11-03 00:00:00馬文玲楊燈海范恒喆盛成旺胡飛楊斌操海群王桂榮蔣興川
植物保護(hù) 2024年5期
摘要昆蟲之間存在復(fù)雜的種內(nèi)及種間競爭關(guān)系,競爭中的獲勝方可以占有食物、配偶等資源,其中,斗爭行為是昆蟲競爭的主要方式之一。草地貪夜蛾Spodoptera frugiperda是我國玉米上的重大入侵性害蟲,其入侵過程伴隨著與本地害蟲的競爭,草地貪夜蛾與本地害蟲的種內(nèi)及種間競爭能力差異是其能否成功入侵、定殖甚至取代本地種群的關(guān)鍵。東方黏蟲Mythimna separata是我國玉米上重要的本地害蟲,本文分別對不同齡期草地貪夜蛾和東方黏蟲幼蟲的種內(nèi)及種間斗爭行為進(jìn)行深入研究。在種內(nèi)斗爭行為中,不同齡期東方黏蟲幼蟲均表現(xiàn)出較高頻次的攻擊行為,以頭部接觸行為發(fā)生最為頻繁;而草地貪夜蛾僅高齡幼蟲對低齡幼蟲表現(xiàn)出顯著攻擊性,如在6齡 vs 4齡中,頭部接觸頻次為(0.318±0.073)次/min、猛擊頻次為(0.313±0.059)次/min。在種間斗爭行為中,不同齡期草地貪夜蛾幼蟲均對東方黏蟲表現(xiàn)出顯著高頻次且具有殺傷力的攻擊行為,如在4齡 vs 4齡、5齡 vs 5齡、6齡 vs 6齡、4齡 vs 6齡和6齡 vs 4齡中,猛擊頻次分別為(0.337±0.052)次/min、(0.695±0.127)次/min、(0.546±0.051)次/min、(0.362±0.043)次/min和(1.026±0.172)次/min;而東方黏蟲僅4齡幼蟲對同齡期草地貪夜蛾表現(xiàn)出顯著攻擊性,其他齡期幼蟲的攻擊和防御行為沒有顯著差異。該研究結(jié)果為闡明鱗翅目害蟲幼蟲的斗爭行為提供了重要的參考依據(jù),為闡明害蟲的種內(nèi)及種間互作關(guān)系提供了理論基礎(chǔ)。
關(guān)鍵詞草地貪夜蛾;東方黏蟲;斗爭行為;害蟲互作;種內(nèi)競爭;種間競爭
中圖分類號:S 435.132文獻(xiàn)標(biāo)識碼:ADOI:10.16688/j.zwbh.2023526The difference of intra- and inter-specific agonistic behaviors between
Spodoptera frugiperda and Mythimna separataMA Wenling YANG Denghai FAN Hengzhe SHENG Chengwang HU Fei YANG Bin CAO Haiqun WANG Guirong JIANG Xingchuan(1. Anhui Provincial Key Laboratory for Integrated Management of Crop Pests, Key Laboratory of Biology
and Sustainable Management of Plant Diseases and Pests of Anhui Higher Education Institutes, College
of Plant Protection, Anhui Agricultural University, Hefei230036, China; 2. State Key Laboratory for
Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of
Agricultural Sciences, Beijing100193, China; 3. Institute of Plant Protection and Agro-Products
Safety, Anhui Academy of Agricultural Sciences, Hefei230031, China)AbstractInsects engage in complicated forms of intra- and inter-specific competition to secure essential resources, including food and mates. Agonistic behaviors, notably fighting, are a crucial aspect of insect competition. The fall armyworm (Spodoptera frugiperda) is an important invasive pest in the maize fields of China, competing with local pests during its invasion. The difference in intra- and inter-specific competitive abilities between the fall armyworm and local pests is key to the successful invasion, colonization, and even replacement of the local population. The oriental armyworm (Mythimna separata) is one of the local pests in the maize fields. This study delves into the agonistic behaviors of the two pests with different instars in intra- and inter-specific competition. In the intraspecific agonistic encounters, the oriental armyworm exhibited a heightened frequency of attack behaviors, especially the head-touching behavior. The older larvae of the fall armyworm showed the conspicuously aggressive tendencies towards their younger counterparts, the head-touching and strike behaviors in the larval combination of 6th instar vs 4th instar were (0.318±0.073) and (0.313±0.059) counts per min, respectively. In the interspecific agonistic encounters, the fall armyworm consistently demonstrated significantly elevated and lethal attack behaviors towards the oriental armyworm, the strike behavior in the larval combinations of 4th instar vs 4th instar, 5th instar vs 5th instar, 6th instar vs 6th instar, 4th instar vs 6th instar, and 6th instar vs 4th instar were (0.337±0.052), (0.695±0.127), (0.546±0.051), (0.362±0.043), and (1.026±0.172) counts per min, respectively. The oriental armyworm did not exhibit noteworthy differentials in attack and defense behaviors towards the fall armyworm, except for the predominantly aggressive behaviors in the larval combinations of 4th instar vs 4th instar. These empirical findings contribute to illustrate the agonistic behaviors of lepidopteran larvae and provide a theoretical underpinning for deciphering the intra- and inter-specific dynamics governing pest interactions.
Key wordsSpodoptera frugiperda;Mythimna separata;agonistic behavior;pest interactions;intraspecific competition;interspecific competition
相同生態(tài)位的物種往往會為了有限的食物資源或生存空間而競爭,同種個體之間的競爭為種內(nèi)競爭,異種個體之間的競爭為種間競爭[1]。昆蟲的種內(nèi)及種間競爭現(xiàn)象發(fā)生較為普遍,種內(nèi)競爭和種間競爭的相對強度是影響種群發(fā)展及演替的重要因素[23]。競爭有2種作用方式,一種是利用性競爭,競爭個體共同損耗有限的資源,而不直接相互作用;第二種是干擾性競爭,競爭個體通過直接的相互作用展開競爭[4]。不同物種的競爭優(yōu)勢與環(huán)境溫度、種群密度及寄主種類等密切相關(guān)[5]。在昆蟲生長過程中,適當(dāng)?shù)姆N內(nèi)競爭有利于種群發(fā)展,然而,一旦種內(nèi)競爭失去平衡,會對種群穩(wěn)定性產(chǎn)生負(fù)面影響。例如,在橘小實蠅Bactrocera dorsalis或番石榴實蠅Bactrocera correcta種群中,過度的種內(nèi)競爭會導(dǎo)致幼蟲發(fā)育歷期縮短、蛹發(fā)育歷期延長及成蟲死亡率增加等現(xiàn)象發(fā)生[2]。在種間競爭中,昆蟲可以通過生理調(diào)節(jié)或行為適應(yīng)而獲得競爭優(yōu)勢。當(dāng)橘小實蠅和番石榴實蠅幼蟲處于同一生境時,橘小實蠅通過縮短發(fā)育歷期競爭有限的生存資源,從而在與番石榴實蠅的種間競爭中獲得更高的存活率[6]。黑豆蚜Aphis fabae和豌豆蚜Acyrthosiphon pisum的種間競爭顯著抑制豌豆蚜的體重和產(chǎn)蚜量增加,但是可在一定程度上促進(jìn)黑豆蚜的生長發(fā)育和生殖[7]。天敵昆蟲的種內(nèi)或種間競爭也會影響其對害蟲的控制作用。隨著捕食性天敵微小花蝽Orius minutus、深點食螨瓢蟲Stethorus punctillum和塔六點薊馬Scolothrips takahashii的密度增加,種內(nèi)及種間的干擾和競爭逐漸增加,對朱砂葉螨Tetranychus cinnabarinus的捕食率逐漸下降[8]。此外,競爭還發(fā)生在昆蟲的不同發(fā)育階段。在寄生性天敵的種間競爭中,棱角腫腿蜂Goniozus nephantidis成蟲和幼蟲在與長角繭蜂Bracon brevicornis競爭同一寄主時均具有明顯優(yōu)勢[9]。
在昆蟲的種內(nèi)及種間競爭中,通過物理攻擊進(jìn)行的斗爭行為非常普遍。例如,雄性黑腹果蠅Drosophila melanogaster為了爭奪配偶會有拍打翅膀、猛撲等攻擊行為,獲勝方擁有優(yōu)先與雌性交配的權(quán)利[1011],當(dāng)雌性不愿意接受雄性的求偶時,會通過蹬腿、揮動翅膀等行為攻擊雄性[12]。果蠅還會為了食物發(fā)生激烈的斗爭行為,雄性的斗爭行為表現(xiàn)比雌性更具等級關(guān)系[13]。澳大利亞黃蜂Pseudabispa paragioides和大黃蜂Abispa ephippium生態(tài)位重疊,雌性澳大利亞黃蜂會攻擊并殺死雌性大黃蜂,從而侵占它們的巢穴并捕食它們的幼蟲[14]。天敵煙蚜繭蜂Aphidius gifuensis和短翅蚜小蜂Aphelinus asychis共寄生于同一寄主時,物理攻擊在煙蚜繭蜂排除競爭者中發(fā)揮了重要的作用[1516]。此外,昆蟲的聲波振動也可以對競爭者發(fā)出驅(qū)避或攻擊信號。雙線鉤蛾Drepana arcuata幼蟲通過下顎骨振動或尾部末端槳狀剛毛摩擦植物葉片發(fā)出聲音驅(qū)趕入侵者,保護(hù)領(lǐng)地[17]。蠶蛾總科幼蟲可以發(fā)出4種截然不同的防御性聲音,用來警告或恫嚇捕食者[18]。
為了明確昆蟲斗爭行為的特點,研究者按動作類型對斗爭行為進(jìn)行了分類,包括攻擊行為和防御行為[1921]。如草地貪夜蛾S.frugiperda和美洲棉鈴蟲Helicoverpa zea斗爭時存在頭部接觸(head touching)和猛擊(strike)2種攻擊行為,及畏縮(recoil)、扭動(wriggle)、離開(move away)和翻滾(roll)4種防御行為[20];草地貪夜蛾和亞洲玉米螟Ostrinia furnacalis斗爭時存在頭部接觸和猛擊2種攻擊行為及畏縮和離開2種防御行為[21];黑腹果蠅之間斗爭時存在靠近(approach)等9種攻擊行為和走開(walk away)等4種防御行為[19]。不同昆蟲的斗爭行為特征有所不同,昆蟲斗爭行為的鑒定、分類及發(fā)生強度是評價昆蟲競爭關(guān)系的重要依據(jù)。
草地貪夜蛾屬于鱗翅目Lepidoptera夜蛾科Noctuidae,于2018年12月入侵我國,是重大遷飛性農(nóng)業(yè)害蟲[22],主要為害玉米,喜食玉米莖葉和果穗等[2324]。田間調(diào)查發(fā)現(xiàn),草地貪夜蛾與入侵地玉米楷夜蛾Busseola fusca、非洲大螟Sesamia calamistis、玉米禾螟Chilo partellus、斜紋夜蛾Spodoptera litura、稻蛀莖夜蛾Sesamia inferens和粉紋夜蛾Trichoplusia ni等害蟲之間存在競爭關(guān)系[2527],其與本地種群之間的競爭能力差異是其能否成功入侵、定殖甚至取代本地種群的關(guān)鍵。在草地貪夜蛾與本地害蟲的競爭研究中,幼蟲的斗爭行為是一個關(guān)注度較高的研究熱點[2021,2829]。東方黏蟲Mythimna separata是嚴(yán)重為害玉米的本地農(nóng)業(yè)害蟲,然而,入侵害蟲草地貪夜蛾與本地害蟲東方黏蟲幼蟲的競爭關(guān)系尚不明確,其斗爭行為的特點及差異有待進(jìn)一步探究。本研究分析了不同齡期的草地貪夜蛾和東方黏蟲幼蟲在種內(nèi)及種間競爭中斗爭行為的發(fā)生頻率,為解析鱗翅目害蟲幼蟲的斗爭行為特點提供了重要的參考依據(jù),為闡明害蟲的種內(nèi)及種間互作關(guān)系提供了理論基礎(chǔ)。
1材料與方法
1.1供試蟲源
草地貪夜蛾和東方黏蟲種群采自安徽農(nóng)業(yè)大學(xué)玉米試驗田內(nèi),以人工飼料飼養(yǎng)于植物保護(hù)學(xué)院昆蟲行為與化學(xué)生態(tài)實驗室。飼養(yǎng)條件:(27±1)℃,相對濕度(70±5)%,光周期L∥D=16 h∥8 h[30]。
1.2試驗方法
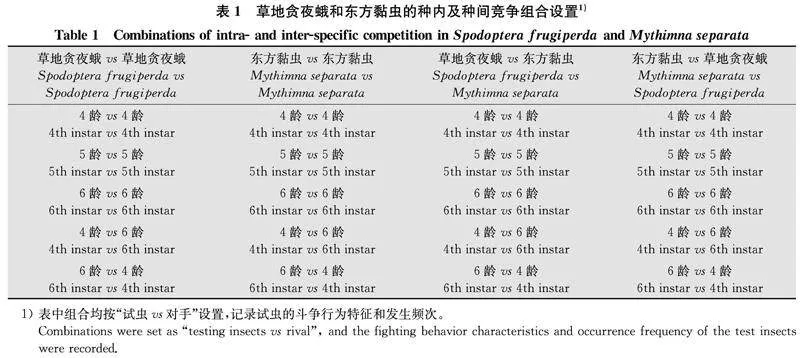
草地貪夜蛾與東方黏蟲的種內(nèi)及種間斗爭行為試驗參考草地貪夜蛾與美洲棉鈴蟲[20]及亞洲玉米螟[21]的斗爭行為研究方法,并結(jié)合實際觀察結(jié)果記錄具體的斗爭行為特征。具體試驗方法如下:草地貪夜蛾和東方黏蟲4齡、5齡及6齡幼蟲蛻皮12 h后先饑餓處理2 h,然后在無食物供給的實驗室環(huán)境條件下,選取2頭幼蟲(不區(qū)分雌雄,試蟲 vs 對手)進(jìn)行試驗。將幼蟲置于培養(yǎng)皿(直徑6 cm,高 1.5 cm)兩側(cè);采用索尼FDR-AX45A錄像機拍攝記錄試蟲在15 min內(nèi)的斗爭行為,分類鑒定試蟲的攻擊和防御行為特征,并統(tǒng)計各類斗爭行為在15 min內(nèi)或試蟲被捕食前的發(fā)生次數(shù),計算在單位時間內(nèi)試蟲每種斗爭行為的發(fā)生次數(shù)。草地貪夜蛾和東方黏蟲的種內(nèi)及種間斗爭行為試驗組合見表1,每個組合進(jìn)行30次獨立重復(fù)試驗。
1.3數(shù)據(jù)分析
根據(jù)所拍攝的幼蟲斗爭視頻,記錄試蟲在15 min內(nèi)各類斗爭行為或其被捕食前攻擊和防御行為發(fā)生次數(shù),統(tǒng)計數(shù)據(jù)經(jīng)Excel 2010處理得到不同行為在單位時間內(nèi)的發(fā)生次數(shù),并采用SPSS 19軟件對單位時間內(nèi)各類斗爭行為發(fā)生次數(shù)進(jìn)行統(tǒng)計分析,其中,同一組合中5種斗爭行為發(fā)生頻次的差異顯著性采用單因素方差分析(α=0.05);同一種斗爭行為發(fā)生頻次在不同組合間的差異顯著性采用t檢驗分析(*:P<0.05,**:P<0.01,***:P<0.001)。采用GraphPad Prism 8軟件和Word 2010對統(tǒng)計數(shù)據(jù)進(jìn)行可視化繪圖。
2結(jié)果與分析
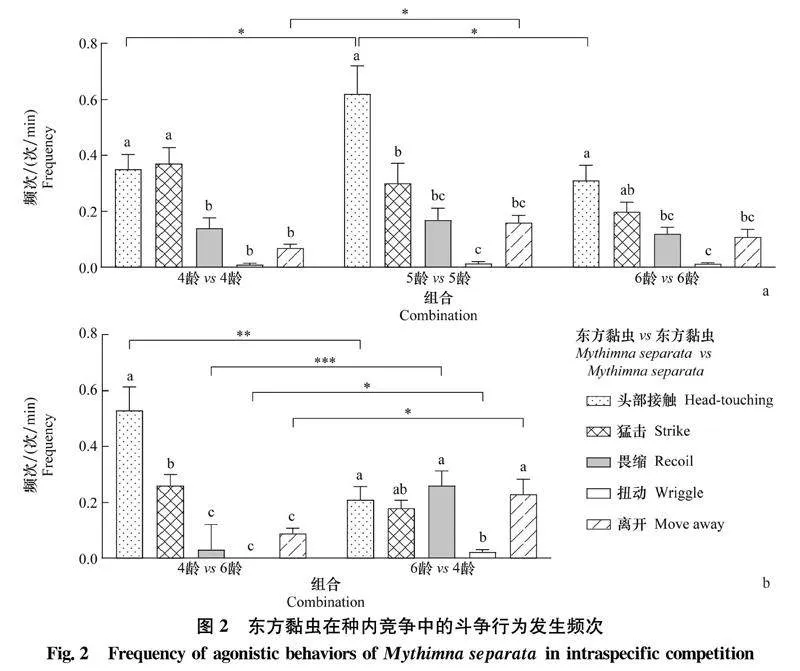
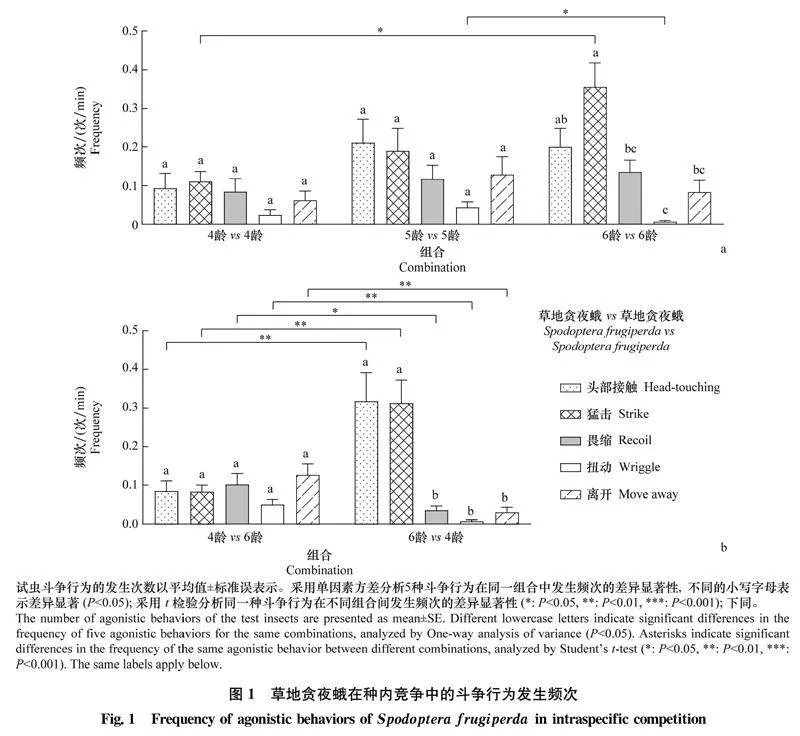
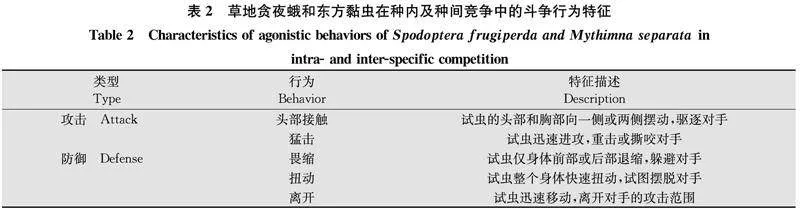
2.1草地貪夜蛾和東方黏蟲的種內(nèi)及種間斗爭行為特征草地貪夜蛾和東方黏蟲的種內(nèi)及種間斗爭行為特征見表2,主要包括5種不同特征的斗爭行為,其中攻擊行為有2種,分別為頭部接觸和猛擊;防御行為有3種,分別為畏縮、扭動和離開。
2.2草地貪夜蛾和東方黏蟲的種內(nèi)斗爭行為
2.2.1草地貪夜蛾的種內(nèi)斗爭行為
在草地貪夜蛾的種內(nèi)競爭中,4齡 vs 4齡及5齡 vs 5齡組合中草地貪夜蛾攻擊和防御行為的發(fā)生頻次無顯著差異,而6齡 vs 6齡組合中草地貪夜蛾猛擊行為的發(fā)生頻次最高,為(0.356±0.062)次/min,扭動行為的發(fā)生頻次最低,為(0.007±0.004)次/min(圖1a);6齡 vs 6齡組合中草地貪夜蛾猛擊行為的發(fā)生頻次顯著高于4齡 vs 4齡組合,其扭動行為的發(fā)生頻次顯著低于5齡 vs 5齡組合(圖1a)。在不同齡期組合中,4齡 vs 6齡組合中4齡草地貪夜蛾攻擊和防御行為的發(fā)生頻次無顯著差異,而6齡 vs 4齡組合中6齡草地貪夜蛾攻擊行為的發(fā)生頻次顯著高于防御行為,其中頭部接觸行為達(dá)(0.318±0.073)次/min,猛擊行為達(dá)(0.313±0.059)次/min(圖1b);6齡草地貪夜蛾攻擊行為的發(fā)生頻次顯著高于4齡幼蟲,其防御行為的發(fā)生頻次顯著低于4齡幼蟲(圖1b)。
2.2.2東方黏蟲的種內(nèi)斗爭行為
在東方黏蟲的種內(nèi)競爭中,4齡 vs 4齡、5齡 vs 5齡及6齡 vs 6齡組合中東方黏蟲攻擊行為的發(fā)生頻次高于防御行為,以頭部接觸行為最為顯著,分別為(0.350±0.053)次/min、(0.620±0.100)次/min和(0.310±0.054)次/min(圖2a),其中5齡 vs 5齡組合中頭部接觸行為的發(fā)生頻次顯著高于4齡 vs 4齡和6齡 vs 6齡組合,其離開行為的發(fā)生頻次顯著高于4齡 vs 4齡組合(圖2a)。在不同齡期組合中,4齡 vs 6齡組合中4齡東方黏蟲攻擊行為的發(fā)生頻次顯著高于防御行為,其中頭部接觸行為達(dá)(0.530±0.082)次/min,猛擊行為達(dá)(0.260±0.039)次/min,而6齡 vs 4齡組合中6齡東方黏蟲扭動行為的發(fā)生頻次[(0.022±0.009)次/min]顯著低于猛擊、畏縮和離開行為(圖2b);4齡東方黏蟲頭部接觸行為的發(fā)生頻次顯著高于6齡幼蟲,其防御行為的發(fā)生頻次顯著低于6齡幼蟲(圖2b)。
2.3草地貪夜蛾和東方黏蟲的種間斗爭行為
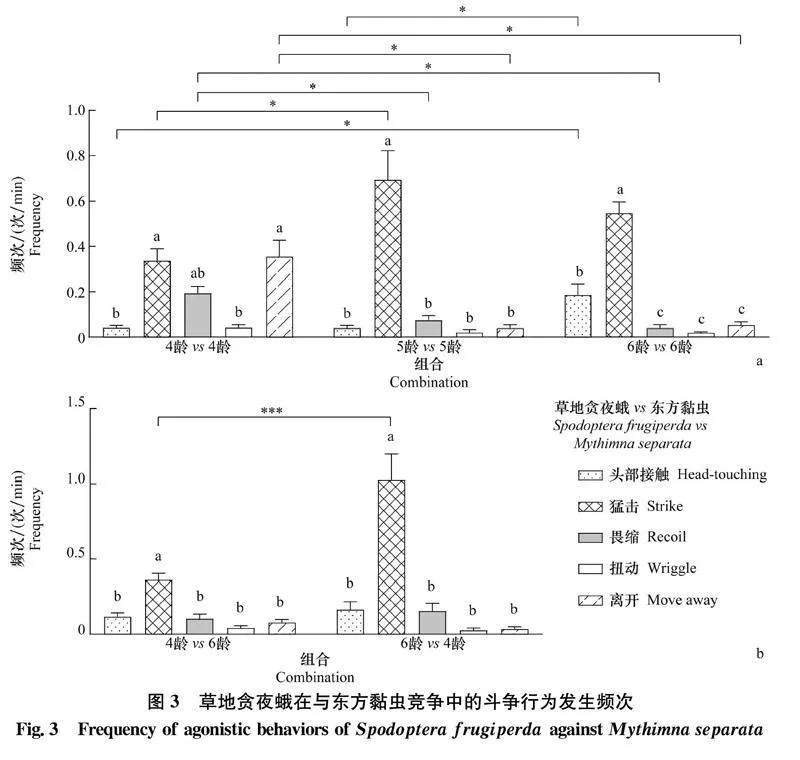
2.3.1草地貪夜蛾對東方黏蟲的種間斗爭行為
在草地貪夜蛾對東方黏蟲的種間競爭中,4齡 vs 4齡組合中草地貪夜蛾猛擊和離開行為的發(fā)生頻次相對較高,分別為(0.337±0.052)次/min和(0.355±0.072)次/min;5齡 vs 5齡組合中草地貪夜蛾猛擊行為的發(fā)生頻次顯著高于其他斗爭行為,為(0.695±0.127)次/min;6齡 vs 6齡組合中草地貪夜蛾攻擊行為的發(fā)生頻次顯著高于防御行為,其中頭部接觸行為達(dá)(0.187±0.048)次/min,猛擊行為達(dá)(0.546±0.051)次/min(圖3a);6齡 vs 6齡組合中草地貪夜蛾頭部接觸行為的發(fā)生頻次顯著高于4齡 vs 4齡和5齡 vs 5齡組合,5齡 vs 5齡組合中草地貪夜蛾猛擊行為的發(fā)生頻次顯著高于4齡 vs 4齡組合,4齡 vs 4齡組合中草地貪夜蛾畏縮和離開行為的發(fā)生頻次顯著高于5齡 vs 5齡和6齡 vs 6齡組合(圖3a)。在不同齡期組合中,草地貪夜蛾猛擊行為的發(fā)生頻次均顯著高于其他斗爭行為,其中4齡 vs 6齡組合中4齡草地貪夜蛾的猛擊行為達(dá)(0.362±0.043)次/min,6齡 vs 4齡組合中6齡草地貪夜蛾的猛擊行為達(dá)(1.026±0.172)次/min(圖3b);6齡草地貪夜蛾猛擊行為的發(fā)生頻次顯著高于4齡幼蟲(圖3b)。
2.3.2東方黏蟲對草地貪夜蛾的種間斗爭行為
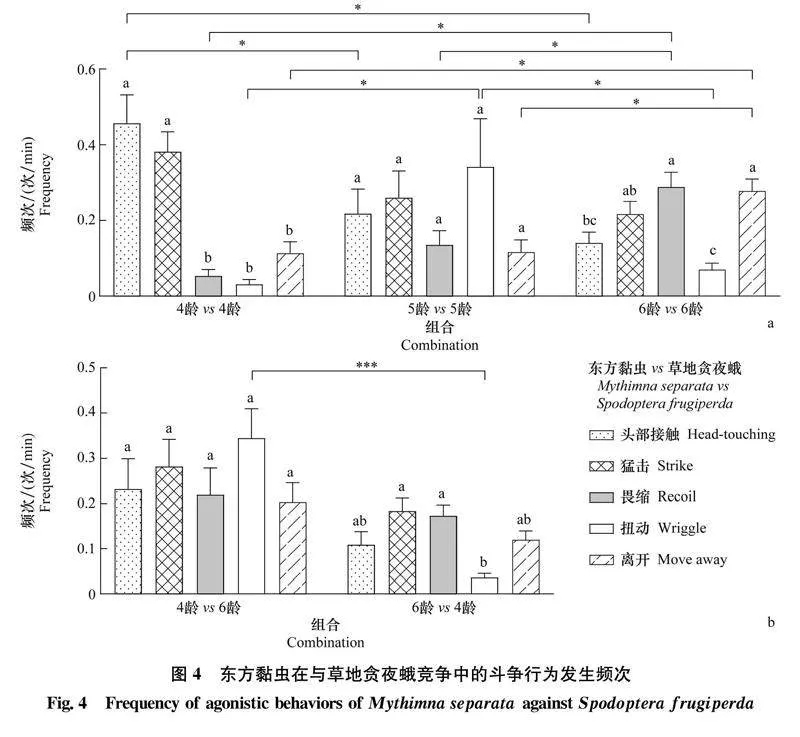
在東方黏蟲對草地貪夜蛾的種間競爭中,4齡 vs 4齡組合中東方黏蟲攻擊行為的發(fā)生頻次顯著高于防御行為,其中頭部接觸行為達(dá)(0.457±0.074)次/min,猛擊行為達(dá)(0.382±0.052)次/min;5齡 vs 5齡組合中東方黏蟲攻擊和防御行為的發(fā)生頻次無顯著差異;6齡 vs 6齡組合中東方黏蟲畏縮行為的發(fā)生頻次最高,為(0.289±0.038)次/min(圖4a);4齡 vs 4齡組合中東方黏蟲頭部接觸行為的發(fā)生頻次顯著高于5齡 vs 5齡和6齡 vs 6齡組合,6齡 vs 6齡組合中東方黏蟲畏縮和離開行為的發(fā)生頻次顯著高于4齡 vs 4齡和5齡 vs 5齡組合,5齡 vs 5齡組合中東方黏蟲扭動行為的發(fā)生頻次顯著高于4齡 vs 4齡和6齡 vs 6齡組合(圖4a)。在不同齡期組合中,4齡 vs 6齡組合中4齡東方黏蟲攻擊和防御行為的發(fā)生頻次無顯著差異,而6齡 vs 4齡組合中6齡東方黏蟲扭動行為的發(fā)生頻次最低,為(0.036±0.009)次/min(圖4b);4齡東方黏蟲扭動行為的發(fā)生頻次顯著高于6齡幼蟲(圖4b)。
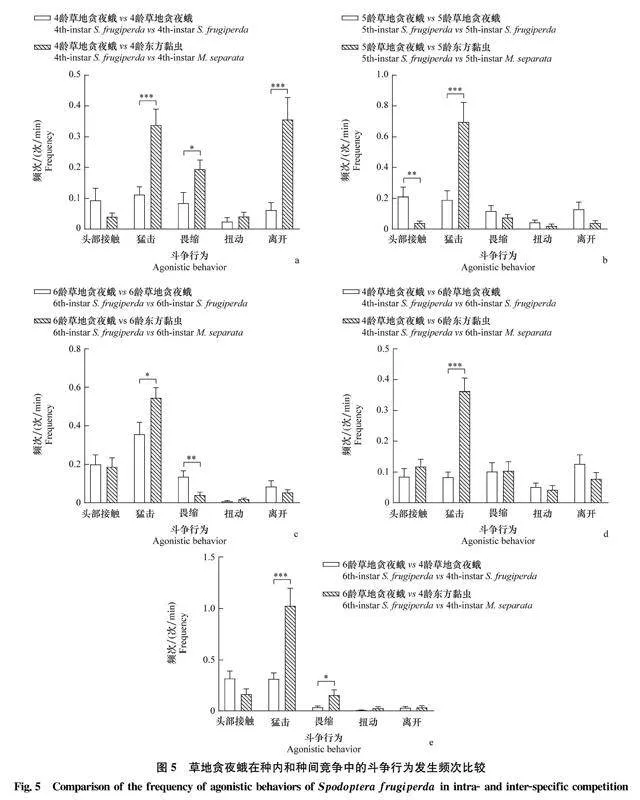
2.4草地貪夜蛾和東方黏蟲的種內(nèi)與種間斗爭行為比較2.4.1草地貪夜蛾的種內(nèi)與種間斗爭行為比較
比較草地貪夜蛾的種內(nèi)競爭及其對東方黏蟲的種間競爭發(fā)現(xiàn),在4齡 vs 4齡組合中,草地貪夜蛾的猛擊、畏縮和離開行為在種間競爭中的發(fā)生頻次顯著高于種內(nèi)競爭(圖5a);在5齡 vs 5齡組合中,草地貪夜蛾頭部接觸行為在種間競爭中的發(fā)生頻次顯著低于種內(nèi)競爭,而其猛擊行為在種間競爭中的發(fā)生頻次顯著高于種內(nèi)競爭(圖5b);在6齡 vs 6齡組合中,草地貪夜蛾猛擊行為在種間競爭中的發(fā)生頻次顯著高于種內(nèi)競爭,而其畏縮行為在種間競爭中的發(fā)生頻次顯著低于種內(nèi)競爭(圖5c)。在4齡 vs 6齡組合中,草地貪夜蛾猛擊行為在種間競爭中的發(fā)生頻次顯著高于種內(nèi)競爭(圖5d);在6齡 vs 4齡組合中,草地貪夜蛾猛擊和畏縮行為在種間競爭中的發(fā)生頻次顯著高于種內(nèi)競爭(圖5e)。
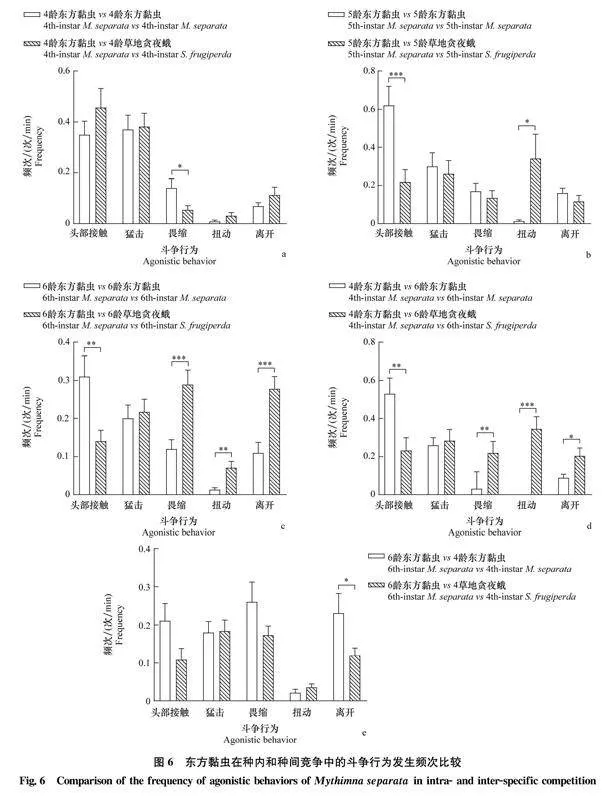
2.4.2東方黏蟲的種內(nèi)與種間斗爭行為比較
比較東方黏蟲的種內(nèi)競爭及其對草地貪夜蛾的種間競爭發(fā)現(xiàn),在4齡 vs 4齡組合中,東方黏蟲畏縮行為在種間競爭中的發(fā)生頻次顯著低于種內(nèi)競爭(圖6a);在5齡 vs 5齡組合中,東方黏蟲頭部接觸行為在種間競爭中的發(fā)生頻次顯著低于種內(nèi)競爭,而扭動行為在種間競爭中的發(fā)生頻次顯著高于種內(nèi)競爭(圖6b);在6齡 vs 6齡和4齡 vs 6齡組合中,東方黏蟲頭部接觸行為在種間競爭中的發(fā)生頻次顯著低于種內(nèi)競爭,而3種防御行為在種間競爭中的發(fā)生頻次顯著高于種內(nèi)競爭(圖6c,6d);在6齡 vs 4齡組合中,東方黏蟲離開行為在種間競爭中的發(fā)生頻次顯著低于種內(nèi)競爭(圖6e)。
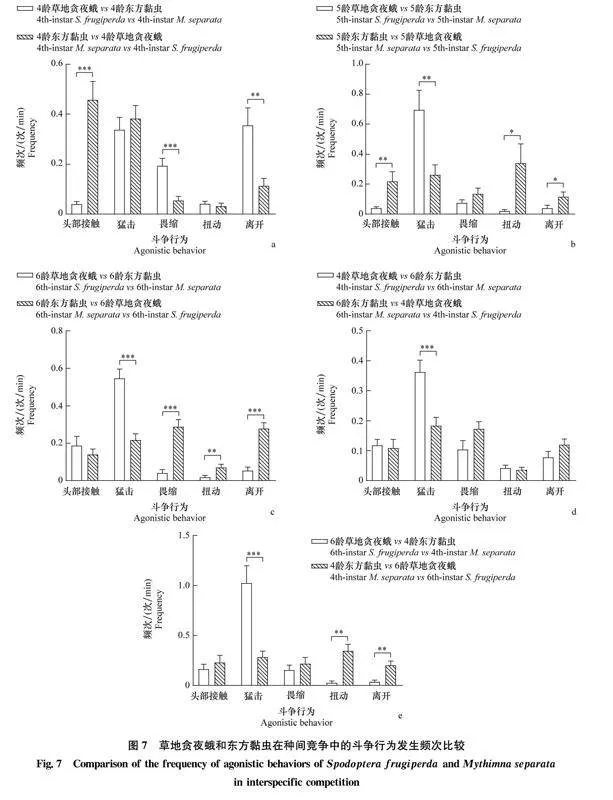
2.4.3草地貪夜蛾和東方黏蟲的種間斗爭行為比較比較草地貪夜蛾和東方黏蟲的種間競爭發(fā)現(xiàn),在同齡期的4齡組合中,草地貪夜蛾頭部接觸行為的發(fā)生頻次顯著低于東方黏蟲,其畏縮和離開行為的發(fā)生頻次顯著高于東方黏蟲(圖7a);在同齡期的5齡組合中,草地貪夜蛾頭部接觸、扭動和離開行為的發(fā)生頻次顯著低于東方黏蟲,其猛擊行為的發(fā)生頻次顯著高于東方黏蟲(圖7b);在同齡期的6齡組合中,草地貪夜蛾猛擊行為的發(fā)生頻次顯著高于東方黏蟲,其3種防御行為的發(fā)生頻次顯著低于東方黏蟲(圖7c);在4齡草地貪夜蛾和6齡東方黏蟲組合中,草地貪夜蛾猛擊行為的發(fā)生頻次顯著高于東方黏蟲,其他斗爭行為的發(fā)生頻次與東方黏蟲無顯著差異(圖7d);在6齡草地貪夜蛾和4齡東方黏蟲組合中,草地貪夜蛾猛擊行為的發(fā)生頻次顯著高于東方黏蟲,其扭動和離開行為的發(fā)生頻次顯著低于東方黏蟲(圖7e)。
3結(jié)論與討論
適當(dāng)?shù)母偁幱欣谖锓N種群數(shù)量和結(jié)構(gòu)的穩(wěn)定,而不同物種的競爭取代是種間競爭最嚴(yán)重的結(jié)果,可導(dǎo)致一個物種被迫從原棲息地遷出甚至消失[3]。在對草地貪夜蛾入侵地區(qū)的田間害蟲種群調(diào)查研究中發(fā)現(xiàn),草地貪夜蛾的種群數(shù)量越來越多,而本地鱗翅目害蟲的種群數(shù)量出現(xiàn)了不同程度減少的現(xiàn)象,表明草地貪夜蛾在與本地種群競爭中通過不同作用方式占據(jù)優(yōu)勢,其中,幼蟲的斗爭行為可能是其占據(jù)競爭優(yōu)勢的重要方式之一[2527]。在本研究中,不同齡期的草地貪夜蛾幼蟲對東方黏蟲均表現(xiàn)出了高頻次的猛擊行為,其中在6齡 vs 4齡組合中6齡草地貪夜蛾的猛擊行為高達(dá)(1.026±0.172)次/min。由此可見,草地貪夜蛾在與東方黏蟲種間競爭中表現(xiàn)出了較強的攻擊行為,同樣高頻次的攻擊行為(頭部接觸)在草地貪夜蛾與亞洲玉米螟種間斗爭行為中也有報道[21,28]。在與美洲棉鈴蟲的種間競爭中,盡管草地貪夜蛾的攻擊行為不是最頻繁的,但是攻擊強度和作用是顯著的,因此在競爭中獲得了更高的存活率[20,29]。在混合飼養(yǎng)草地貪夜蛾與東方黏蟲的研究中也發(fā)現(xiàn)了類似的結(jié)果,草地貪夜蛾的存活率遠(yuǎn)高于東方黏蟲[31]。該現(xiàn)象表明草地貪夜蛾可以通過發(fā)起主動攻擊驅(qū)離東方黏蟲,最終獲得食物資源,占據(jù)生態(tài)位成為優(yōu)勢種,草地貪夜蛾在與本地東方黏蟲種間斗爭中表現(xiàn)出的競爭優(yōu)勢預(yù)示著其具有取代東方黏蟲的可能性。類似的現(xiàn)象也出現(xiàn)在長足捷蟻Anoplolepis gracilipes[32]和西花薊馬Frankliniella occidentalis[3334]中,它們在與本地種群的種間競爭中占據(jù)一定優(yōu)勢可能是其成功入侵并定殖的主要因素之一。
此外,本研究發(fā)現(xiàn),草地貪夜蛾或東方黏蟲在不同的種內(nèi)競爭組合中所表現(xiàn)的斗爭行為特點也存在差異。對于草地貪夜蛾,僅6齡幼蟲的攻擊行為發(fā)生次數(shù)多于防御行為,其中6齡 vs 6齡組合中猛擊行為為(0.356±0.062)次/min,6齡 vs 4齡組合中頭部接觸行為為(0.318±0.073)次/min、猛擊行為為(0.313±0.059)次/min,而其他齡期幼蟲的攻擊和防御行為并沒有顯著差異。草地貪夜蛾高齡幼蟲逐漸分散為害,避免同類相食,可能是因為高齡幼蟲間的斗爭比低齡幼蟲間更強烈,尤其是高齡幼蟲對低齡幼蟲的攻擊行為更顯著[35]。然而值得關(guān)注的是,在東方黏蟲的種內(nèi)競爭中,低齡幼蟲攻擊行為的發(fā)生次數(shù)高于防御行為,以4齡 vs 4齡[頭部接觸:(0.350±0.053)次/min,猛擊:(0.370±0.057)次/min]和4齡 vs 6齡[頭部接觸:(0.530±0.082)次/min,猛擊:(0.260±0.039)次/min]組合中的4齡東方黏蟲表現(xiàn)最為顯著。該現(xiàn)象可能與聚集為害的昆蟲個體間爭奪有限的食物和空間資源有關(guān),低齡幼蟲需要更多的資源來完成發(fā)育,適當(dāng)?shù)母偁幱兄谒鼈償U(kuò)充自己的生存資源[36]。該現(xiàn)象也發(fā)生于4齡東方黏蟲與4齡草地貪夜蛾幼蟲的種間競爭中,東方黏蟲頭部接觸行為達(dá)(0.457±0.074)次/min,猛擊行為達(dá)(0.382±0.052)次/min。此外,觀察發(fā)現(xiàn),東方黏蟲4齡幼蟲的體型顯著大于同齡期的草地貪夜蛾,因此其對4齡草地貪夜蛾表現(xiàn)出較強的攻擊性可能與其體型較大有關(guān)。然而,無論在種內(nèi)或種間競爭中,東方黏蟲的攻擊行為以頭部接觸行為為主,很少具有如草地貪夜蛾般高頻的猛擊行為。頭部接觸是一種競爭強度相對較弱的攻擊行為,該行為可能更多地起警示對手的作用,并不能造成致命傷害,而猛擊行為可以對對手造成更大的傷害,甚至導(dǎo)致對手的死亡。草地貪夜蛾比東方黏蟲猛擊行為的殺傷力性更強,也可能與草地貪夜蛾發(fā)達(dá)的上顎切齒葉有關(guān)[37]。
綜上所述,外來入侵昆蟲草地貪夜蛾和本地昆蟲東方黏蟲存在明顯不同的斗爭行為特點。草地貪夜蛾對東方黏蟲會表現(xiàn)出強烈的猛擊行為,撕咬并損傷對方,從而使其更具競爭優(yōu)勢;然而,該攻擊特點在草地貪夜蛾低齡幼蟲種內(nèi)的競爭行為中并不顯著。該研究結(jié)果為解析鱗翅目害蟲幼蟲的斗爭行為提供了重要的參考依據(jù),為闡明害蟲在種內(nèi)及種間競爭中的互作關(guān)系及種群演替規(guī)律提供了理論基礎(chǔ)。
參考文獻(xiàn)
[1]NEUMANN K, PINTER-WOLLMAN N. The effect of resource availability on interspecific competition between a native and an invasive ant [J/OL]. Philosophical Transactions of the Royal Society B-Biological Sciences, 2022, 377(1851): 20210146. DOI: 10.1098/rstb.2021.0146.
[2]劉慧, 陳澤銘, 侯柏華, 等. 桔小實蠅和番石榴實蠅幼蟲取食階段的種內(nèi)競爭[J]. 環(huán)境昆蟲學(xué)報, 2015, 37(6): 11631169.
[3]REITZ S R, TRUMBLE J T. Competitive displacement among insects and arachnids [J]. Annual Review of Entomology, 2002, 47: 435465.
[4]DELONG J P, VASSEUR D A. Linked exploitation and interference competition drives the variable behavior of a classic predator-prey system [J]. Oikos, 2013, 122(10): 13931400.
[5]李志紅, BUAHOM N, 胡俊韜, 等. 實蠅科害蟲入侵來源與入侵機制研究進(jìn)展[J]. 植物檢疫, 2013, 27(3): 112.
[6]劉慧, 龔碧涯, 向敏, 等. 橘小實蠅和番石榴實蠅幼蟲之間的競爭機制[J]. 環(huán)境昆蟲學(xué)報, 2023, 45(1): 239245.
[7]石永秀, 上官超智, 王婷婷, 等. 黑豆蚜與豌豆蚜的種間競爭及密度效應(yīng)[J]. 應(yīng)用昆蟲學(xué)報, 2022, 59(4): 862873.
[8]任茂瓊, 盧永宏, 楊群芳. 3種朱砂葉螨捕食性天敵昆蟲干擾及其競爭作用研究[J]. 中國植保導(dǎo)刊, 2014, 34(8): 1417.
[9]VENKATESAN T, JALALI S K, SRINIVASAMURTHY K. Competitive interactions between Goniozus nephantidis and Bracon brevicornis, parasitoids of the coconut pest Opisina arenosella [J]. International Journal of Pest Manw0BT9LIiWPaaIZXJ4LrOZlsuny5Fn4HCgzYS7t5mb+M=agement, 2009, 55(3): 257263.
[10]BAXTER C M, BARNETT R, DUKAS R. Aggression, mate guarding and fitness in male fruit flies [J]. Animal Behaviour, 2015, 109: 235241.
[11]LEGROS J, TANG G, GAUTRAIS J, et al. Long-term dietary restriction leads to development of alternative fighting strategies [J/OL]. Frontiers in Behavioral Neuroscience, 2021, 14: 599676. DOI: 10.3389/fnbeh.2020.599676.
[12]BURNET B, CONNOLLY K, KEARNEY M, et al. Effects of male paragonial gland secretion on sexual receptivity and courtship behaviour of female Drosophila melanogaster [J]. Journal of Insect Physiology, 1973, 19(12): 24212431.
[13]NILSEN S P, CHAN Y B, HUBER R, et al. Gender-selective patterns of aggressive behavior in Drosophila melanogaster [J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(33): 1234212347.
[14]MATTHEWS R W, MATTHEWS J R. Interspecific nest parasitism by Pseudabispa paragioides, a solitary Australian wasp [J/OL]. Journal of Insect Science, 2010, 10: 160. DOI: 10.1673/031.010.14120.
[15]LIANG Qifu, LIU Tongxian. Interspecific host discrimination and intrinsic competition between Aphelinus asychis and Aphidius gifuensis in Myzus persicae [J]. Entomologia Experimentalis et Applicata, 2017, 163(3): 265271.
[16]CUSUMANO A, PERI E, VINSON S B, et al. Interspecific extrinsic and intrinsic competitive interactions in egg parasitoids [J]. Biocontrol, 2012, 57(6): 719734.
[17]YACK J E, SMITH M L, WEATHERHEAD P J. Caterpillar talk: acoustically mediated territoriality in larval Lepidoptera [J]. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(20): 1137111375.
[18]BURA V L, KAWAHARA A Y, YACK J E. A comparative analysis of sonic defences in Bombycoidea caterpillars [J/OL]. Scientific Reports, 2016, 6: 31469. DOI: 10.1038/srep31469.
[19]CHEN S, LEE A Y, BOWENS N M, et al. Fighting fruit flies: a model system for the study of aggression [J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(8): 56645668.
[20]BENTIVENHA J P F, BALDIN E L L, MONTEZANO D G, et al. Attack and defense movements involved in the inter3srDDpN3/JZ1ynj7Uel7KIAI8laju7rP6xpOUHLjjAc=action of Spodoptera frugiperda and Helicoverpa zea (Lepidoptera: Noctuidae) [J]. Journal of Pest Science, 2017, 90(2): 433445.
[21]施建琴, 郭井菲, 何康來, 等. 草地貪夜蛾和亞洲玉米螟種內(nèi)及種間的競爭行為[J]. 植物保護(hù), 2021, 47(6): 148152.
[22]SUN Xiaoxu, HU Chaoxing, JIA Huiru, et al. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China [J]. Journal of Integrative Agriculture, 2021, 20(3): 664672.
[23]張磊, 柳貝, 姜玉英, 等. 中國不同地區(qū)草地貪夜蛾種群生物型分子特征分析[J]. 植物保護(hù), 2019, 45(4): 2027.
[24]林丹敏, 黃德超, 邵屯, 等. 不同生育期玉米上草地貪夜蛾的發(fā)生為害規(guī)律[J]. 環(huán)境昆蟲學(xué)報, 2020, 42(6): 12911297.
[25]SOKAME B M, MALUSI P, SUBRAMANIAN S, et al. Do the invasive fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), and the maize lepidopteran stemborers compete when sharing the same food? [J]. Phytoparasitica, 2022, 50(1): 2134.
[26]SOKAME B M, MUSYOKA B, OBONYO J, et al. Impact of an exotic invasive pest, Spodoptera frugiperda (Lepidoptera: Noctuidae), on resident communities of pest and natural enemies in maize fields in Kenya [J/OL]. Agronomy, 2021, 11(6): 1074. DOI: 10.3390/agronomy11061074.
[27]DIVYA J, KALLESHWARASWAMY C M, MALLIKARJUNA H B, et al. Does recently invaded fall armyworm, Spodoptera frugiperda displace native lepidopteran pests of maize in India? [J]. Current Science, 2021, 120(8): 13581367.
[28]ZHAO Jing, HOFFMANN A, JIANG Yiping, et al. Competitive interactions of a new invader (Spodoptera frugiperda) and indigenous species (Ostrinia furnacalis) on maize in China [J]. Journal of Pest Science, 2022, 95(1): 159168.
[29]BENTIVENHA J P F, MONTEZANO D G, HUNT T E, et al. Intraguild interactions and behavior of Spodoptera frugiperda and Helicoverpa spp. on maize [J]. Pest Management Science, 2017, 73(11): 22442251.
[30]蘇湘寧, 李傳瑛, 黃少華, 等. 草地貪夜蛾人工飼料及飼養(yǎng)條件的優(yōu)化[J]. 環(huán)境昆蟲學(xué)報, 2019, 41(5): 992998.
[31]常向前, 呂亮, 許冬, 等. 草地貪夜蛾(Spodoptera frugiperda)與東方粘6f8479a3ce206ec8eb193464b45e80d9babe415e506bd8d3a3ca6b070f5b6e37蟲(Mythimna separata)種間競爭的室內(nèi)模擬研究[J]. 湖北農(nóng)業(yè)科學(xué), 2022, 61(6): 6165.
[32]呂曉艷, 劉霞, 張媛. 西雙版納入侵長足捷蟻與土著黃猄蟻的種間競爭[J]. 昆蟲學(xué)報, 2021, 64(10): 11961204.
[33]禹云超, 郅軍銳, 曾廣, 等. 入侵種西花薊馬與其它昆蟲的種間競爭[J]. 環(huán)境昆蟲學(xué)報, 2020, 42(1): 94100.
[34]WU Shengyong, XING Zhenlong, MA Tiantian, et al. Competitive interaction between Frankliniella occidentalis and locally present thrips species: a global review [J]. Journal of Pest Science, 2021, 94(1): 516.
[35]YASUDA H, EVANS E W, KAJITA Y, et al. Asymmetric larval interactions between introduced and indigenous ladybirds in North America [J]. Oecologia, 2004, 141(4): 722731.
[36]BOLNICK D I. Intraspecific competition favours niche width expansion in Drosophila melanogaster [J]. Nature, 2001, 410(6827): 463466.
[37]ZHAO Hailong, ZHU Zeng, XING Gaoliang, et al. Deformed mediated larval incisor lobe development causes differing feeding behavior between oriental armyworm and fall armyworm [J/OL]. Insects, 2022, 13(7): 594. DOI: 10.3390/insects13070594.
(責(zé)任編輯:楊明麗)
