紅霉素對蔬菜萌發的脅迫效應及土壤環境風險表征
2024-11-09 00:00:00石禮虎田書磊吳宗儒周睫雅吳騫王嵯周秀艷吳昊
農業環境科學學報 2024年9期
摘要:為健全紅霉素的陸生生物毒性數據并對紅霉素在土壤環境中的風險進行表征,本研究選用生菜、油菜、油麥菜和白菜為供試蔬菜進行紅霉素不同濃度下的水培試驗和盆栽實驗,探究了紅霉素對蔬菜種子萌發的脅迫效應和機制,通過ECOTOX數據庫搜集相關數據,得出紅霉素在淡水環境和土壤環境中的預測無效應濃度(PNEC),并根據盆栽實驗結果對紅霉素進入土壤后的環境風險進行了表征。結果表明:在紅霉素脅迫下,供試4種蔬菜種子的發芽率無明顯變化,根長均受到顯著抑制;紅霉素脅迫抑制了蔬菜種子的相對吸水量,促進了膜滲透性和幼苗體內的過氧化氫酶活性;供試4種蔬菜對紅霉素的敏感程度為油菜,生菜,油麥菜,白菜,半抑制濃度依次為29.387、64.075、69.255 mg·L-1和127.681 mg·L-1,紅霉素在淡水和土壤中的PNEC分別為0.2 μg·L-1和0.11 mg·kg-1,當盆栽土壤中的紅霉素含量超過250 mg·kg-1時土壤環境風險較高。研究表明,根長可作為紅霉素的毒性敏感指標,油菜可作為紅霉素毒性指示作物,紅霉素影響蔬菜種子萌發的主要機制為滲透脅迫,紅霉素菌渣肥料化產物進入土壤環境中的最高紅霉素含量應控制在250 mg·kg-1以內。
關鍵詞:紅霉素;蔬菜;脅迫效應;影響機制;風險表征
中圖分類號:X503.231;X820.4 文獻標志碼:A 文章編號:1672-2043(2024)09-2022-10 doi:10.11654/jaes.2023-1073
近年來,抗生素類污染物的環境風險防范受到世界各國的高度關注。我國也相繼發布了《新污染物治理行動方案》(國辦發[2022]15號)、《遏制微生物耐藥國家行動計劃(2022-2025年)》、《重點管控新污染物清單(2023年版)》等一系列文件,并將抗生素菌渣列為重點管控對象。我國每年產生的抗生素菌渣近1×107,其主要環境風險是殘留的抗生素進入環境中可能誘導抗性基因產生,從而影響生態環境和人體健康。目前,已經有大量研究表明在地下水沉積物、土壤、水體、污水處理廠、蔬菜和肉類中檢測到各類抗生素和抗生素抗性基因(ARGs)。《國家危險廢物名錄》規定,抗生素菌渣應按照危險廢物進行處置。但抗生素菌渣高含水率、高抗生素殘留的特性使其填埋和焚燒成本高昂且處理難度較大,從而對生物制藥行業造成了嚴重的影響。目前國內外對抗生素菌渣中殘余抗生素處置技術的研究多集中在生物處理方向,例如:動物糞便和青霉素菌渣堆肥處置后,絕大部分殘留青霉素得到了去除;青霉素菌渣與污水污泥的協同處置結果也顯示青霉素殘留可以被有效去除;林可霉素與糠醛渣的協同發酵處理也可以有效降解殘余的林可霉素。盡管目前的研究表明生物處理法具有低成本和有效去除殘余抗生素的優勢,但其資源化產物進入環境后是否會造成耐藥菌傳播風險還需要進一步評估。
本課題組前期的研究發現,無害化處理可將紅霉素菌渣中殘留的抗生素降至檢出限以下,有機質、總養分和含水率分別為77.2%、9.62%和4.63%,符合《有機肥料》(NY/T 525-2021)中的技術指標要求,重金屬As、Hg、Pb、Cd、Cr的含量分別為3.6、0.06、11.6、0.5、76 mg·kg-1,滿足NY/T 525-2021中限量指標的要求,表明抗生素菌渣的肥料化利用是較為可行的利用方式。但目前關于抗生素菌渣肥料化的無害化利用處置污染控制技術規范和有機肥中抗生素殘留限值的標準缺乏。為此,中國環境科學研究院和抗生素菌渣國家工程技術中心已在生態環境部和國家標準化管理委員會立項了青霉素、頭孢菌素以及紅霉素等典型大宗抗生素菌渣作為有機肥基料利用處置污染控制技術規范和肥料基料產物技術要求等標準。抗生素對陸生生物的毒性作用是抗生素菌渣肥料化利用相關標準與技術規范制定需要研究的重要內容。現階段,國內外對于該方面的研究主要集中在通過種子發芽試驗考察不同類型和濃度抗生素對植物的脅迫作用,綜合考慮抗生素脅迫對植物種子萌發和幼苗生長發育的影響,并以此評估其毒性作用。但抗生素濃度與類型、植物類型以及植物的不同部位對于抗生素脅迫作用的響應各不相同。Pan等的研究表明抗生素脅迫對于植物的生長發育具有低劑量促進、高劑量抑制的雙相作用;Farkas等在四環素對大豆和玉米生長發育的研究中發現,四環素脅迫下大豆的產量受到了抑制,而玉米的產量卻得到促進;Pan等的研究發現抗生素對于植物種子萌發的影響小于對其根/芽長的影響;An等發現在抗生素脅迫下,根長的抑制率高于芽長抑制率;而Jin等發現磺胺類抗生素對于芽長的抑制效果要強于對根長的抑制效果。同時,課題組通過相關文獻和美國生態毒理數據庫收集了大環內酯類(包括紅霉素)的水/陸生生物的毒性效應數據,發現紅霉素的陸生生物毒性效應數據較少,且毒性作用機理不清,難以支撐后續紅霉素菌渣肥料化產物的可行性研究,因此亟需相關研究進行填補數據空白以及對紅霉素進入土壤環境后的風險進行表征。
本研究選用紅霉素為試驗污染物,以生菜、油菜、油麥菜和白菜4種常見蔬菜作為試供植物,以種子發芽率、幼苗根長和芽長為指標考察了不同濃度紅霉素對常見蔬菜種子萌發期間的脅迫效應;以過氧化氫酶(CAT)活性、相對吸水量和膜滲透性為指標探究了紅霉素對試供植物的影響機制;對紅霉素濃度和幼苗根長抑制率進行擬合,考察供試植物對紅霉素的敏感程度及半數致死濃度,并根據敏感程度,選用最為敏感的作物在不同濃度紅霉素作用下進行盆栽試驗,考察了作物成熟后盆栽土壤的環境風險。以期為紅霉素類抗生素菌渣肥料化利用相關標準與風險閾值的建立提供數據參考和科學依據。
1 材料與方法
1.1 試驗材料
試驗所用紅霉素純品由川寧生物技術有限公司提供,純度大于99%。試供蔬菜為生菜(Lactuca sati-va var. ramosa Hort.)、油菜(Br,assica rapa var. chinensis Kitamura)、油麥菜(Lactuca sativa var. longifoliaf Lam)和白菜(Brassica rapa var. glabra Regel),種子均購自淘寶,品牌為農科院京研。CAT活性由試劑盒(紫外分光光度法)測定,品牌為索萊寶。電導率由便攜式電導率儀測定,品牌為雷磁。盆栽土壤取自連續5a未施用有機肥的表層耕作土壤,土壤類型為棕壤,中性偏堿。盆栽試驗所用蛭石粒徑為2-5 mm,購自淘寶,品牌為百土集。
1.2 試驗設計
試劑配制:取1g紅霉素純品粉末,用95%無水乙醇溶解后用去離子水在1000 mL棕色容量瓶中定容,配制為1 000 mg·L-1的紅霉素母液,隨后根據在0-2 000mg-L條件下開展的預實驗結果,在根長抑制率為10%-60%的區間內設置7種紅霉素濃度(20、50、150、200、300、400、600 mg·L-1),按照所需紅霉素的濃度對母液進行稀釋后保存于棕色廣口瓶中等待后續使用。
種子發芽試驗:試供種子用10% H2O2浸泡消毒10 min后用去離子水反復沖洗并濾干。取兩張濾紙修剪后放入一次性培養皿中,使其邊緣貼合,隨后加入3 mL紅霉素溶液作為試驗組,對照組則加入等量去離子水,每個培養皿中整齊放入15粒蔬菜種子,每個處理設置3組平行。試驗在恒溫恒濕培養箱中進行,設置溫度為25℃,濕度為60%,培養7d后結束試驗,測定種子發芽率、幼苗根長、芽長及CAT活性等指標。
吸水量和膜滲透性測定:取10 g左右試供種子記錄干質量,放入燒杯后加入20 mL紅霉素溶液,此為試驗組,對照組加入等量去離子水。種子在培養箱中培養48 h后取出,平鋪在濾紙上自然晾干至表面無水漬后稱得濕質量。隨后將種子用去離子水沖洗5-6次后放入干凈的燒杯中,加入50 mL去離子水,在培養箱中培養6h后取出測定電導率得到電導率1,使用電爐加熱煮沸,待冷卻后測得電導率2。計算后得出相對吸水量和相對電導率,分別表征種子萌發初期的吸水量和膜滲透性,其中,相對吸水量=(濕質量—干質量)/濕質量,相對電導率=電導率1/電導率2。
盆栽實驗:土壤風干后過4 mm篩子并去除殘留蟲卵及植物根莖后裝盆,花盆規格為23.5 cm×4 cm,每盆裝入風干土壤1 kg并混入50 g蛭石以增加土壤透氣性和保水性且防止植物爛根,將土壤與蛭石混合均勻后每盆加入300 mL水完全浸潤土壤并平衡24h,根據種子發芽試驗中敏感性結果,選擇油菜作為盆栽作物。育苗過程選取純蛭石作為育苗基質,選取大小均一、顆粒飽滿的油菜種子,浸泡15 min后,均勻點入128孔育苗盤中,每孔點入3粒種子,置于(25±1)℃光照培養箱中,每隔24 h觀察育苗盤底部水量,加適量水以維持油菜幼苗的正常生長,培養7d直至幼苗長出第4片葉,每盆加入相應濃度的紅霉素標準品并與土壤混勻后進行移栽,每盆均勻移栽3顆,每組處理設置5個平行。盆栽實驗于中國環境科學研究院玻璃溫室內進行,溫度為20-30℃,濕度為8%- 12%,自然光照。為保證光照的均勻性,每2d隨機調換花盆的位置。每2d澆1次水,保證每盆的澆水量一致,盆栽的時間周期為60 d,整個生長期不使用殺蟲劑。在第60天油菜成熟后進行土壤取樣,并根據課題組之前的研究結果進行紅霉素含量的測定。具體分組情況見表1。
1.3 統計分析
使用SPSS 25軟件對實驗數據進行單因素方差分析和顯著性檢驗,條形圖和點線圖使用Origin 2019繪制,Spearman相關系數圖使用R 4.1.1中的ggplot2和ggpubr包繪制。
2 結果與討論
2.1 紅霉素脅迫對蔬菜種子發芽率的影響
如表2所示,在不同濃度紅霉素處理后白菜、生菜、油菜和油麥菜4種蔬菜的種子發芽率總體與空白對照組無顯著差異(P>0.05),僅白菜和油菜種子分別在紅霉素濃度為400 mg·L-1和600 mg·L-1下發芽率顯著低于空白對照組(P<0.05),種子整體發芽率均大于86.67%。該結果表明紅霉素脅迫對4種蔬菜種子的發芽率無顯著影響。這與Bellino等進行氯霉素、螺旋霉素、大觀霉素和萬古霉素4種抗生素對番茄種子萌發影響的研究結果相似,即抗生素脅迫對種子萌發無顯著影響,該現象也在Pino等和Pan等的研究中出現。產生這種現象的原因一方面可能是蔬菜種子發芽較快(24-48 h),在紅霉素還未對其產生影響時就已經萌發,因此對發芽率無顯著影響;另一方面則可能是由于種子在未萌發時有種皮保護,紅霉素被吸附在種皮表面,阻礙了紅霉素與胚根的直接接觸,進而對種子萌發影響較小。種子在萌發期間,其內部會發生各種酶促反應,在此期間,激活的各組酶對種子內儲存的各種能量物質產生作用,進行分解活動,為后期幼苗的生長發育提供初始能量。這也就意味著,盡管在紅霉素脅迫下種子可以正常萌發,但紅霉素脅迫對后續幼苗的生長發育是否存在毒性效應還不能確定。
2.2 紅霉素脅迫對蔬菜幼苗根長和芽長的影響
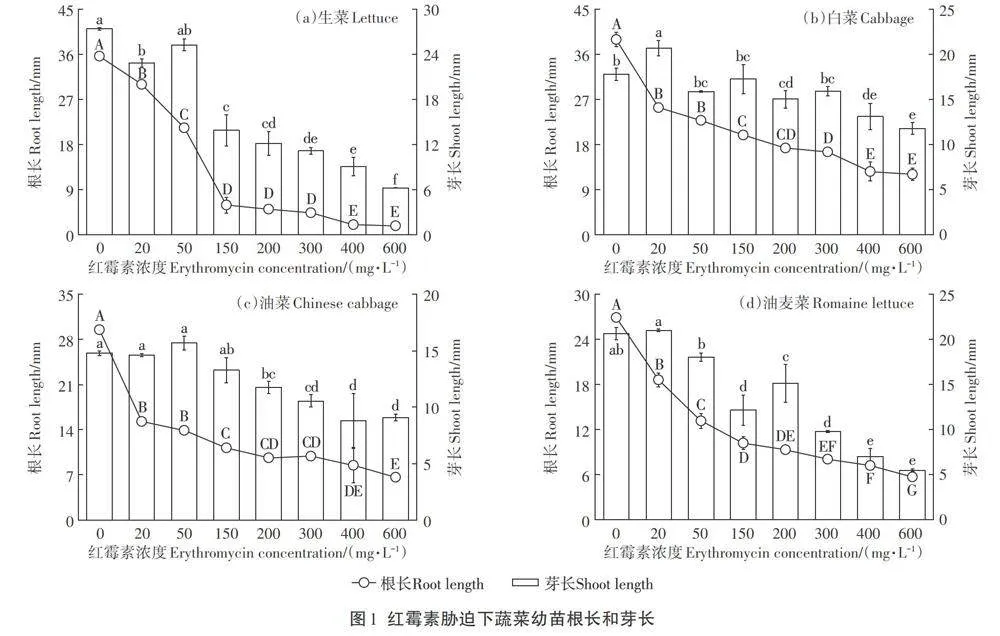
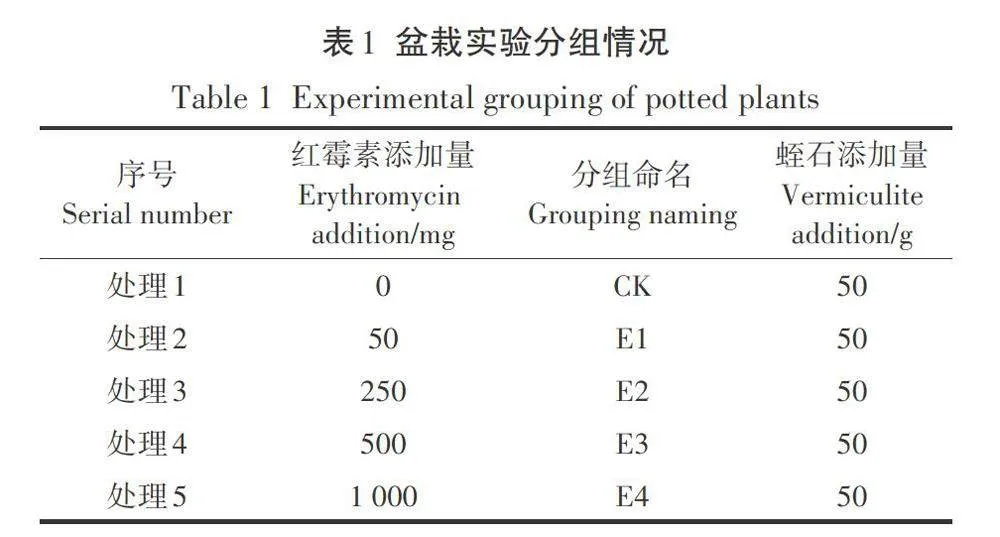
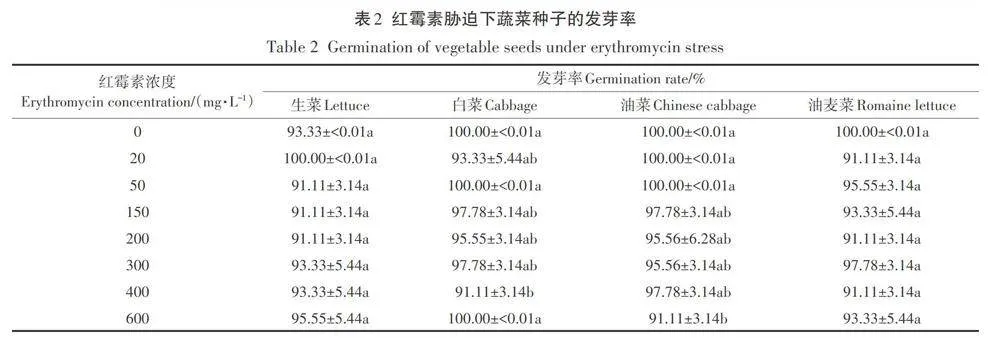
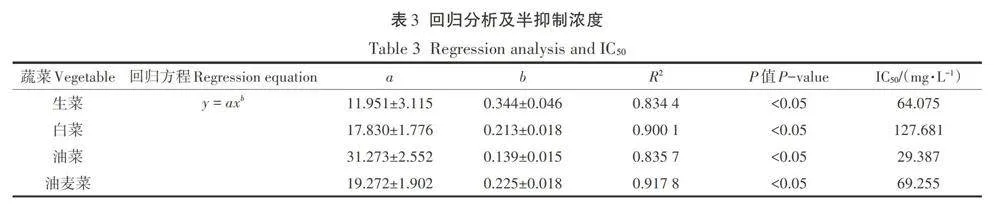
紅霉素脅迫下4種蔬菜幼苗的根長和芽長如圖1所示。從圖中可以看出,隨著紅霉素濃度的增加,4種蔬菜幼苗的根長均受到明顯的抑制作用,白菜、生菜、油菜和油麥菜的最大根長抑制率分別為68.87%、95.16%、74.27%和78.80%。
根是種子在萌發過程中最先生長出的部位,可通過輸導組織將環境中的水分、無機鹽、有機物等營養物質和養分輸送給其他部位,因此,根也是與環境接觸最為緊密的部位,環境的影響也最先作用到根部。在本實驗中,紅霉素的脅迫作用也直接作用于根部,通過對根部抗氧化防御系統和細胞分裂產生影響進而抑制根伸長。此外,由于種子的萌發都發生在暗生芽期,而在此期間根部是生長最為快速的器官,對環境中的抗生素最為敏感,所以導致了在紅霉素脅迫下,根長受到明顯的抑制作用。Bellino等在不同類型抗生素對番茄種子萌發影響的研究中也發現在抗生素脅迫下番茄的根系發育會受損。Gomes等在環丙沙星和除草劑對玉米根系發育影響的研究中進一步發現抗生素的添加會通過影響根系細胞有絲分裂和活性氧(ROS)起到對根系生長的抑制作用。而蔬菜幼苗芽長的抑制效果不如根長主要是由于在幼苗生長期間所需要的水分由胚根向胚軸運輸,而紅霉素濃度會沿水勢方向遞減,這就導致胚軸所接觸的紅霉素濃度較低,因此對芽長的影響不如根長明顯。
2.3 紅霉素脅迫對蔬菜種子萌發和生長發育的影響機制
在非生物逆境脅迫下種子的萌發調控機制主要有干旱脅迫、鹽脅迫、高溫脅迫、蔭蔽脅迫、滲透脅迫和淹澇脅迫,之前關于植物種子在金霉素、環瑞沙星等藥物脅迫下的萌發調控機制研究均表明鹽脅迫和滲透脅迫是主要的調控機制,具體表現為鹽脅迫和滲透脅迫對ROS和細胞代謝過程的影響。其中,ROS是一種在外界脅迫下對植物生長發育起到調節作用的信號分子,與ROS清除酶共同作用,保證其在植物體內的平衡以正常參與植物的生長發育過程。而細胞代謝過程對種子的正常萌發也至關重要,在非生物逆境脅迫下,種子可以通過調節代謝過程和能量的物質儲存方式應對脅迫。因此,本節以供試4種蔬菜種子體內CAT活性、細胞膜滲透性和吸水量為指標,探究了紅霉素脅迫對蔬菜生長發育的影響機制,其中,CAT活性用以反映植物體內的ROS水平,細胞膜滲透性和吸水量用以反映紅霉素脅迫對細胞代謝過程的影響。
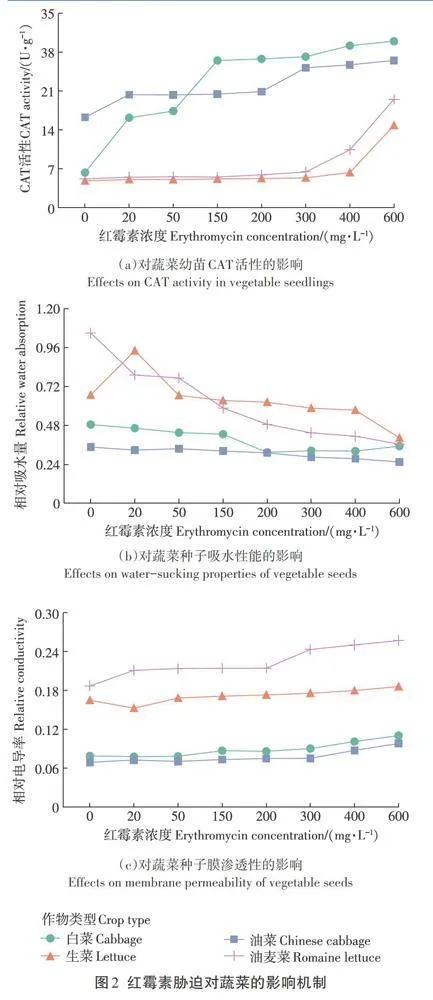
CAT是植物體內最重要的一種ROS清除酶,通過催化H2O2形成H2O而阻止H2O2在體內的過量積累進而降低ROS對細胞膜的過氧化作用,其活性反映了植物生長條件的好壞。不同濃度紅霉素脅迫下4種蔬菜幼苗體內CAT活性的變化如圖2(a)所示。從圖中可以看出4種蔬菜幼苗體內CAT活性在整體上都隨著紅霉素濃度的增加而升高,但對于不同類型的蔬菜,CAT活性的增加幅度和增速不盡相同。對于白菜和油菜,兩者體內的CAT活性在紅霉素濃度為20mg·L-1時就開始明顯增加;對于生菜和油麥菜,兩者體內的CAT活性在紅霉素濃度較低時均在5 U·g-1附近波動,隨著紅霉素濃度增加到300 mg·L-1時開始明顯增加。由于白菜和油菜同屬于十字花科十字花目蕓薹屬,而生菜和油麥菜同屬于菊科菊目萵苣屬,因此推測不同的植物種屬導致了在紅霉素脅迫下CAT活性變化的差異。如Chen等在金霉素對大豆和玉米發育影響的研究中發現金霉素可以促進玉米體內CAT活性增強而對大豆體內的CAT活性無明顯影響,該結果與不同類別植物中營養物質的存儲方式和包括胱甘肽S-轉移酶在內的多種調控因子的活性相關。盡管不同蔬菜幼苗中CAT活性的變化不盡相同,但整體而言,均隨著紅霉素濃度的增加而上升,表明紅霉素脅迫會通過增加CAT活性破壞蔬菜中ROS濃度信號的傳遞,進而抑制蔬菜的生長發育。該結果也與Gomes等在環丙沙星對玉米根系發育的影響研究中的結果相似,即抗生素的添加會促進植物體內CAT和APX(抗壞血酸過氧化物酶)的活性,進而抑制根伸長。
4種蔬菜種子在不同濃度紅霉素脅迫下吸水量的變化如圖2(b)所示。從圖中可以看出,4種蔬菜種子的吸水量整體上隨著紅霉素濃度的增加而具有降低的趨勢。在種子萌發初期,種子的吸水性能是影響種子呼吸代謝的重要因素,對種子萌發和后續根莖的發育至關重要,而紅霉素的添加會對植物種子初期吸水量產生抑制作用,產生這一結果的原因可能是種子初期是依賴于細胞質電位的被動吸水,而添加的紅霉素影響了細胞膜內外的電位平衡,最終影響種子的吸水量,這一原因也在圖2(c)中得到了證實。如圖2(c)所示,隨著紅霉素濃度的增加,4種蔬菜種子的相對電導率也增加。該結果表明,隨著紅霉素濃度的增加,細胞質膜受到影響,胞內可溶物有所滲漏,細胞內外電位平衡遭到破壞,引起滲透失衡,進而造成滲透脅迫。而滲透脅迫會阻礙種子的正常生長發育,并會限制植物對環境中水和營養物質的吸收,這就解釋了在本研究中,4種蔬菜種子相對吸水量隨環境中紅霉素濃度的增加而降低。由細胞膜內外電位失衡所引發的滲透應激反應會促使植物產生氧化應激反應,進而導致植物體內ROS的積累速率超出CAT的清除能力,這也解釋了紅霉素脅迫會導致植物體內CAT活性增加的原因。
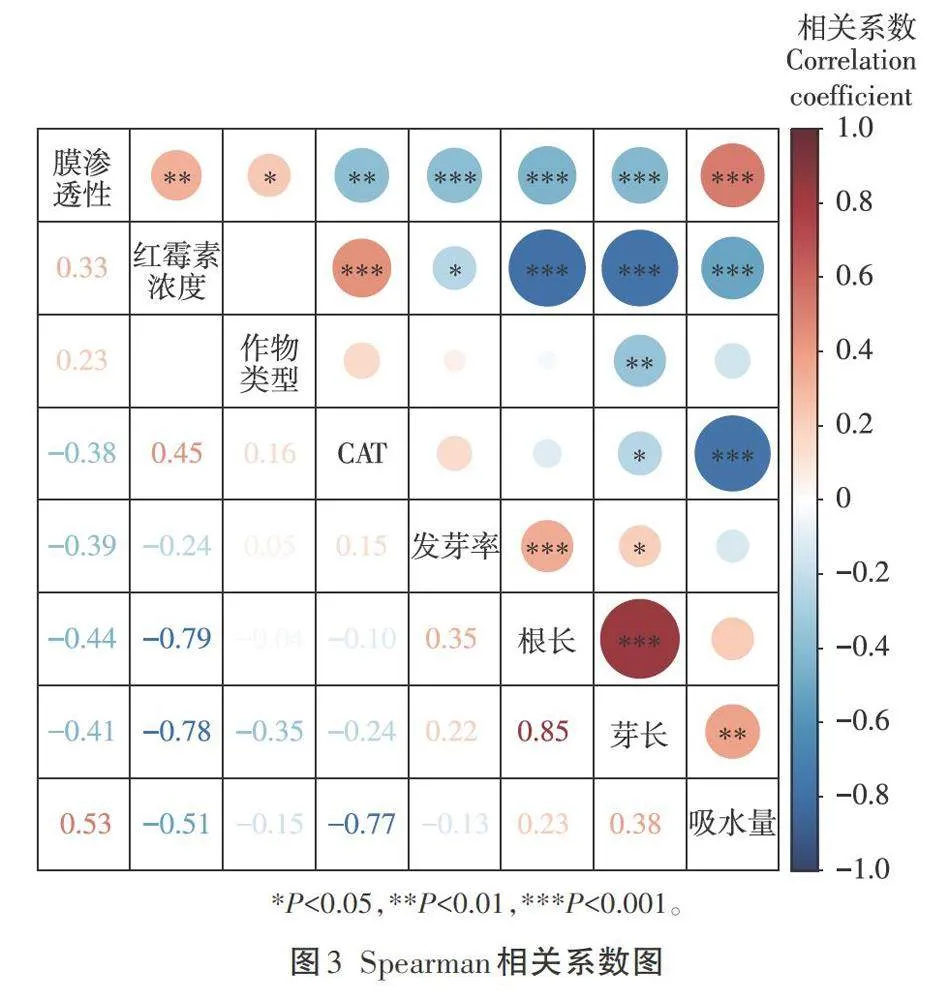
上述結果表明,在紅霉素脅迫下,蔬菜的CAT活性、吸水量和膜滲透性對其生長發育均有不同程度的影響,即紅霉素脅迫對于蔬菜生長發育的影響是一個多因素的復雜過程。為此,有必要進一步探究上述影響因素之間的潛在聯系及主要作用機理。如圖3的Spearman分析結果所示,紅霉素濃度與根長呈極顯著負相關性(r=-0.79,P<0.001),表明在種子萌發初期,根長對于環境中紅霉素濃度的變化最為敏感,可作為敏感性指標。膜滲透性與所有指標均存在顯著相關性,其中,與吸水量之間的相關性最強且極顯著(r=0.53,P<0.001),同時發現與吸水量最為相關的指標為CAT活性(r=-0.77,P<0.001),表明膜滲透性是影響蔬菜種子萌發最主要的因素。植物在滲透脅迫下體內會產生解毒信號,而該信號的功能就包括激活ROS去除酶和其他解毒蛋白。這也表明紅霉素對于植物的影響機制在本質上可能是一種滲透脅迫,具體表現為紅霉素影響細胞膜內外電位平衡,進而引發滲透脅迫,而在滲透脅迫下植物體內的多種解毒蛋白被激活,進一步誘發多種應激反應,最終影響植物的生長發育。
2.4 不同蔬菜對于紅霉素脅迫的敏感程度
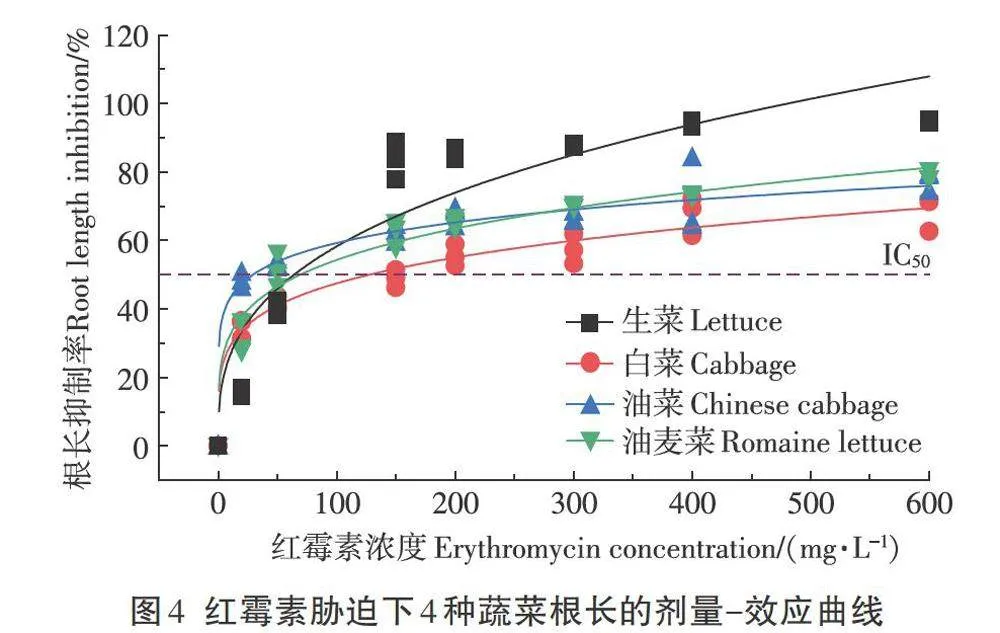
根據前文中根長對紅霉素濃度變化最為敏感的結論,選用根長為敏感性指標,建立根長抑制率和紅霉素濃度的響應曲線(圖4),并得到回歸方程和半抑制濃度(IC50)(表3)。從表3可知,4種蔬菜根長抑制率與紅霉素濃度的劑量—效應曲線滿足Allometricl模型,R2均大于0.8,表明該模型對數據的擬合度較好,可以用作分析。
目前,IC50常用作評價污染物對植物的生態毒性,其值大小與植物對污染物的敏感程度呈負相關關系。從表3可以得出生菜、白菜、油菜和油麥菜的ICso分別為64.075、127.681、29.387 mg·L-1和69.255mg·L-1,油菜的IC50最小,即油菜對于紅霉素濃度的變化最為敏感,隨后依次為生菜、油麥菜和白菜。不同蔬菜對紅霉素濃度變化的敏感程度不同可能是由于不同蔬菜中營養物質的存儲狀態不一致,如萵苣屬蔬菜在體內主要通過淀粉粒儲存能量,而蕓薹屬蔬菜并不具有淀粉粒,這就可能會導致紅霉素在蔬菜中作用的靶分子不同,進而導致不同的蔬菜對紅霉素的敏感程度不同,但具體作用機制還需在未來做進一步探究。而Pan等的研究發現四環素(TC)、磺胺甲嘧啶(SMZ)、諾氟沙星(NOR)和氯霉素(CAP)對小油菜的IC50分別為14.4、157、49.4 mg·L-1和204 mg·L-1,即不同類型抗生素對同種蔬菜的毒性也不盡相同,這主要是由于抗生素對植物的毒性主要與其分子結構和穩定性相關。總體而言,抗生素和種植作物的類型均會對毒性產生影響,后續應完善大宗抗生素對常見作物的毒性數據,以期在抗生素菌渣肥料化利用的過程中合理指導施加量。
上述結果表明,對紅霉素的敏感程度為油菜>生菜>油麥菜>白菜,因此在后續紅霉素生態毒性的相關研究中可選用油菜作為敏感作物。在紅霉素菌渣肥料化相關標準中應針對不同作物的耐受性做細分要求,按照不同作物對紅霉素的敏感程度合理指導施肥量。
2.5 紅霉素生態風險表征
從ECOTOX數據庫(http://cfpub.epa. gov/ecotox/)中檢索紅霉素水生及陸生生物的急性/慢性毒性數據。根據歐盟適用于現有化學物質的風險評價技術指南(TGD)中規定的毒性數據篩選原則進行篩選數據,數據的處理原則參照鞏秀賢的方法,同物種不同毒性終點的數據取最小值,同物種同毒性終點的數據取幾何平均值。
2.5.1 淡水環境中紅霉素預測無效應濃度(PNEC水)
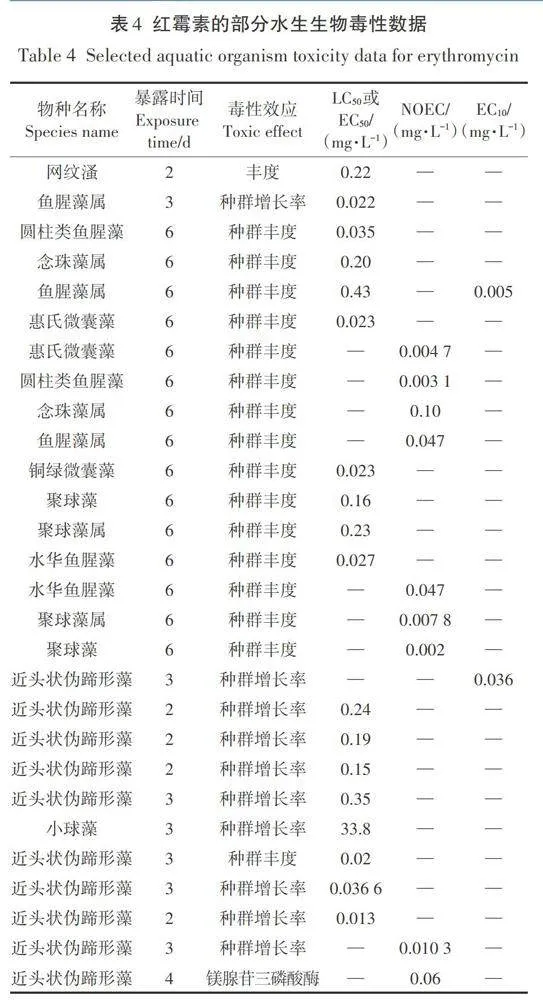
根據上述篩選原則,在數據庫中共篩選出105個紅霉素對水生生物的毒性數據,其中生物物種類別包括魚類(條紋鱸、斑馬魚、青鮪魚)、甲殼類(大型潘、多刺裸腹潘、長刺潘、南美白對蝦)、藻類(魚腥藻、念珠藻、惠氏微囊藻、銅綠微囊藻、聚球藻、鐮形纖維藻、近頭狀偽蹄形藻、小球藻)和高等植物(青萍、浮萍)。其中部分急慢性毒性數據見表4。
因為在篩選出的105個數據中共有44個慢性無觀測效應濃度(NOEC),共包含了3個營養級別(藻、潘、魚)的至少3個物種的長期ⅣOEC,所以采用評估系數法計算PNEC水,對應評估系數選用10。評估系數法是根據評價終點除以評估系數進而得到PNEC,對于多個物種多項評價終點時,取最低值除以評估系數得到PNEC。在比較藻、潘、魚3個營養級別的長期數據后發現近頭狀偽蹄形藻3d以種群豐富度為毒性終點的NOEC=0.002 mg·L-1是藻類最低值,魚腥藻和近頭狀偽蹄形藻的EC10值(10%最大效應濃度)分別為0.005 mg·L-1和0.036 mg·L-1,均低于ⅣOEC值,因此,PNEC水=0.002 mg·L-1/10=0.2 μg·L-1。
2.5.2 土壤環境中紅霉素預測無效應濃度(PNEC土壤)
由于可獲得的土壤中紅霉素對生物的毒性數據較少,因此采用平衡分配法計算PNEC土壤,即:PNEC土壤=K土壤-水/RHO土壤×PNEC水×1000,其中K土壤-水為土壤-水分配系數,RHO土壤為土壤的濕體積密度。使用EPI Suite V4.10軟件獲得土壤呼吸系數為25.49L·kg-1,得到土壤中紅霉素固-水分配系數為2.6 L·kg-1;按照TGD中標準環境特征參數,忽略紅霉素在水體、土壤與氣體的分配,RHO土壤采用TGD默認值1 700 kg·m-3,可以得到K土壤-水為4.1,而在2.5.1中得到PNEC水為0.2 μg·L-1。因此,根據分配法計算公式可得PNEC土壤=0.11 mg·kg-1。
2.5.3 紅霉素進入土壤后的生態風險表征
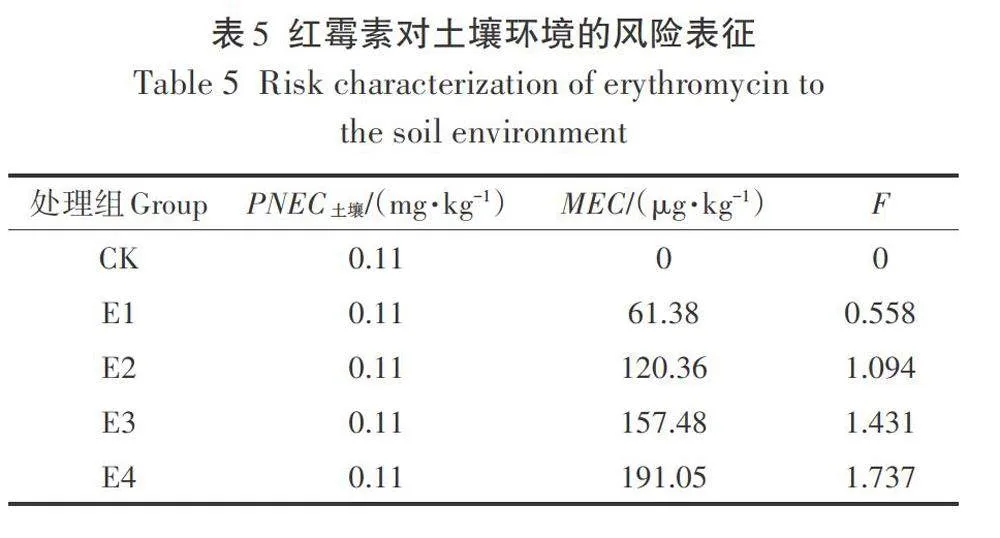
采用定量描述的方式對紅霉素的土壤生態風險進行描述,具體使用風險指數(F)進行表征,F的計算公式為:F=MECIPNEC。其中,MEC為環境中紅霉素的實測濃度,PNEC為環境中紅霉素的預測無效應濃度。紅霉素在土壤中的PNEC由2.5.2可得,為0.11mg·kg-1,MEC由盆栽實驗第60天作物成熟后的土壤中的紅霉素含量得出,具體見表5。
根據F定量描述的規定,F<0.1時,環境風險可忽略;0.1≤F≤1時,風險較低,但還需長期觀測;F>1時,環境風險較高,不可接受。從盆栽實驗數據可發現,當流入土壤環境中的紅霉素濃度為50 mg·kg-1時,在經歷60d的穩定后,F<1,土壤環境風險較低;而當流入土壤環境中的紅霉素濃度高于250 mg·kg-1時,土壤環境短期的穩定難以自發降解紅霉素,環境風險較高。因此,未來紅霉素菌渣肥料化產物進入土壤環境的濃度應控制在250 mg·kg-1以內(按肥料化產物中有效紅霉素濃度計),以降低其對環境的影響。
3 結論
(1)紅霉素對4種供試蔬菜幼苗根長的抑制作用最為明顯,對種子萌發無明顯影響。根長抑制情況可較好反映紅霉素的植物毒性及其敏感程度。
(2)紅霉素對蔬菜種子萌發的脅迫效應是一種包括氧化應激、滲透脅迫在內的復雜過程,主要影響機制為紅霉素對細胞膜內外電位平衡的破壞而引發的滲透脅迫。
(3)供試4種蔬菜對于紅霉素的敏感程度表現為油菜>生菜>油麥菜>白菜,油菜是本試驗中對紅霉素最為敏感的作物,IC50為29.387 mg·L-1。后續可將油菜作為探究紅霉素菌渣肥料化產物對土壤生態毒性影響的指示性作物。
(4)紅霉素在淡水環境和土壤環境中的預測無效應濃度分別為0.2 μg·L-1和0.11 mg·kg-1。其中,紅霉素菌渣肥料化產物進入土壤環境中的最高紅霉素濃度應控制在250 mg·kg-1以內。
(責任編輯:李丹)
