納米鐵氧化物強化牛糞厭氧發酵產甲烷
2024-11-09 00:00:00李煜葉青青瞿云山劉子墨任玉瑩于莉民李欣
農業環境科學學報 2024年5期

關鍵詞:厭氧發酵;牛糞;納米鐵氧化物;揮發性脂肪酸;宏基因組學
集約化養殖快速發展產生了大量畜禽糞污,以牛糞為例,根據中國統計年鑒(http://www.stats.gov.cn/sj/ndsj/)和農業農村部頒布的養殖畜禽糞尿產生系數(https: //www. gov. cn/zhengce/zhengceku/2018-12/31/content_5442613.htm)估算,2018年我國年產牛糞約12.8億t。若不經處理直接排放會對養殖場周圍的水體、土壤和大氣環境造成嚴重污染。厭氧發酵通過降解有機質產沼氣,目前已廣泛應用于牛糞處理。然而,產氣效率低是牛糞厭氧發酵面臨的主要問題,這是因為牛糞富含木質纖維素,其緩慢的水解效率制約了產甲烷效率。目前,研究人員采用多種方法來提高牛糞厭氧發酵產氣性能,包括:(1)與低木質素含量底物共發酵以減輕木質素對水解菌的生物抑制作用;(2)預處理破壞木質纖維素提高其生物降解性;(3)新厭氧發酵體系構建(如微生物電化學);(4)外源添加劑強化水解和產甲烷階段。
在上述方法中,外源添加劑在提升產氣性能和經濟可行性方面具有顯著優勢。其中,鐵及其化合物作為多種酶(如甲酰一甲呋喃脫氫酶、氫化酶、一氧化碳脫氫酶)的輔因子參與甲烷合成,是公認的可改善牛糞厭氧發酵性能的添加劑之一。此外,鐵還能參與厭氧微生物能量代謝途徑Wood-Ljungdahl(WL),并能作為最終電子受體改善厭氧發酵過程。
在鐵化合物中,納米鐵氧化物(Iron oxidenanoparticle,INP)因其具有高比表面積、超準磁性、易跨膜運送和低生物毒性受到研究者的廣泛關注。如Abdelsalam等添加20mg·L-1 INP,將牛糞厭氧發酵甲烷產量提升1.96倍。Farghali等開展系列實驗,證實了添加INP可將牛糞甲烷產量提升10.5%-56.9%。基于作者前期文獻調研發現,INP促進牛糞厭氧發酵相關研究主要集中于短期批式實驗,對于更接近牛糞沼氣工程實際運行情況的長周期半連續實驗鮮有報道。此外,前人研究對INP強化微生物代謝途徑這一核心作用缺乏深入分析。因此,本研究將開展以牛糞為底物的半連續試驗,并基于16S rRNA分析和宏基因組闡明INP添加對牛糞厭氧發酵產氣效率和微生物群落演替和代謝途徑的影響。研究結果以期為牛糞沼氣工程INP的使用和相關理論基礎的完善提供參考。
1材料和方法
1.1材料與實驗設計
半連續試驗在兩個完全相同的連續攪拌反應器(Continuous stirred tank reactor. CSTR)中進行,CSTR的工作容積為2.4L。其中R-Fe為添加INP的實驗組,R-C為對照組。表1列出了試驗的基本運行狀況。牛糞理化性質如下:pH: 8.59+0.03;總固體:(278+15)mg·g-1;揮發性固體:(168+12)mg·g-1;纖維素:(153.1+6.1)mg·g-1;半纖維素:(140.5+3.4)mg·g-1;C:(314.3+0)mg·g-1;H:(42.9+0.1)mg·g-1;N:(22.1+0.7)mg·g-1; 0: (615.7+0.1)mg·g-1;S:(5.1±0.2)mg·g-1。試驗開始前,兩個CSTR均以牛糞為唯一底物穩定運行100d,并具有相近的日產甲烷量(Pgt;0.05)。本試驗中有機負荷(Organic loading rate,OLR)和水力停留時間(Hydraulic retention time,HRT)分別為1g·L-1·d-1(以VS計)和25d,并在整個試驗中保持不變。試驗共分3個階段(表1),第1階段(第1~25天),R-Fe和R-C僅添加牛糞。第2階段(第26-51天),R-Fe每天額外加入1.5gINP(Fe304,粒徑50-100nm,CAS:1317-61-9),R-C無任何外源添加劑。在第3階段(第52-115天),R-Fe停止添加INP,R-C無任何外源添加劑。CSTR運行模式為半連續進料,即每日進出料一次,先出料后進料。反應溫度為37℃,轉速設定為120r·min-1。
1.2測定指標和分析方法
1.2.1氣體、液體化學組成分析
總固體含量(Total solid,TS)、揮發性固體含量(Volatile solid,VS)、纖維素和半纖維素測定方法參考Li等。沼氣體積由氣體流量計(Ritter,德國)測定,沼氣中CH4和C02含量由微型氣相色譜儀(C2V-200Micro GC,Thermo Scientific,美國)測定。
液體樣品的pH值由pH計測定。總揮發性脂肪酸(Total
volatile fattv
acids,TVFAs)和總堿度(TotalAlkalinity,TA)由自動滴定儀(AT1000,Hach,德國)測定,總化學需氧量(Total chemical oxygen demand.TCOD)用哈希試劑盒(LCK 014,Hach,德國)測定。
1.2.2 16S rRNA分析
在第2階段末期(第51天,標記為R-Fe1和R-C1)和第3階段中期(第75天,標記為R-Fe2和R-C2),取樣(每個處理取3個樣品)進行16S rRNA分析。具體方法:使用FastDNA@Spin Kit提取樣品中總DNA。提取的DNA作為后續PCR反應的模板。使用引物擴增16S-rRNA基因的V3-V4區。其中,細菌用引物338F(5’-ACT CCT ACG GGA GGC AGC AG-3’)和806R(5’-GGA CTA CHV GGG TWT CTA AT-3’)擴增,古菌用引物524F(5’-TGY CAG CCG CCGCGG TAA-3’)和958R(5’-YCC GGC GTT GAV TCCAAT T-3’)擴增。
1.2.3宏基因組分析
為了進一步闡明INP對牛糞厭氧發酵的長期影響,在第3階段結束時(第115天)取樣(每個處理取3個樣品)進行宏基因組分析。使用fastp( https://github.com/OpenGene/fastp)去除適配序列(3’端和5’端)、低質量堿基(lt;20個)、短長度(lt;50 bp)和有N個堿基的讀長,以獲得純凈讀長。然后使用MEGAHIT(https://github.com/voutcn/megahit)對讀長進行組裝(gt;300 bp),用于進一步的基因預測和注釋。使用BLASTP(BLAST
Version
2.2.28+, http: //blast.ncbi.nlm.nih.gov/Blast.cgi)分別對EGGNOG數據庫(http://egg-nog.embl.de/)和KEGG數據庫(http://www.genome.jp/kegg/)進行COG(蛋白質同源群)和KEGG注釋,e值截斷值為e-。
1.2.4數據處理
使用Origin(9.4版)對數據進行t檢驗,以0.05做為顯著性的閾值。
2結果與討論
2.1反應器運行情況
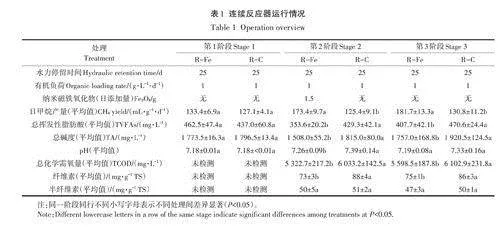
表1總結了反應器的運行情況。在第1階段,R-Fe和R-C的日甲烷產量(以VS計,下同)無顯著差異(Pgt;0.05)。R-Fe為(133.4+6.9) mL·g-1·d-1;R-C為(127.1+4.1)mL·g-1·d-1。這表明在沒有外加INP的情況下兩個反應器具有相似的運行情況。在第2階段,R-Fe中添加INP后,日甲烷產量迅速提高到173.4mL·g-1·d-1,與對照相比提高了38.3%(表1)。類似地,含鐵化合物對牛糞厭氧發酵產甲烷的促進作用在前人的批式實驗中已有報道,如Abdelsalam等發現添加10mg·L-1 FeC13可提升牛糞累積產氣量21.2%;Yun等發現添加鐵鹽[Fe2(S04)3、Fe (N03)3、FeCl3和FeC12]可以縮短反應啟動時間(15-18d),累積甲烷產量提升2.7%-6.4%。此外,R-Fe出料殘渣中纖維素含量顯著低于R-C(Plt;0.05),表明INP添加有利于纖維素水解,這與Dehhaghi等的發現一致。此外,R-Fe中的TVFAs濃度顯著低于R-C(Plt;0.05),表明更多的酸轉化為甲烷,這也與Preeti等、劉莉莉等的發現一致。上述結果表明INP添加有利于水解菌、酸氧化菌和產甲烷菌的富集。然而,添加INP導致TA急劇下降,這是由于發酵料液中C02(以C03-和HC03形式存在)與INP發生礦物碳酸化形成FeC03或Fe2(C03)3所致,而C02的減少會導致堿度下降。第3階段,當R-Fe停止添加INP后,其對產甲烷過程的促進作用仍然持續存在,具體表現在R-Fe中日甲烷產量仍顯著高于R-C(Plt;0.05)(表1)。一個可能的原因是,INP被微生物(如產甲烷菌)吸收并構建甲烷合成酶的相關輔酶因子,并隨著老化微生物裂解釋放到環境中,再被新生微生物利用,如此反復,從而能持續強化產甲烷過程。
2.2 16S rRNA分析
2.2.1 Alpha多樣性
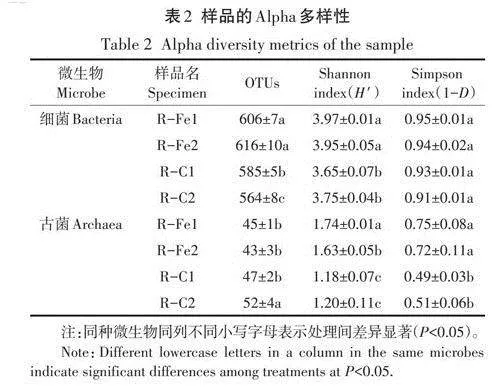
表2通過OTUs、Shannon指數和Simpson指數反映了不同樣本的Alpha多樣性。前人研究表明,優異的厭氧消化性能通常與微生物Alpha多樣性相關,具體表現為更高的Shannon或Simpson指數。本試驗中,R-Fe與R-C相比具有顯著性更高的Alpha多樣性指數(Plt;0.05),尤其體現在產甲烷古菌中(表2)。這也解釋了R-Fe中更優異的產甲烷性能。
2.2.2細菌
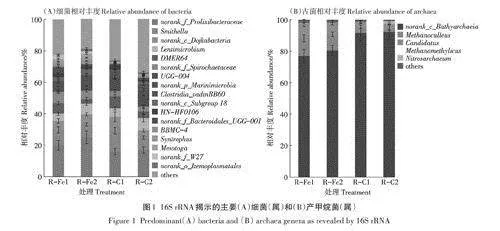
從屬的層面來看,添加INP顯著提高了Syntrophu.s,Me.s otoga和noranh_f_W27的相對豐度(Plt;0.05,圖1)。其中,nora nh_f_Prolixibacteraceae[R-Fel:(19.7±2.8)%,R-Fe2:( 25.3±4.4)%;R-C1:(16.4±2.1)%,R-C2:(17.2±1.9)%]具備降解纖維素的能力,這與R-Fe出料中纖維素濃度顯著低于R-C一致(表1)。[R-Fe 1:(7.4±1.1)%,R- Fe2: (5.3±1.4)%; R-C1:(3.2+ 1.0)%, R-C2:(3.1±1.2)%]和noranh_f_W27[R-Fel:(2.7±0.8)%,R-Fe2:(1.9±0.7)%;R-CI:低于檢測限,R-C2:低于檢測限]均參與酸氧化途徑,并與嗜氫產甲烷菌互利共生。Hassaneen等發現牛糞厭氧發酵中添加鐵鋅氧化物有利于和嗜氫產甲烷菌的富集,并最終提高了甲烷產量;[R-Fel:(5.5±1.3)%,R-Fe2:(7.4±1.1)%;R—Cl:(2.6±0)%,R—C2:(4.1±0.7)%]參與蛋白質氧化產乙酸、二氧化碳和氫氣,并可以分泌酶參與氫氣所攜帶的電子后續利用;Syntro phu.s[R-Fel:(2.0+0.1)%, R-Fe2:(2.0±0.2)%:R-C1:(1.1±0.4)%,R-C2:低于檢測限]是一種典型產氫菌,能參與氫氣介導的種間電子傳遞;Me.sotoga[R-Fe1:(2.3±0.6)%,R-Fe2:(3.1±0.4)%;R-C1:低于檢測限,R-C2:低于檢測限]可通過與嗜氫產甲烷菌的協同作用進行高效的糖氧化(如纖維生物糖和木糖)。此外Mesotoga還能參與乙酸氧化(Syntrophic acetate oxi-dation.SAO)途徑。綜上,添加INP顯著提高了纖維素降解菌和酸氧化菌的相對豐度,這與表1中R-Fe中較低的纖維素濃度和TVFAs濃度一致。
2.2.3產甲烷菌
產甲烷階段是影響厭氧發酵效率的關鍵階段,其中產甲烷菌是關鍵菌群。鐵是構成產甲烷菌細胞組成的重要元素,且在所有的微量元素中,產甲烷菌對鐵的需求量遠高于其他元素。
與細菌的多樣性不同,產甲烷菌主要由四個屬組成。其中BathVarchaeia在所有樣本中占絕對優勢。基于作者之前的研究,Bathyarchaeia是一種具有多種功能的產甲烷菌,且主要分布于以牛糞為主的厭氧發酵體系中。在本研究中,添加INP會顯著降低BathVar-chaeia[R-Fel:(77.3+4.7)%,R-Fe2:( 80.1+3.4)%;R-Cl:(92.0±5.2)%,R-C2:(92.4±4.4)%]的相對豐度,但有利于Methanoculleu.s的富集[R-Fel: (11.3+2.1)%,R-Fe2:( 10.7+1.4)%: R-CI:(4.9±1.1)%,R-C2:(1.5+0.1)%,Plt;0.05]。Methanoculleus作為典型的嗜氫產甲烷菌,可與乙酸氧化菌通過SAO途徑協同產甲烷。有研究表明,在以纖維素或蛋白質為底物的厭氧發酵中,Methanoculleus是產甲烷的主要貢獻者。此外,在作者之前的研究中發現Methanoculleus富集有助于提升牛糞厭氧發酵產氣性能。類似地,高銘雪發現在牛糞批式厭氧發酵中添加INP后,Methanoculleus的相對豐度從對照的2.93%提升到4.58%,相應的甲烷產量提升了33.6%;Preeti等也發現添加鐵化合物將產甲烷菌濃度從4.6x108個·mL-1提升到1.5X10個·mL-1,甲烷產量提升40%。綜合乙酸氧化菌如Mesotoga的富集,本研究揭示了Mesotoga和Methanoculleus之間協同共生的互營關系,這可能是INP提升甲烷產量的主要原因。
2.3宏基因組分析
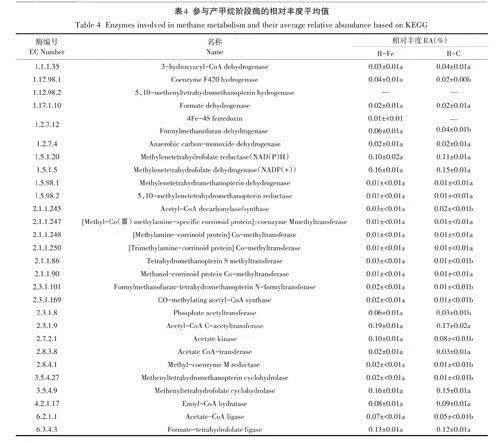
為了進一步闡明INP對牛糞厭氧發酵的長期影響,在第3階段結束時進行了宏基因組分析。基于分析結果預測了參與牛糞厭氧發酵不同代謝途徑的關鍵酶,如表3、表4所示。R-Fe中共含有19445492+674 628個純凈讀長,R-C中共有19286875+1145429個純凈讀長。牛糞富含較難降解的木質纖維素,其中可生物降解的成分主要是纖維素和半纖維素。針對纖維素水解,首先微生物分泌脫支鏈酶,如乙酰木聚糖酯酶(EC 3.1.1.72)、阿拉伯糖苷酶(EC 3.2.1.185/3.2.1.55)和羧酯酶(EC 3.1.1.1),用于去除木聚糖骨架的側鏈單元,使水解酶更容易接觸纖維素和半纖維素單元。這些酶的活性在R-Fe和R-C中無顯著差異(Pgt;0.05)。要徹底降解纖維素需要來自細胞酶相關酶(纖維素酶、外切纖維酶和B -葡萄糖苷酶)的協同作用,尤其是纖維素酶(EC3.2.1.4)。宏基因組結果表明R-Fe的纖維素酶活性顯著高于R-C(Plt;0.05),表明添加INP強化纖維素降解,這與R-Fe中出料殘渣中更低的纖維素濃度以及前人研究一致(表1、表3)。此外,添加INP還顯著強化了葡萄糖苷酶(EC3.2.1.21)的活性(Plt;0.05),這表明添加INP有利于纖維素水解產物(如纖維二糖)的降解,從而加速水解產物的轉化。針對半纖維素水解,首先木聚糖酶(EC3.2.1.8)隨機切斷B-1,4-木聚糖骨架結構中的B-1,4-糖苷鍵,生成木糖和木寡糖。然后,乙酰木聚糖酯酶(EC 3.1.1.72)、a-D-葡萄糖醛酸酶(EC 3.2.1.1)、木聚糖1,4-B-木糖苷酶(EC 3.2.1.37)和阿拉伯糖苷酶(EC 3.2.1.55)協同切斷木糖單體和木聚糖側鏈之間的連接,方便微生物進一步利用。INP添加未顯著提升這些酶的活性(Pgt;0.05).這與出料中接近的半纖維素濃度相一致(表1)。這可能是因為半纖維素的高支鏈結構、低結晶度和異質性本身使其很容易被水解微生物降解。
宏基因組進一步揭示了將水解產物轉化為乙酰輔酶A(乙酸前體物質)的相關酶。從纖維素/半纖維素降解產生的兩個中間產物:B-D-果糖6-磷酸和a-D-葡萄糖6-磷酸主要通過一系列酶轉化為丙酮。生成的丙酮通過丙酮一二氫化輔酶還原酶(EC 1.2.7.1)進一步轉化為乙酰輔酶A。R-Fe中與纖維素水解中間產物利用的相關酶活性顯著高于R-C(Plt;0.05),但與半纖維素水解中間產物利用的相關酶活性無明顯差異(Pgt;0.05)。該結果進一步表明INP添加主要促進纖維素水解及中間代謝產物利用,而對于半纖維素水解和后續利用的提升效果有限。
本質上,厭氧發酵是碳的一種生物固定過程,并以甲烷為最終產物。Fe通過作為多種酶(甲酰一甲呋喃脫氫酶、氫化酶、一氧化碳脫氫酶)的輔因子參與甲烷生成。宏基因組數據表明,嗜乙酸產甲烷途徑和嗜氫產甲烷途徑相關酶均參與了甲烷合成(表4)。經過水解酸化產生的乙酰輔酶A最終通過一系列代謝途徑磷酸乙酰轉移酶(EC 2.3.1.8)、乙酸激酶(EC2.7.2.1).乙酸-CoA連接酶(EC 6.2.1.1)、CO-甲基化乙酰-CoA合成酶(EC 2.3.1.169)、乙酰輔酶脫碳酶/合成酶(EC 2.1.1.245)、四氫甲氧基蝶呤S甲基轉移酶(EC 2.1.1.86)和甲基輔酶M還原酶(EC 2.8.4.1)]最終轉化為甲烷。本研究中,R-Fe的磷酸乙酰轉移酶(EC 2.3.1.8)、乙酸激酶(EC 2.7.2.1)和乙酸-CoA合成酶(EC 6.2.1.1)活性顯著高于R-C(Plt;0.05),表明INP促進了乙酰輔酶A和乙酸之間的相互轉化,從而強化了嗜乙酸產甲烷途徑(表4)。對于嗜氫產甲烷途徑,其主要依賴WL途徑,WL途徑在產甲烷古菌的能量產生和碳固定中起重要作用。嗜氫產甲烷菌(如Methanoculleus)通過WL途徑的甲基支鏈Methyl-Branch Pathway:包含鐵元素的甲酰基甲呋喃脫氫酶(EC 1.2.7.12)、甲酰甲呋喃一四氫氫蝶呤N-甲酰轉移酶(EC 2.3.1.101)、四氫甲氧基苯甲蝶呤環氫酶(EC3.5.4.27)、亞甲基四氫蝶呤脫氫酶(EC 1.5.98.1)和5,10-亞甲基四氫蝶呤還原酶(EC 1.5.98.2)]逐步將二氧化碳還原為Methyl-H4MPT。形成的Methyl-H4MPT的甲基基團通過四氫甲氧基蝶呤S甲基轉移酶(MTR: EC 2.1.1.86)催化到輔酶M,生成CH3-S-CoM,并由甲基輔酶M還原酶(MCR: EC 2.8.4.1)將其還原為甲烷和異二硫酸(CoM-S-S-CoB)。R-Fe中參與嗜氫產甲烷途徑相關酶,如嗜氫產甲烷途徑核心酶一輔酶F420(EC 1.12.98.1),以及甲酰基甲呋喃脫氫酶(EC 1.2.7.12)、甲酰甲呋喃一四氫氫蝶呤N-甲酰轉移酶(EC 2.3.1.101)和四氫甲氧基苯甲蝶呤環氫酶(EC 3.5.4.27)的活性均顯著高于R-C,表明INP添加也促進了嗜氫產甲烷途徑(Plt;0.05,表4)。該結果與16S rRNA中嗜氫產甲烷菌Methanoculleu.s的富集相一致。作為甲烷合成的最后一步,R-Fe中MCR(EC2.8.4.1)活性顯著高于R-C(Plt;0.05,表4),表明添加INP強化了甲烷合成,這與R-Fe中更高的甲烷產量相一致(表1)。
3結論
(1)牛糞連續厭氧發酵中添加納米鐵氧化物(INP)可顯著提升日產甲烷量(Plt;0.05),與對照相比提升38.3%。在停止添加INP后,反應器的日產甲烷量依然顯著高于對照,并持續了至少2個水力停留時間。
(2)16S rRNA分析表明,添加INP富集了纖維素水解菌,以及協同乙酸氧化互營菌:乙酸氧化菌Me.sotoga和嗜氫產甲烷菌Methanoculleu.s,有利于揮發性脂肪酸的代謝,從而促進產甲烷過程。
(3)宏基因組學分析表明,添加INP顯著促進了水解階段、嗜乙酸產甲烷和嗜氫產甲烷階段相關酶活性,并顯著強化了甲烷合成關鍵酶一甲基輔酶M還原酶(EC2.8.4.1)的活性(Plt;0.05)。
