柚皮苷可通過多種途徑修復豬胸膜肺炎放線桿菌誘導的小鼠肺上皮屏障損傷
2024-12-18 00:00:00黃麒霖劉晨景曉涵黃麗娜潘香逸石雄偉仇正英辛蕊華
畜牧獸醫學報 2024年11期


摘 要: 旨在研究柚皮苷(naringin,NAR)對豬胸膜肺炎放線桿菌(Actinobacillus pleuropneumoniae,APP)感染引起小鼠肺上皮屏障損傷的防治效果。將40只6~8周齡的雄性昆明小鼠隨機分為5組,分別為對照組、模型組、NAR低劑量組、NAR中劑量組以及NAR高劑量組。連續灌胃給藥8 d,空白組和模型組給予生理鹽水。在第7天,小鼠以氣管插管灌注的方式感染APP(1×108 CFU·mL-1,100 μL·只-1),空白組灌注等體積無菌生理鹽水,各組小鼠于造模后48 h處死,采樣并評估NAR的防治作用。通過X射線圖像表明,NAR可降低小鼠肺泡灌洗液中APP載菌量,并且能夠緩解肺部病理損傷;此外,NAR干預后,可逆轉由于APP誘導的小鼠肺組織中ZO-1、Occludin-1和Claudin-1緊密連接蛋白的表達降低,從而促進肺上皮屏障修復。為進一步探討NAR促進肺上皮屏障修復的機制,研究發現NAR不僅減少小鼠肺組織中促炎因子IL-6和TNF-α的分泌,還增加了抑炎因子IL-10表達,同時能夠增加抗氧化酶SOD2和Hmox1表達,表明NAR能夠增強機體抗炎及抗氧化能力;此外,NAR可抑制TGF-β1/Smad2/3通路,減少由APP誘導的肺纖維組織增生;通過分析凋亡相關關鍵蛋白和Tunel熒光標記發現,NAR可呈劑量依賴性的減少細胞凋亡。綜上所述,NAR可通過抗炎、抗氧化、抗纖維化和抗凋亡等途徑,修復小鼠肺上皮屏障損傷,從而有效逆轉APP誘導的肺部炎癥損傷、發揮防治APP的功效。
關鍵詞: 天然藥物;柚皮苷;豬胸膜肺炎放線桿菌;豬;肺上皮屏障損傷
中圖分類號: S853.74
文獻標志碼:A
文章編號:0366-6964(2024)11-5299-11
收稿日期:2023-12-26
基金項目:“十四五”國家重點研發項目(2022YFD1801103);國家自然科學基金(32002328);甘肅省重點研發計劃項目(22JR5RA038;21YF5FA166;23YFNA0010;23YFFA0014);蘭州市人才創新創業項目(2021-1-159;2022-RC-21)
作者簡介:黃麒霖(1998-),女,重慶涪陵人,碩士生,主要從事中藥天然免疫研究,E-mail:kylin70521834@163.com
*通信作者:辛蕊華,主要從事中藥天然免疫與中獸藥產品研發,E-mail:xinruihua@caas.cn;仇正英,主要從事獸醫病理學研究,E-mail:qiumoying@163.com
Naringin Repairs Actinobacillus pleuropneumoniae-induced Lung Epithelial Barrier
Damage in Mice via Multiple Pathways
HUANG" Qilin1, LIU" Chen1, JING" Xiaohan1, HUANG" Lina2, PAN" Xiangyi1, SHI" Xiongwei1, QIU" Zhengying1*, XIN" Ruihua1*
(1.Key Laboratory of Veterinary Drug Creation, Ministry of Agriculture and
Rural Affairs, and Gansu Engineering and Technology Research Centre of Chinese Veterinary
Drugs, Lanzhou Institute of Animal Husbandry and Veterinary Medicine, Chinese Academy of
Agricultural Sciences," Lanzhou 730050," China;
2.State Key Laboratory of Applied Organic
Chemistry, College of Pharmacy, Lanzhou University, Lanzhou 730013, "China)
Abstract: The effect of naringin (NAR) on the prevention and treatment of lung epithelial barrier damage induced by Actinobacillus pleuropneumoniae (APP) infection in mice was investigated. Forty male Kunming mice between 6 and 8 weeks of age were randomly divided into five groups, namely, the control group, the model group, the NAR low-dose group, the NAR medium-dose group and the NAR high-dose group. The drug was administered by gavage continuously for 8 d. The blank and model groups were given saline. On d 7, mice were infected with APP (1×108 CFU·mL-1, 100 μL per mouse) by tracheal intubation, and the blank group was infused with an equal volume of sterile saline, and the mice in each group were executed at 48 h after modelling, sampled and evaluated for the preventive effect of NAR. X-ray images showed that NAR reduced the APP load in the alveolar lavage fluid of mice and alleviated the pathological lung injury; moreover, NAR intervention reversed the APP-induced decrease in the expression of ZO-1, Occludin-1, and Claudin-1 tight junction proteins in the lung tissues of mice, thus promoting the repair of the lung epithelial barrier. To further explore the mechanism of NAR promoting lung epithelial barrier repair, it was found that NAR not only reduced the secretion of pro-inflammatory factors IL-6 and TNF-α, but also increased the expression of anti-inflammatory factor IL-10, and increased the expression of antioxidant enzymes SOD2 and Hmox1 in mouse lung tissues, suggesting that NAR enhances the anti-inflammatory and antioxidant capacity of the organism. In addition, NAR can inhibit the TGF-β1/Smad2/3 pathway, reduce APP-induced proliferation of lung fibrotic tissue; analysis of key proteins related to apoptosis and Tunel fluorescent labelling revealed that NAR could reduce apoptosis in a dose-dependent manner. In conclusion, NAR can repair the lung epithelial barrier damage in mice through anti-inflammatory, antioxidant, anti-fibrotic and anti-apoptotic pathways, thus effectively reversing APP-induced inflammatory lung injury and exerting the efficacy of preventing and controlling APP.
Key words: natural medicine; naringin; Actinobacillus pleuropneumoniae; pig; lung epithelial barrier injury
*Corresponding authors:" XIN Ruihua, E-mail: xinruihua@caas.cn; QIU Zhengying, E-mail: qiumoying@163.com
豬胸膜肺炎放線桿菌(Actinobacillus pleuropneumoniae,APP)是引起豬傳染性胸膜肺炎(porcine pleuropneumonia, PCP)重要的病原體,主要侵襲豬的扁桃體和上呼吸道,并在肺部定植和繁殖[1]。PCP在臨床上急性病例以急性出血性、壞死性肺炎為特征[2],亞急性病變以纖維組織包圍的多灶性肺壞死為特征,慢性病例以肺組織局部壞死和慢性纖維素性胸膜肺炎為特征[3]。近年來,PCP在我國多地頻發,且致死率逐年倍增,對養豬業造成重大經濟損失[4]。目前,疫苗接種和抗生素治療是該病的主要防治手段,但由于APP血清型較多[5],疫苗交叉免疫力弱,加劇了養殖業對抗生素的依賴程度,使得APP對抗生素的耐藥性與日俱增[6-7],導致對于該病的防治愈發困難,而且抗生素殘留也增加了食品安全的風險。因此,研發防治APP的藥物,早已迫在眉睫。
中藥具有多靶點、廣效能、綠色環保和耐藥風險低等優勢[8]。應用中藥化橘紅治療呼吸系統疾病在中國已有數千年的歷史,根據清朝《本經逢原》和《本草綱目拾遺》中記載,化橘紅具有祛風寒、化痰、止咳、潤肺等功效,而NAR是其重要的黃酮類化合物[9]。不僅如此,NAR還廣泛存在于蕓香科水果果皮中[10],不僅擁有傳統藥學溯源,且兼顧藥食同源、資源豐富、價格低廉等優勢。多項研究表明,NAR對呼吸道疾病具有良好的治療作用,其作用機制包括抗炎[11]、抗氧化[12-13]、抗纖維化[14]和抗凋亡[15-16]等;而NAR對APP誘導的肺損傷的防治效果及作用機制尚不明確。
肺上皮屏障作為肺組織的第一道防線,在呼吸系統的防御中發揮重要作用,其主要由微血管內皮細胞、肺泡上皮細胞和細胞間連接蛋白組成[17];其中緊密連接蛋白是維持正常上皮屏障功能的重要結構蛋白,以ZO-1、Occludin-1和Claudin-1為主導[18]。這些緊密連接蛋白使相鄰細胞膜緊靠在一起,形成環繞細胞的物理屏障結構,保護機體免受病原微生物的侵害[19]。當APP入侵機體時,會誘發過度的炎癥和氧化應激反應[20],通過損傷上皮細胞及緊密連接蛋白,削弱氣道上皮屏障完整性,引起肺部功能受損[21];其次,受損的肺上皮屏障易為體外的病原微生物提供可乘之機,進一步加劇了炎癥和氧化損傷,這種持續損傷可誘發肺組織纖維化[22];此外,當肺組織中細胞受到強烈損傷會破壞細胞核中DNA,此時受損的細胞會激活凋亡程序以維持組織的正常功能,但過度的細胞凋亡也會加重組織損傷,使機體穩態失衡[23]。綜上所述,肺上皮屏障受損可能會提高機體對APP的易感性,進而引發更進一步的損傷。因此,在本研究中,通過構建小鼠APP肺損傷模型,探究NAR通過抗炎、抗氧化、抗纖維化和抗凋亡等多重機制發揮修復肺上皮屏障損傷的功效,為NAR防治APP提供試驗依據和數據支撐。
1 材料與方法
1.1 材料
NAR購自上海西格瑪奧德里奇貿易有限公司;Bax、Bcl-2、Caspase 3、Cleaved-Caspase 3、ZO-1、Occludin-1、Claudin-1、TGF-β1、Smad2、Smad3、p-Smad2、p-Smad3抗體購自Affinity 公司;GAPDH購自Cell Signaling Technology公司;超敏ECL化學發光液購自北京普利萊基因技術有限公司;BCA蛋白濃度測定試劑盒購自上海碧云天生物技術有限公司;變性蛋白預制膠(濃度4%~15%)、Tris-glycine-SDS電泳緩沖液(10×)、Tris-glycine-SDS轉膜緩沖液(10×)、TBS-T漂洗緩沖液(10×)、SDS-PAGE蛋白上樣緩沖液(5×)、脫脂奶粉、Prestained Protein Marker(11~190 ku)購自上海雅酶生物科技有限公司;NC膜購自武漢博士德生物工程有限公司;Trizol購自賽默飛世爾科技(中國)有限公司;反轉錄試劑盒、qPCR試劑盒購自湖南艾科瑞生物工程有限公司;IL-6、TNF-α和IL-10 ELISA試劑盒、麥康凱瓊脂培養基、無菌PBS溶液購自北京索萊寶科技有限公司。TUNEL試劑盒(11684817910),瑞士羅氏公司產品。
1.2 細菌、試驗動物和處理方法
本實驗室保藏的APP分離自豬場,已經測序Blast比對鑒定。40只SPF級昆明雄性小鼠,體重18~20 g購自中國農業科學院蘭州獸醫研究所(許可證號:SCXK甘肅2023-016)。動物福利聲明遵守機構倫理和動物護理指南,所有程序均根據中國實驗動物護理和使用指南(方案編號:IACUC-157031)進行。將小鼠隨機分為5組,分別為對照組(Control)、模型組(Model)、NAR低劑量組(NARL,20 mg·kg-1)、NAR中劑量組(NARM,40 mg·kg-1)以及NAR高劑量組(NARH,80 mg·kg-1)。連續灌胃給藥8 d,空白組和模型組給予生理鹽水。在第7 天,小鼠腹腔注射戊巴比妥鈉(50 mg·kg-1)麻醉,用氣管插管術感染APP(每只灌注100 μL,菌液濃度為1×108 CFU·mL-1),空白組灌注等體積無菌生理鹽水,各組小鼠于造模成功48 h后麻醉,X射線檢查(DRGEM X射線成像系統)后,處死,取不同樣本用于后續試驗。
1.3 支氣管肺泡灌洗液(BALF)的制備和分析
處死小鼠后,每組隨機取3只小鼠打開胸腔進行氣管插管,用4℃預冷的PBS進行支氣管肺泡灌洗3次,每次0.5 mL,收集共約1.2 mL灌洗液(回收率80%)。然后,通過平板劃線將BALF接種在含5%胎牛血清和1%NAD的TSB固體培養基中,并放置于37℃ 培養箱中培養24 h。計算BALF中的菌落數,并分離單個菌株用于測序驗證。
1.4 實時熒光定量PCR分析
使用TRIzol試劑提取各組細胞或肺組織總RNA,根據PrimeScript RT反轉錄試劑盒將總RNA反轉錄成cDNA。使用SYBR預混Ex Taq II試劑及應用生物系統實時熒光定量PCR系統,利用2-ΔΔCt方法參考β-actin內參基因,對靶基因mRNA轉錄進行定量分析,靶基因引物序列見表1。
1.5 蛋白質印跡分析
用PMSF裂解液處理肺組織或細胞,4℃離心收集上清液作為蛋白質提取物并根據二喹啉甲酸測定法(BCA)測量蛋白質樣品的濃度。將等量的蛋白質樣本用8%~12% SDS-PAGE分離,然后電轉移到聚偏二氟乙烯膜(Millipore,MA,USA)中。將膜用5%脫脂牛奶室溫密封1.5 h,并分別用Bax(1∶1 000)、Bcl-2(1∶1 000)、Caspase 3(1∶1 000)、Cleaved-Caspase 3(1∶1 000)、ZO-1(1∶1 000)、Occludin-1(1∶1 000)、Claudin-1(1∶1 000)、TGF-β1(1∶1 000)、Smad2(1∶1 000)、Smad3(1∶1 000)、p-Smad2(1∶1 000)、p-Smad3(1∶1 000)抗體孵育,加入HRP偶聯的山羊抗鼠IgG二抗(1∶5 000)或HRP偶聯的山羊抗兔IgG二抗(1∶5 000)后,通過
化學發光檢測試劑盒對膜進行可視化,使用Biosciences Imager進行檢測,GAPDH作為參考蛋白質進行比較。此外,將磷酸化蛋白質針對它們各自的總蛋白質進行對比。條帶的灰度值由Image J軟件分析計算。
1.6 ELISA
收集小鼠肺組織,在預冷PBS中清洗去除血液,將清洗好的組織放入含有1 mL PBS的離心管中并在組織研磨機上研磨15 s,最后以3 500 g離心15 min,取上清備用。之后,用IL-6、TNF-α和IL-10 ELISA試劑盒檢測炎癥反應水平。所有程序均按照試劑盒的說明進行。
1.7 組織病理學檢查
收集小鼠的肺組織,在4%中性甲醛中固定24 h,包埋在石蠟中,然后切成5 μm厚。切片用Masson染色與組織免疫熒光。將切片置于顯微鏡(DM 4000B,德國徠卡)下觀察小鼠肺組織。
1.8 組織免疫熒光染色
切片脫蠟至水后浸入EDTA緩沖液進行抗原修復,之后用3%BSA室溫封閉30 min后,4℃一抗孵育過夜。玻片PBS洗滌后,滴加二抗避光室溫孵育50 min。接著避光DAPI復染細胞核,將玻片清洗甩干后,滴加抗熒光淬滅劑封片。使用IF激光掃描共聚焦顯微鏡(NIKON ECLIPSE C1,日本尼康),觀察488 nm(綠色)和405 nm(藍色)的激發波長下的熒光圖像。
切片脫蠟至水后浸入EDTA緩沖液進行微波抗原修復,待自然冷卻,用3% BSA室溫封閉30 min;使用TUNEL試劑盒(11684817910,瑞士羅氏),制備TUNEL反應混合液,處理組用50 μL TdT+450 μL熒光素標記的dUTP液混勻;而陰性對照組僅加50 μL熒光素標記的dUTP液,陽性對照組先加入100 μL DNase 1,后面步驟同處理組。玻片干后,加50 μL TUNEL反應混合液(陰性對照組僅加50 μL熒光素標記的dUTP液)于標本上,加蓋玻片或封口膜在暗濕盒中37℃反應1 h。玻片PBS洗滌后,可以加1滴PBS在熒光顯微鏡下計數凋亡細胞(激發光波長為450~500 nm,檢測波長為515~565 nm);DAPI復染核,PBS漂洗3次,每次8 min;抗熒光淬滅劑封片。使用IF激光掃描共聚焦顯微鏡(NIKON ECLIPSE C1,日本尼康),觀察488 nm(綠色)和405 nm(藍色)的激發波長下的熒光圖像。
1.9 數據統計與分析
使用Image J對 Western blot圖片進行處理。各數據均以“x±s”表示,數據先行正態分布和方差齊性檢驗,符合正態分布且方差齊選用 One-Way ANOVA法進行各組間比較,不符合正態分布采用非參數檢驗。Plt;0.05為差異有統計學意義。SPSS 27用于數據統計分析,GraphPad Prism 9用于作圖。
2 結 果
2.1 NAR對APP誘導小鼠肺組織損傷具有緩解作用
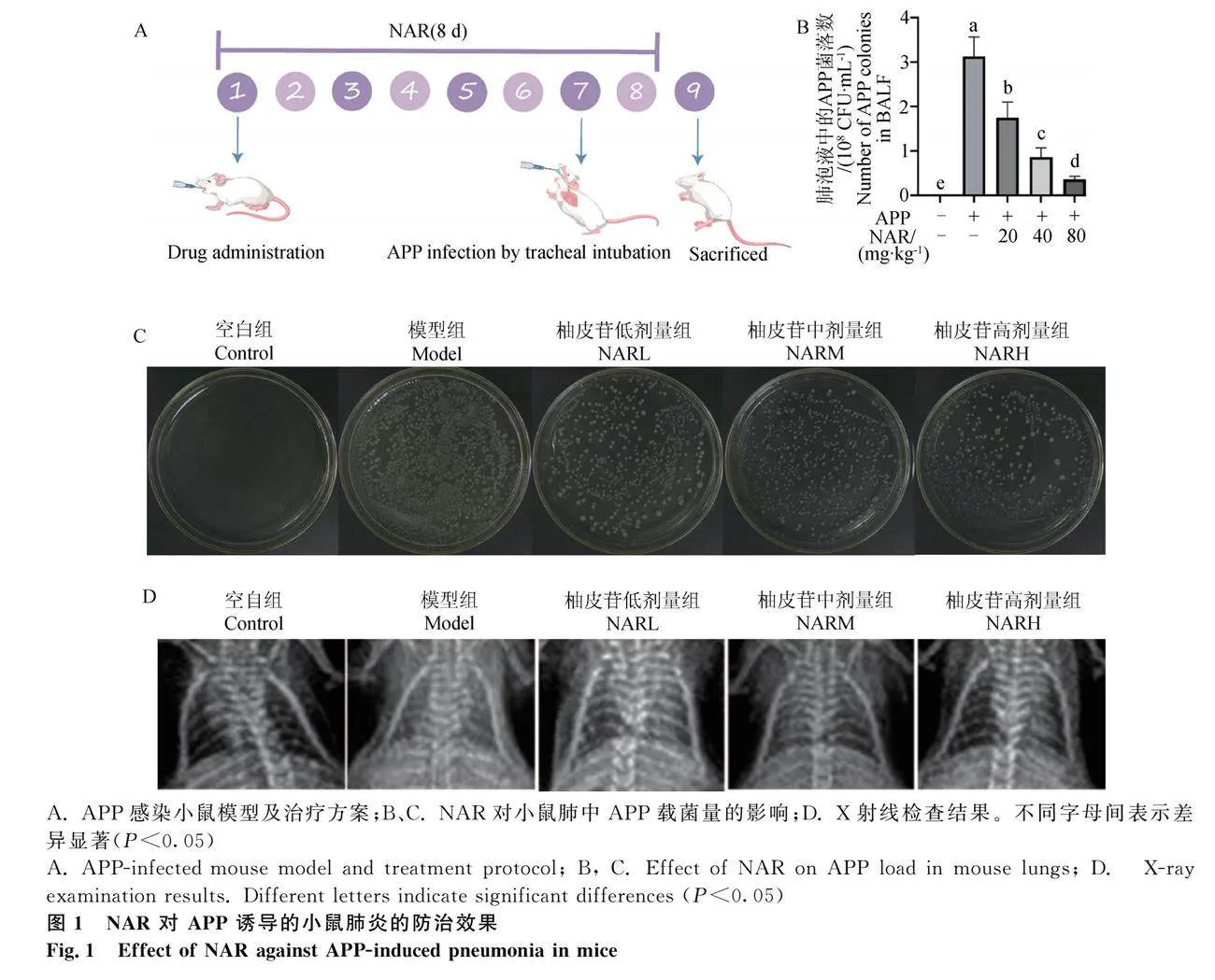
利用X射線檢查、肺泡灌洗液病原菌分析NAR對APP肺炎的預防干預效果。由于肺部病原菌載菌量與肺炎的嚴重程度呈正相關,因此檢測小鼠支氣管肺泡灌洗液(BALF)中APP菌落的數量,結果顯示,模型組小鼠肺組織中載菌量較高,NAR干預后,BALF中APP細菌含量顯著降低(Plt;0.05)(圖1C)。細菌稀釋涂布平板法的結果表明,模型組小鼠BALF中的APP菌落數最多(3.13×108 CFU·mL-1),而低劑量組、中劑量組和高劑量組在NAR作用下的APP菌落數分別為1.74×108、0.85×108和0.36×108 CFU·mL-1(圖1B),表明NAR可減少肺中APP載菌量,且呈劑量依賴性。X射線檢查結果顯示,空白組小鼠雙肺紋理清晰,未見明顯實質性浸潤,兩側肋膈角銳利明晰,未見明顯異常,APP感染組小鼠肺部實變密度增高,雙肺紋理增粗紊亂,邊緣模糊,兩側肋膈角模糊,而NAR干預后各劑量組癥狀均有所緩解(圖1D)。
2.2 NAR可修復由APP誘導的小鼠肺上皮屏障損傷
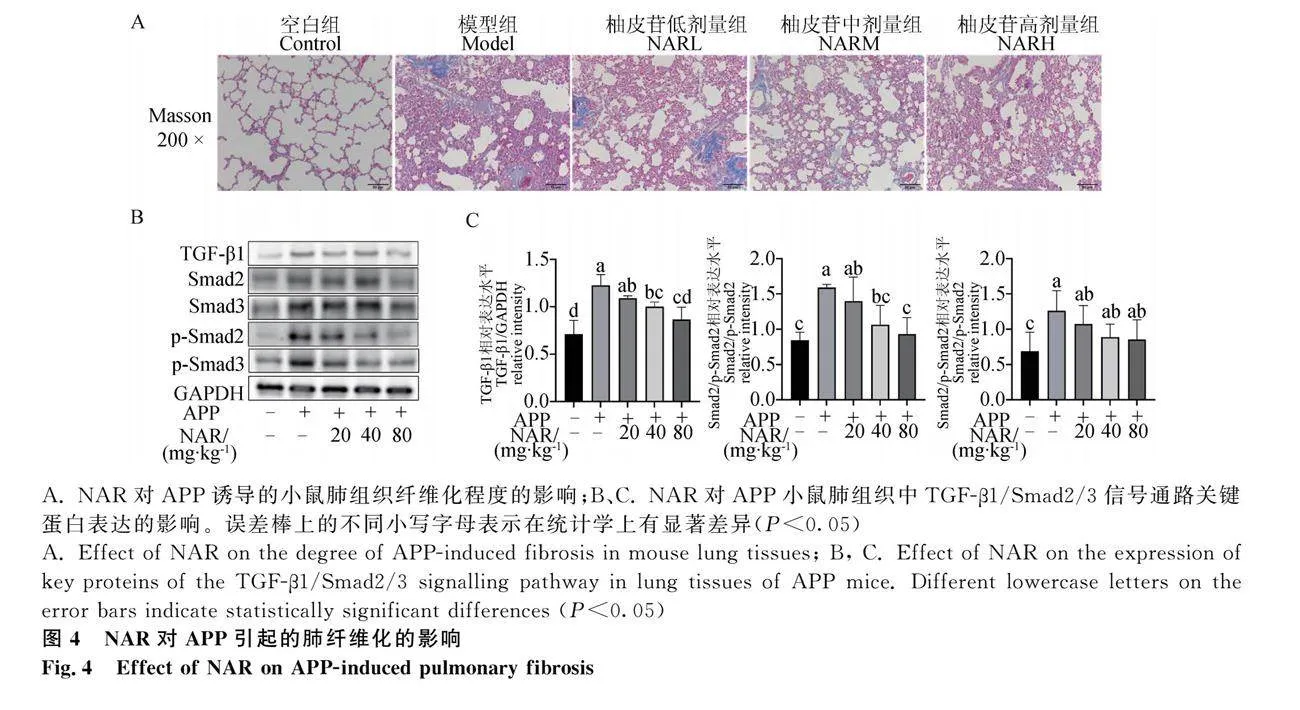
緊密連接蛋白在肺上皮屏障功能中發揮重要的防御作用。為了探索APP對小鼠肺上皮屏障功能的影響,通過對肺組織中緊密連接蛋白ZO-1、Occludin-1和Claudin-1的表達量進行檢測,發現APP感染后,小鼠肺組織中這些蛋白相較于空白組表達量顯著降低(Plt;0.05),APP感染后會破壞肺上皮屏障功能。NAR干擾后,能夠顯著抑制肺組織中由于APP誘導的緊密連接蛋白表達量下降(圖2 A、B);同時,基因檢測分析也證實NAR可劑量依賴性地促進小鼠肺組織中這些緊密連接蛋白的基因表達水平(圖2C)。因此,NAR可通過修復肺上皮屏障功能,從而緩解由APP引起的肺上皮屏障損傷。
2.3 NAR對APP引起的肺部損傷具有抗炎和抗氧化作用
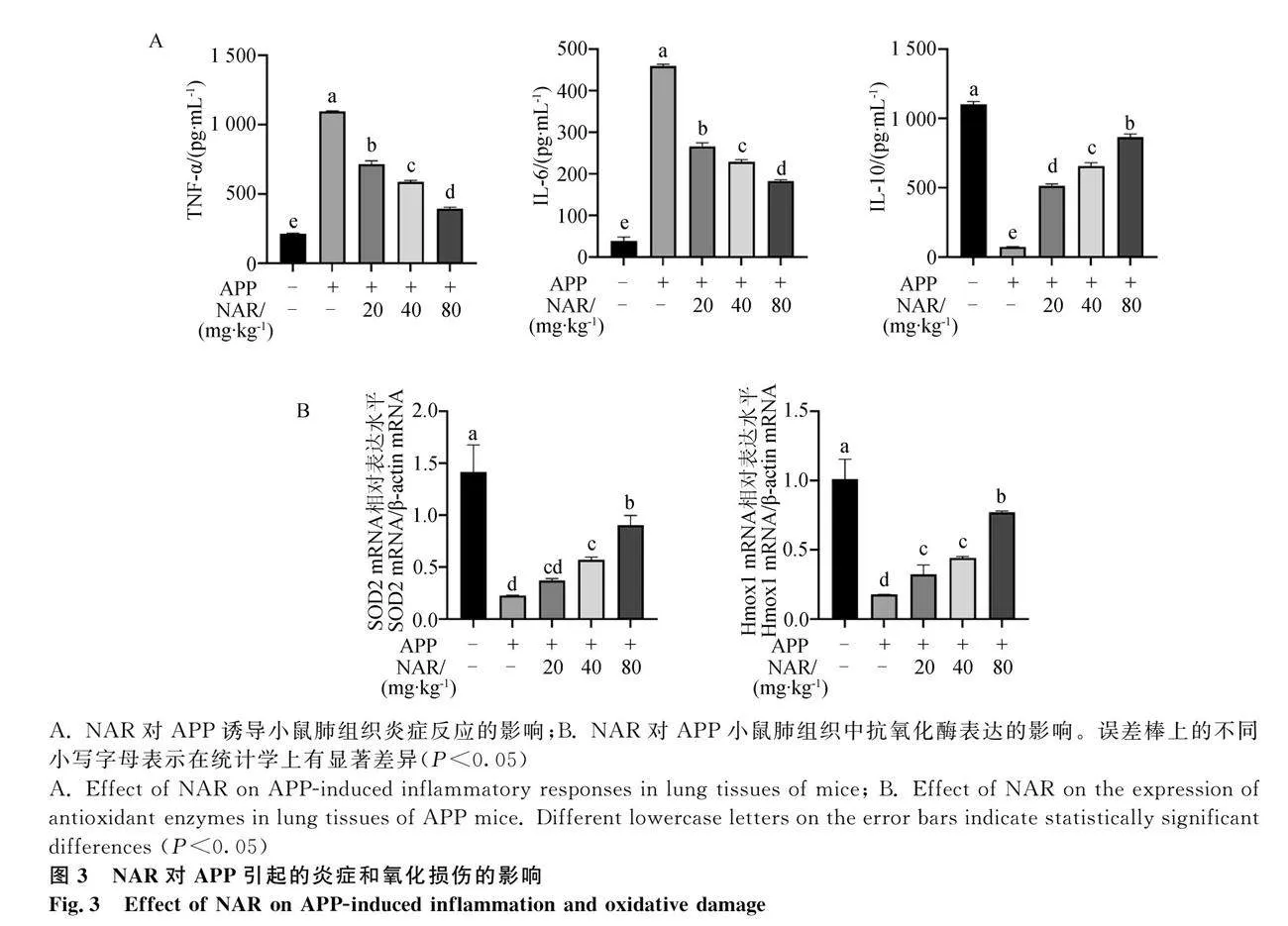
過度的炎癥反應和氧化損傷是造成肺上皮屏障損傷的重要誘因。通過檢測促炎因子TNF-α和IL-6及抑炎因子IL-10的表達水平,發現與空白組相比,APP感染促進小鼠肺組織中促炎因子IL-6、TNF-α的表達,降低抑炎因子IL-10的表達(Plt;0.05),然而NAR干預后可降低APP誘導的小鼠肺組織中TNF-α和IL-6的過度表達,并且提高了肺組織中IL-10的表達水平,緩解APP引起的小鼠肺組織中炎癥因子分泌失衡(圖3A);進一步對抗氧
化酶SOD2和Hmox1進行檢測,發現模型組肺組織中SOD2和Hmox1的表達顯著下降,而NAR給藥后均顯著增加(Plt;0.05)(圖3B)。結果提示,NAR可通過抗炎和抗氧化作用,調控炎癥因子平衡,緩解氧化應激,降低APP引起的肺部損傷。
2.4 NAR可抑制TGF-β1/Smad2/3信號通路緩解肺纖維化
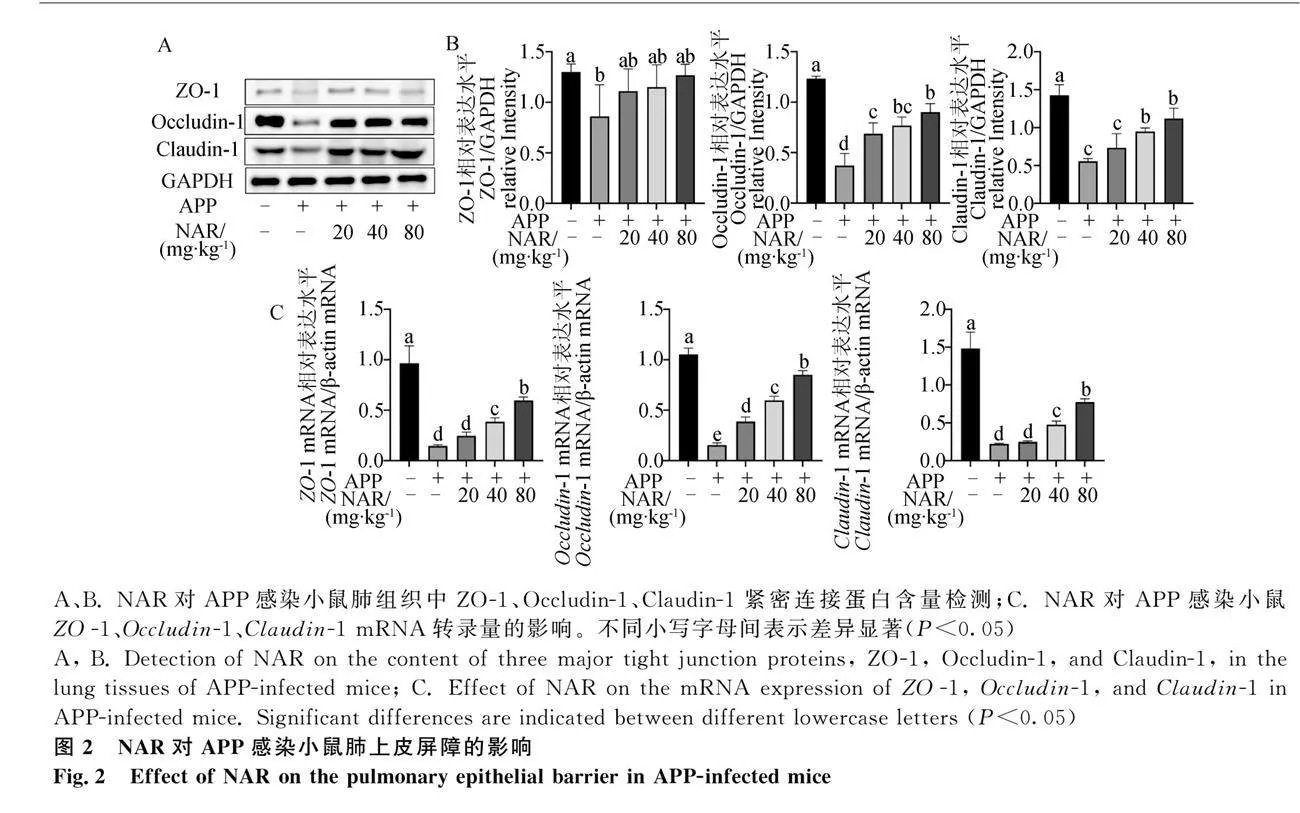
通過Masson染色法觀察各組小鼠肺組織纖維化程度及病理損傷(圖4A)。結果顯示,空白組小鼠肺組織結構完整,肺間質內無明顯膠原纖維沉積。模型組小鼠的黏膜上皮出現損傷,肺泡結構被明顯破壞,肺泡腔內出血,肺泡間隔增厚并充實肺小葉,肺泡隔膜出現膠原纖維沉積,血管及支氣管周圍組織出現疏松、水腫及纖維化形成等病理現象。NAR干預后,各劑量組與模型組相比肺組織損傷存在不同程度的降低,尤其在高劑量組中,氣管結構較完整,氣管黏膜上皮細胞排列較整齊,肺泡壁結構恢復,纖維化程度減輕,僅有少量膠原纖維沉積。為探究NAR抑制肺纖維化的潛在機制,進一步檢測小鼠肺組織中TGF-β1/Smad2/3通路中蛋白表達水平。結果表明,相較于空白組,模型組肺組織中TGF-β1和p-Smad2/3的表達量升高,NAR處理后有所下調(圖4B)。通過Image J對 Western blot圖片進行灰度值計算,結果同樣表明NAR可通過抑制TGF-β1/Smad2/3信號通路的活化從而發揮抗纖維化的作用(圖4C)。
2.5 NAR可抑制細胞凋亡緩解肺損傷
當細胞受到強烈的損傷時,受損的細胞會進行程序性死亡(凋亡),以維持組織正常功能,但過度凋亡也會加重組織損傷。在細胞凋亡信號通路中,活化的Caspase-3和Bax是誘導細胞凋亡的關鍵介質,而Bcl-2是一種抗凋亡蛋白。通過Western blot結果發現,與空白組相比,APP感染會顯著增加小鼠肺組織中裂解的Caspase-3和Bax蛋白水平,降低肺組織中Bcl-2蛋白的表達。NAR干預后,可顯著減弱肺組織中Caspase-3和Bax蛋白的裂解、增加Bcl-2蛋白表達(圖5A、B)。通過熒光Tunel標記發現,與空白組相比模型組小鼠肺組織中凋亡的細胞數量顯著增加,而NAR干預后各組肺組織中細胞凋亡數量顯著降低,并呈劑量依賴性(圖5C)。結果表明,NAR能夠通過抑制肺組織中Caspase-3和Bax活化、降低Bcl-2蛋白表達,從而抑制因APP感染誘導的細胞凋亡。
3 討 論
NAR具有豐富的藥理活性,如抗炎[24]、抗氧化[25]、抗凋亡[26]和抗纖維化[27]等藥理活性,對呼吸系統疾病具有良好的治療效果[28]。本試驗探究了NAR對APP誘導的小鼠肺炎的防治作用。根據預試驗結果,小鼠感染 APP后48 h時小鼠出現明顯的臨床癥狀,因此在后期試驗選擇APP感染后48 h進行相關指標評價。通過X射線檢測發現,APP感染后小鼠肺組織出現雙肺紋理增粗紊亂,肺部實變密度增高,邊緣模糊等癥狀,這與Brauer等[29]的發現一致,而NAR給藥后,其癥狀明顯減輕。APP入侵后在體內大量增殖[30],模型組小鼠肺組織載菌量顯著高于空白組,而NAR給藥后肺組織中APP載菌量顯著降低,且呈劑量依賴性。結果表明,NAR可通過抑制APP增殖,進而緩解肺組織損傷。
APP入侵機體后,破壞宿主上皮屏障的完整性,引起血漿滲透到肺泡間隙,阻礙正常的氣體交換,從而加劇靶器官或組織缺氧的癥狀[31]。緊密連接蛋白在維持上皮屏障完整性方面發揮重要作用,主要包括ZO-1、Occludin-1和Claudin-1在內的緊密連接蛋白[32]。在本研究中,分別從蛋白和基因水平對小鼠肺組織中ZO-1、Occludin-1和Claudin-1的表達量進行檢測。結果表明,NAR能夠緩解APP誘導產生的緊密連接蛋白表達降低,從而恢復肺上皮屏障功能。肺上皮屏障破損后所導致的炎癥與氧化損傷加劇,被認為是APP感染誘導肺損傷的另一個重要因素,可導致肺功能繼續惡化。APP感染機體后,所攜帶的多種毒力因子觸發一系列炎癥級聯反應,激活促炎因子的釋放,如TNF-α、IL-6等,持續的炎癥損傷會進一步激活氧化應激反應,并誘導上皮細胞增生和凋亡[33-35]。本研究發現,NAR不僅能夠降低小鼠肺組織中促炎因子IL-6、TNF-α的表達,并提高抑炎因子IL-10表達,而且還促進肺組織中抗氧化酶SOD2、Hmox1的表達,表明NAR通過抗炎和抗氧化作用緩解APP感染所導致的肺損傷。
前期研究發現,NAR具有緩解肺纖維化的藥理作用[36],本研究通過Masson染色發現,APP感染后小鼠肺組織出現明顯病變,如肺泡結構明顯破壞,肺泡腔內出血,肺泡間隔增厚,肺泡隔膜膠原纖維沉積,血管及支氣管周圍疏松結締組織出現水腫和纖維化形成,且部分病變出現纖維組織包圍的多灶性肺壞死;而NAR干預后,小鼠肺組織纖維化病變程度呈劑量依賴性減輕,表明NAR可一定程度逆轉由APP誘導的纖維化水平。由于TGF-β1/Smad2/3信號通路失調是組織纖維化的重要致病機制[37],主要包括TGF-β1刺激肺上皮細胞發生上皮-間充質轉化(EMT),并通過Smad2/3通路促進肺纖維化[38]。本研究發現,NAR可抑制TGF-β1/Smad2/3信號通路活化,從而減弱APP引起的肺纖維化。此外,本研究發現APP感染后會引起細胞過度凋亡[39],而NAR各劑量組均降低了小鼠肺組織中凋亡蛋白Bax和Caspase-3的活化程度,并增加肺組織中Bcl-2的表達水平。“TUNEL”是末端脫氧核苷酸轉移酶末端標記的縮寫,可通過用修飾的核苷酸對 3′-OH 末端進行酶促標記來檢測細胞凋亡早期 DNA 鏈斷裂的形成,這種方法被廣泛用于檢測細胞凋亡[40]。本試驗通過熒光TUNEL標記試驗,發現與空白組相比,APP感染后小鼠肺組織中細胞凋亡數量明顯增加,而NAR干預后細胞凋亡數量顯著減少。結果表明NAR可通過減少細胞凋亡,從而緩解由APP引起的肺部損傷。
4 結 論
本研究表明,NAR能夠通過促進修復肺上皮屏障、恢復肺上皮屏障功能達到防治APP的目的。其作用機制主要通過減輕肺組織炎癥和氧化應激損傷、抑制TGF-β1/Smad2/3信號通路減緩肺纖維化以及逆轉細胞凋亡誘導的損傷,從而發揮抗APP感染的功效。本研究結果闡述了NAR治療APP的作用機制,為其作為防治APP的新型天然藥物提供理論依據。
參考文獻(References):
[1] pathogen[J]. Livestock, 2020, 25(6):308-314.GALE C, VELAZQUEZ E. Actinobacillus pleuropneumoniae: a review of an economically important pathogen[J]. Livestock, 2020,25(6):308-314.
[2] 劉雨瀟, 胡海兵, 李樹清, 等. 豬胸膜肺炎放線桿菌(APP)1型感染小鼠模型的建立及其病理學觀察[J]. 上海交通大學學報:農業科學版, 2015, 33(5):1-5.
LIU Y X, HU H B, LI S Q, et al." Establish Actinobacillus pleuropneumonise type I infection model in mice and odserve pathologicdl changes[J]. Journal of Shanghai Jiaotong University:Agricultural Science, 2015, 33(5):1-5. (in Chinese)
[3] COHEN L M, GRNTVEDT C A, KLEM T B, et al. A descriptive study of acute outbreaks of respiratory disease in Norwegian fattening pig herds[J]. Acta Vet Scand, 2020, 62(1):35.
[4] LU Y C, LI M C, CHEN Y M, et al. DNA vaccine encoding type IV pilin of Actinobacillus pleuropneumoniae induces strong immune response but confers limited protective efficacy against serotype 2 challenge[J]. Vaccine, 2011, 29(44):7740-7746.
[5] DON V, RAMETTE A, PERRETEN V. Comparative genomics of 26 complete circular genomes of 18 different serotypes of Actinobacillus pleuropneumoniae[J]. Microb Genom, 2022, 8(2):000776.
[6] SITEAVU M I, DRUGEA R I, PITOIU E, et al. Antimicrobial resistance of Actinobacillus pleuropneumoniae, Streptococcus suis, and Pasteurella multocida isolated from Romanian swine farms[J]. Microorganisms, 2023, 11(10):2410.
[7] VILAR A, NOVELL E, ENRIQUE-TARANCON V, et al. Susceptibility trends of swine respiratory pathogens from 2019 to 2022 to antimicrobials commonly used in Spain[J]. Porcine Health Manag, 2023, 9(1):47.
[8] KONG F S, DING Z D, ZHANG K, et al. Optimization of extraction flavonoids from Exocarpium Citri Grandis and evaluation its hypoglycemic and hypolipidemic activities[J]. J Ethnopharmacol, 2020, 262:113178.
[9] LIU G X, LI S Y, ZHANG N M, et al. Sequential grade evaluation method exploration of Exocarpium Citri Grandis (Huajuhong) decoction pieces based on “network prediction→grading quantization→efficacy validation”[J]. J Ethnopharmacol, 2022, 291:115149.
[10] 羅 曦, 包永睿, 李天嬌, 等. 基于“質-量”雙標的枳殼質量分析方法研究[J]. 中草藥, 2023, 54(22):7293-7299.
LUO X, BAO Y R, LI T J, et al." Research on quality analysis method of Aurantii fructus based on“quality-quantity”double standard[J]. Chinese Traditional and Herbal Drugs, 2023, 54(22):7293-7299. (in Chinese)
[11] ZHANG H H, ZHOU X J, ZHONG Y S, et al. Naringin suppressed airway inflammation and ameliorated pulmonary endothelial hyperpermeability by upregulating Aquaporin1 in lipopolysaccharide/cigarette smoke-induced mice[J]. Biomed Pharmacother, 2022, 150:113035.
[12] MOHAMED E E, AHMED O M, ZOHEIR K M A, et al. Naringin-dextrin nanocomposite abates diethylnitrosamine/acetylaminofluorene-induced lung carcinogenesis by modulating oxidative stress, inflammation, apoptosis, and cell proliferation[J]. Cancers (Basel), 2023, 15(20):5102.
[13] 劉 澈, 馬 源, 董旭鵬, 等. 柚皮苷對膿毒癥引起的急性肺損傷小鼠保護作用的研究[J]. 中國臨床藥理學雜志, 2024, 40(5):693-697.
LIU C, MA Y, DONG X P, et al. Protective effect of naringin on sepsis-induced acute lung injury in mice[J]. Chinese Journal of Clinical Pharmacology, 2024, 40(5):693-697. (in Chinese)
[14] WEI Y, SUN L, LIU C, et al. Naringin regulates endoplasmic reticulum stress and mitophagy through the ATF3/PINK1 signaling axis to alleviate pulmonary fibrosis[J]. Naunyn Schmiedebergs Arch Pharmacol, 2023, 396(6):1155-1169.
[15] ZHAO G Y, HUANG Q L, JING X H, et al. Therapeutic effect and safety evaluation of naringin on Klebsiella pneumoniae in mice[J]. Int J Mol Sci, 2023, 24(21):15940.
[16] CHEN Y, NIE Y C, LUO Y L, et al. Protective effects of naringin against paraquat-induced acute lung injury and pulmonary fibrosis in mice[J]. Food Chem Toxicol, 2013, 58:133-140.
[17] JIANG J Y, HUANG K, XU S Q, et al. Targeting NOX4 alleviates sepsis-induced acute lung injury via attenuation of redox-sensitive activation of CaMKII/ERK1/2/MLCK and endothelial cell barrier dysfunction[J]. Redox Biol, 2020, 36:101638.
[18] DALTON C M, SCHLEGEL C, HUNTER C J. Caveolin-1:a review of intracellular functions, tissue-specific roles, and epithelial tight junction regulation[J]. Biology (Basel), 2023, 12(11):1402.
[19] MIN S M, TAO W T, DING D S, et al. Tetramethylpyrazine ameliorates acute lung injury by regulating the Rac1/LIMK1 signaling pathway[J]. Front Pharmacol, 2023, 13:1005014.
[20] ZHANG L, ZHAO F, XU H, et al. HtrA of Actinobacillus pleuropneumoniae is a virulence factor that confers resistance to heat shock and oxidative stress[J]. Gene, 2022, 841:146771.
[21] LI X Y, LIU Z W, GAO T, et al. Tea polyphenols protects tracheal epithelial tight junctions in lung during Actinobacillus pleuropneumoniae Infection via suppressing TLR-4/MAPK/PKC-MLCK signaling[J]. Int J Mol Sci, 2023, 24(14):11842.
[22] AJITO T, HAGA Y, HOMMA S, et al. Immunohistological evaluation on respiratory lesions of pigs intranasally inoculated with Actinobacillus pleuropneumoniae serotype 1[J]. J Vet Med Sci, 1996, 58(4):297-303.
[23] TANG H, WANG R, PANG S Q, et al. Native ApxIIA secreted by Actinobacillus pleuropneumoniae induces apoptosis in porcine alveolar macrophages dependent on concentration and acylation[J]. Vet Microbiol, 2023, 287:109908.
[24] WANG Y Y, WANG X, LI Y X, et al. Xuanfei Baidu Decoction reduces acute lung injury by regulating infiltration of neutrophils and macrophages via PD-1/IL17A pathway[J]. Pharmacol Res, 2022, 176:106083.
[25] LEE D H, WOO J K, HEO W, et al. Citrus junos Tanaka peel extract and its bioactive naringin reduce fine dust-induced respiratory injury markers in BALB/c male mice[J]. Nutrients, 2022, 14(5):1101.
[26] SALAMA A A A, YASSEN N N, MANSOUR H M. Naringin protects mice from D-galactose-induced lung aging and mitochondrial dysfunction:Implication of SIRT1 pathways[J]. Life Sci, 2023, 324:121471.
[27] TURGUT N H, KARA H, ELAGOZ S, et al. The Protective effect of naringin against bleomycin-induced pulmonary fibrosis in Wistar rats[J]. Pulm Med, 2016, 2016:7601393.
[28] HUANG Q L, LI W, JING X H, et al. Naringin’s alleviation of the inflammatory response caused by Actinobacillus pleuropneumoniae by downregulating the NF-κB/NLRP3 signalling pathway[J]. Int J Mol Sci, 2024, 25(2):1027.
[29] BRAUER C, HENNIG-PAUKA I, HOELTIG D, et al. Experimental Actinobacillus pleuropneumoniae challenge in swine:comparison of computed tomographic and radiographic findings during disease[J]. BMC Vet Res, 2012, 8(1):47.
[30] ZHANG Q H, PENG L, HAN W Y, et al. The morphology and metabolic changes of Actinobacillus pleuropneumoniae during its growth as a biofilm[J]. Vet Res, 2023, 54(1):42.
[31] 黃麒霖, 仇正英, 王貴波, 等. 豬傳染性胸膜肺炎發病機制研究進展[J]. 黑龍江畜牧獸醫, 2024(3):31-37.
HUANG Q L, QIU Z Y, WANG G B, et al." Research progress on pathogenesis of porcine pleuropneumonia[J]. Heilongjiang Animal Science and Veterinary Medicine, 2024(3):31-37. (in Chinese)
[32] CAO Q, WEI W B, WANG H, et al. Cleavage of E-cadherin by porcine respiratory bacterial pathogens facilitates airway epithelial barrier disruption and bacterial paracellular transmigration[J]. Virulence, 2021, 12(1):2296-2313.
[33] TANG H, ZHANG Q H, HAN W Y, et al. Identification of FtpA, a Dps-like protein involved in anti-oxidative stress and virulence in Actinobacillus pleuropneumoniae[J]. J Bacteriol, 2022, 204(2):e0032621.
[34] HU Y F, JIANG C S, ZHAO Y Q, et al. TurboID screening of ApxI toxin interactants identifies host proteins involved in Actinobacillus pleuropneumoniae-induced apoptosis of immortalized porcine alveolar macrophages[J]. Vet Res, 2023, 54(1):62.
[35] JING X H, ZHAO G Y, WANG G B, et al. Naringin alleviates pneumonia caused by Klebsiella pneumoniae infection by suppressing NLRP3 inflammasome[J]. Biomed Pharmacother, 2024, 170:116028.
[36] WU Y H, CAI C H, XIANG Y J, et al. Naringin ameliorates monocrotaline-induced pulmonary arterial hypertension through endothelial-to-mesenchymal transition inhibition[J]. Front Pharmacol, 2021, 12:696135.
[37] LV Q, WANG J J, XU C Q, et al. Pirfenidone alleviates pulmonary fibrosis in vitro and in vivo through regulating Wnt/GSK-3β/β-catenin and TGF-β1/Smad2/3 signaling pathways[J]. Mol Med, 2020, 26(1):49.
[38] WANG Z J, LIU M, AI Y, et al. The compound artemisinin-hydroxychloroquine ameliorates bleomycin-induced pulmonary fibrosis in rats by inhibiting TGF-β1/Smad2/3 signaling pathway[J]. Pulm Pharmacol Ther, 2023, 83:102268.
[39] DUQUETTE S C, FISCHER C D, WILLIAMS A C, et al. Immunomodulatory effects of tulathromycin on apoptosis, efferocytosis, and proinflammatory leukotriene B4 production in leukocytes from Actinobacillus pleuropneumoniae-or zymosan-challenged pigs[J]. Am J Vet Res, 2015, 76(6):507-519.
[40] RESNICK-SILVERMAN L. Using TUNEL assay to quantitate p53-induced apoptosis in mouse tissues[J]. Methods Mol Biol, 2021, 2267:181-190.
(編輯 白永平)
