鎘脅迫下甜瓜幼苗葉片抗氧化酶活性及根系活力變化差異研究
2024-12-25 00:00:00孔維萍唐桃霞任凱麗程鴻岳宏忠
甘肅農業科技 2024年12期
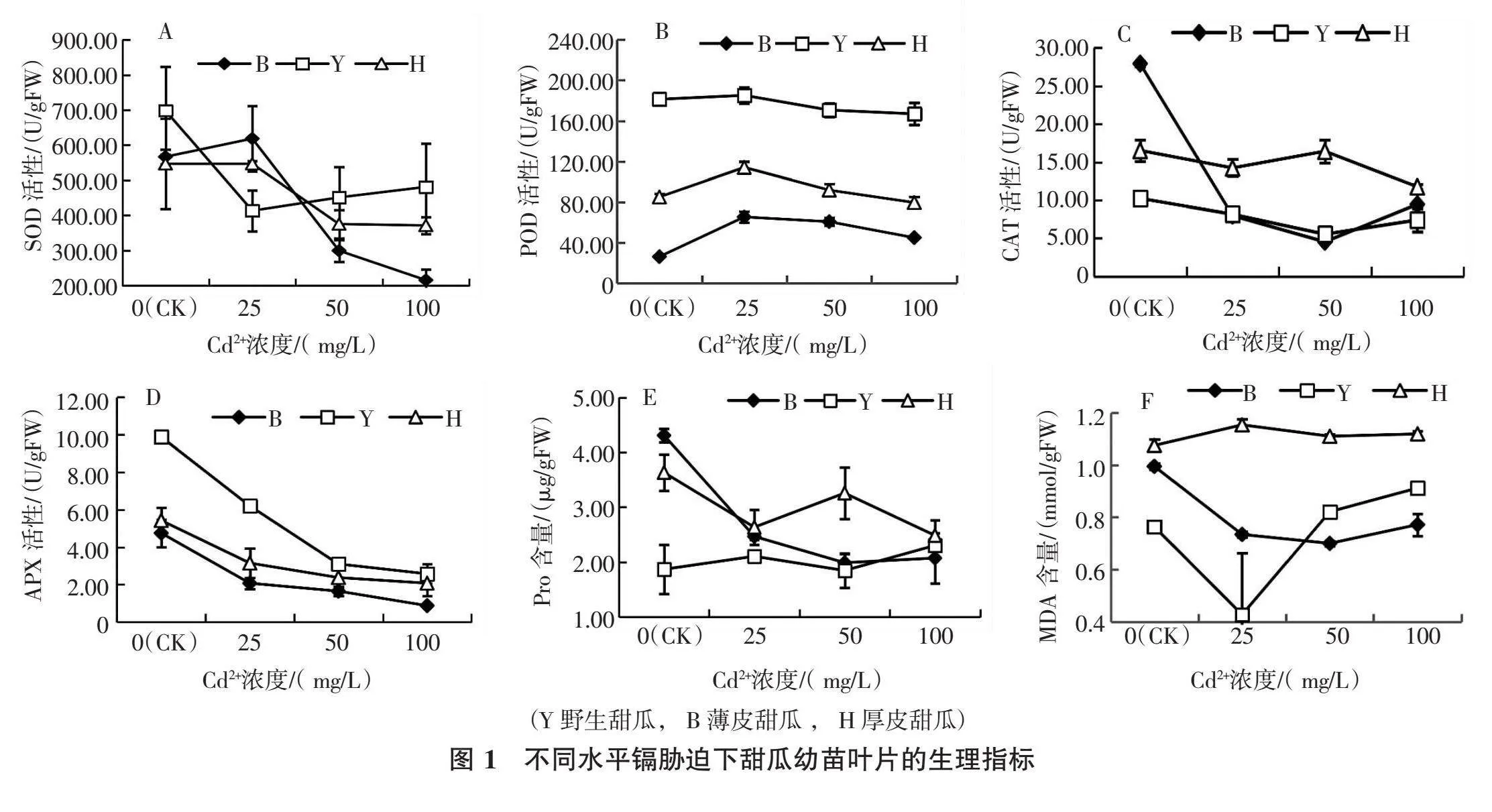


摘要:通過探究鎘脅迫下不同甜瓜幼苗葉片抗氧化酶活力及根系活力變化,為篩選抗鎘甜瓜種質提供參考。以野生甜瓜、薄皮甜瓜及厚皮甜瓜為試材,在苗期用不同濃度鎘溶液培養后,檢測甜瓜幼苗葉片相關酶活及根系活力變化差異。結果表明,鎘脅迫下甜瓜幼苗葉片中SOD、POD、CAT、APX活性均低于對照不添加Cd2+。隨著Cd2+濃度增大,薄皮甜瓜和厚皮甜瓜幼苗葉片的SOD活性大幅下降,而野生甜瓜幼苗葉片的SOD活性略有回升,且在Cd2+中高濃度(50~100 mg/L)時均以野生甜瓜幼苗葉片的SOD活性最高,薄皮甜瓜的幼苗葉片的SOD活性最低。各處理下野生甜瓜幼苗葉片的POD活性均最高,薄皮甜瓜幼苗葉片的POD活性均最低,隨Cd2+脅迫濃度的增大,不同甜瓜材料幼苗葉片的POD活性均下降,但降幅不大。Cd2+脅迫濃度為0 mg/L時,薄皮甜瓜幼苗葉片的CAT活性最高,野生甜瓜幼苗葉片的CAT活性最低;隨Cd2+脅迫濃度的增大,薄皮甜瓜幼苗葉片的CAT活性降幅最大。各處理下野生甜瓜幼苗葉片的APX活性均最高,薄皮甜瓜幼苗葉片的APX活性最低,而野生甜瓜幼苗葉片的APX活性隨Cd2+濃度增大降幅較大。當Cd2+脅迫濃度為0 mg/L時,野生甜瓜幼苗葉片的Pro含量最低,薄皮甜瓜幼苗葉片的Pro含量最高;隨Cd2+脅迫濃度的增大,野生甜瓜幼苗葉片的Pro含量呈變化不明顯,薄皮甜瓜則逐步遞減,厚皮甜瓜呈先降后升又略下降趨勢。各處理下均以厚皮甜瓜幼苗葉片的MDA含量最高,隨Cd2+脅迫濃度的增大,野生甜瓜幼苗葉片的MDA含量呈先降后升趨勢,薄皮甜瓜的MDA含量呈逐步遞減趨勢,厚皮甜瓜的MDA含量先升后降又略上升。在相同Cd2+脅迫濃度處理下,野生甜瓜根系活力最強,薄皮甜瓜根系活力降幅最大。由此可見,鎘脅迫下薄皮甜瓜幼苗葉片的酶活性降幅較大,厚皮甜瓜次之,野生甜瓜幼苗葉片的酶活性降幅相對較小,且各處理下均以野生甜瓜根系活力最強,這說明參試的薄皮甜瓜對鎘脅迫較敏感,野生甜瓜抗鎘性較厚皮甜瓜、薄皮甜瓜強。
關鍵詞:甜瓜;幼苗葉片;鎘脅迫;抗氧化酶;活性;根系活力
中圖分類號:S651 文獻標志碼:A 文章編號:2097-2172(2024)12-1147-06
doi:10.3969/j.issn.2097-2172.2024.12.013
Differences of Leaf Antioxidant Enzymes Activitiesand Root Vigor of
Melon Seedlings under Cadmium Stress
KONG Weipin, TANG Taoxia, REN Kaili, CHANG Hong, YUE Hongzhong
(Institute of Vegetables, Gansu Academy of Agricultural Sciences, Lanzhou Gansu 730070, China)
Abstract: This study explored the changes of antioxidant enzymeactivities and rootvigor in different melon seedling leaves under cadmium (Cd) stress so as to provide references for selecting Cd-resistant melon germplasm. Wild, thin-skinned and thick-skinned melons were used as experimental materials. After cultivation with different concentrations of cadmium solutions during the seedling stage, the variations in enzyme activity and root vigor in240fd3a0b3fb3db236c69d8c9f774ecc97c108da9b68490e82a42bc4e76298a2 the leaves of different melon seedlings were detected. Results showed that under cadmium stress, the activities of SOD, POD, CAT, and APX in melon seedling leaves were all lower than the control without added Cd??. As the Cd?? concentration increased, the SOD activity in thin-skinned and thick-skinned melon seedlings decreased significantly, while the SOD activity in wild melon seedlings slightly increased. At medium to high Cd?? concentrations(50 to 100 mg/L), the SOD activity in wild melon seedling leaves was the highest, and the SOD activity in thin-skinned melon seedlings was the lowest.Under all treatments, the POD activity in wild melon seedling leaves was the highest, and that in thin-skinned melon seedlings was the lowest. With the increasing concentration of Cd2+ stress, the POD activity in the leaves of different melon materials decreased, but the decrease was not significant. When the Cd2+ concentration was 0 mg/L, the CAT activity in thin-skinned melon seedling leaves was the highest, while that in wild melon seedlings was the lowest. As the Cd2+ concentration increased, the CAT activity in thin-skinned melon seedling leaves decreased the most. Under all treatments, the APX activity in wild melon seedling leaves was the highest, and the APX activity in thin-skinned melon seedling leaves was the lowest. The APX activity in wild melon seedling leaves decreased significantly with increasing Cd2+ concentration. When the Cd2+ stress concentration was 0 mg/L, the Pro content in wild melon seedling leaves was the lowest, and that in thin-skinned melon seedling leaves was the highest. As the Cd2+ concentration increased, the Pro content in wild melon seedling leaves showed no significant change, while the Pro content in thin-skinned melon steadily decreased, and in thick-skinned melon, it decreased initially, then slightly increased, and decreased again.Under all treatments, the MDA content in thick-skinned melon seedling leaves was the highest. As the Cd?? stress concentration increased, the MDA content in wild melon seedling leaves showed a decreasing and then increasing trend, while the MDA content in thin-skinned melon steadily decreased, and that in thick-skinned melon increased initially, then decreased, and slightly increased again. At the same Cd concentration, wild melon exhibited the strongest root vigor, while thin-skinned melon showed the largest decrease in root vigor. It can be concluded that under cadmium stress, thin-skinned melon seedlings showed the largest decrease in enzyme activity, followed by thick-skinned melon, while wild melon seedlings exhibited a relatively smaller decrease. Furthermore, wild melon consistently showed the strongest root vigor under all treatments, indicating that thin-skinned melon is more sensitive to cadmium stress, while wild melon has stronger cadmium resistance compared to thick-skinned and thin-skinned melons.
Key words: Melon; Seedling leave; Cadmium stress; Antioxidant enzyme; Vigor; Root activity
鎘(cadmium, Cd)是一種毒性較大的重金屬,人體若攝入過量的鎘,就會有致畸、致癌的風險。隨著工業“三廢”排放、化肥農藥濫用、污水灌溉及城鎮污泥的使用,使得環境鎘污染越來越嚴重,因此植物抗、耐鎘機理方面的研究越來越受到人們的重視[1 - 2 ]。作物中鎘殘留和抗鎘機制方面的研究比較多[3 - 7 ]。鎘脅迫對甜瓜幼苗生長的影響目前也有研究[8 - 9 ],主要涉及光合系統和抗氧化酶活性變化等內容。植物對鎘脅迫應激反應因基因型而異,本試驗選取3份不同類型甜瓜單系材料為試材,開展了不同鎘濃度脅迫下幼苗葉片抗氧化酶活性變化差異研究,旨在為篩選抗耐鎘甜瓜種質資源提供數據支撐,為選育抗鎘甜瓜新品種奠定基礎。
1 材料與方法
1.1 供試材料
供試鎘源為CdCl2·5H2O(分析純試劑),由Sigma公司生產。種子萌發袋(每袋50 mL)購自北京啟維益成科技有限公司。供試材料為厚皮甜瓜白蘭瓜(H)、薄皮甜瓜白脆瓜(B)、野生甜瓜泡狀瓜(Y),均為多年自交選純單系材料,其來源及特性見表1。
1.2 試驗方法
試驗分別設置鎘溶液脅迫濃度為0(CK)、25、50、100 mg/L。將正常露白的甜瓜種子置于種子萌發袋萌發孔處,內盛50 mL 1/8 的霍格蘭氏營養液,營養液每48 h更換1次。約7 d后子葉展開,2葉1心時按試驗設計添加不同濃度CdCl2·5H2O 的1/8霍格蘭氏營養液培養,每天更換新的培養液。處理8 d后取樣檢測生理生化指標進行對比分析。
1.3 指標測定
超氧化物歧化酶(SOD)活性測定采用氮藍四唑(NBT)光還原法,以抑制NBT光化還原的50%為1個酶活力單位[10 - 11 ];過氧化物酶(POD)活性測定采用愈創木酚比色法測定,提取緩沖液代替測定液測定△OD470ck,以OD470每增加1為一個酶活力單位[10 - 11 ];過氧化氫酶(CAT)活性采用過氧化氫分解法測定,提取緩沖液代替測定液測定△ OD240ck,以OD240每分鐘減少1為一個酶活力單位[10 - 11 ];抗壞血酸過氧化物酶(APX)采用Grace SG(1996)的方法[10 - 11 ];可溶性蛋白(Pro)含量測定采用考馬斯亮藍G250法,以牛血清蛋白為標準蛋白[10 - 11 ];丙二醛(MDA)含量測定采用硫代巴比妥酸法[10 - 11 ];根系活力測定采用TTC法[10 - 11 ]。
1.4 數據處理
采用Excel 2016軟件對試驗數據進行整理,運用SPSS 22.0軟件進行數據分析。
2 結果與分析
2.1 超氧化物歧化酶(SOD)活性
植物在長期的進化過程中,體內同時存在2種主要的抗氧化系統,可以及時清除機體內過多的活性氧,以維持自由基代謝的動態平衡[12 - 14 ]。SOD在活性氧清除系統中起主要作用,它催化2O2-2+2H+=H2O2+O2[12 - 14 ]。 Cd2+脅迫濃度為0 mg/L(CK)時,野生甜瓜幼苗葉片的 SOD活性最大,薄皮甜瓜的 SOD活性和厚皮甜瓜相當。Cd2+脅迫濃度為25 mg/L時,薄皮甜瓜幼苗葉片的SOD活性略高于CK,而野生甜瓜幼苗葉片的SOD活性大幅下降,約為CK的57%;厚皮甜瓜幼苗葉片的SOD活性基本不變。但隨著Cd2+脅迫濃度繼續增大,薄皮甜瓜和厚皮甜瓜幼苗葉片的SOD活性大幅下降,而野生甜瓜幼苗葉片的SOD活性略有回升,且以野生甜瓜的SOD活性最高,薄皮甜瓜的SOD活性最低(圖1 A)。在Cd2+脅迫濃度為100 mg/L時,薄皮甜瓜幼苗葉片的SOD活性顯著低于野生甜瓜和厚皮甜瓜。
2.2 過氧化物酶(POD)活性
POD可催化H2O2與酚類的反應形成H2O[12 - 14 ]。各Cd2+脅迫濃度處理下均以野生甜瓜幼苗葉片的POD活性最高,約為薄皮甜瓜的3倍,厚皮甜瓜的2倍。Cd2+脅迫濃度為25 mg/L時,不同甜瓜材料幼苗葉片的POD活性均高于CK。隨Cd2+脅迫濃度的增大,不同甜瓜材料幼苗葉片的POD活性均下降,但降幅不大(圖1 B)。薄皮甜瓜幼苗葉片POD活性各處理間差異均達顯著水平,相同Cd2+脅迫濃度處理下不同甜瓜材料幼苗葉片的POD活性差異也均達顯著水平。
2.3 過氧化氫酶(CAT)活性
CAT是一種含Fe的血蛋白酶類,催化H2O2分解成水和氧,CAT活性的提高能更有效地清除H2O2,與SOD均可對植物生長起保護作用[12 - 14 ]。在CK條件下,薄皮甜瓜幼苗葉片的CAT活性最高,野生甜瓜的最低。Cd2+脅迫濃度為25 mg/L時,不同甜瓜材料幼苗葉片的CAT活性均下降,薄皮甜瓜幼苗葉片的CAT活性下降明顯,約為CK的29%。Cd2+脅迫濃度繼續增加時,不同甜瓜材料幼苗葉片的CAT活性均趨于平穩(圖1 C)。除CK外,厚皮甜瓜幼苗葉片的CAT活性在不同Cd2+脅迫濃度處理下顯著高于薄皮甜瓜和野生甜瓜。
2.4 抗壞血酸過氧化物酶(APX)活性
APX是葉綠體中清除H2O2的主要酶[12 - 14 ]。不同Cd2+脅迫濃度處理下均以野生甜瓜幼苗葉片的APX活性均最高,薄皮甜瓜的最低;隨著Cd2+脅迫濃度的變大,不同甜瓜材料幼苗葉片的APX活性均呈下降趨勢,其中以野生甜瓜的APX活性降幅最大,當Cd2+脅迫濃度為100 mg/L時,幼苗葉片的APX活性約為CK的1/3,而薄皮甜瓜和厚皮甜瓜幼苗葉片的APX活性下降幅度基本相當(圖1 D)。不同甜瓜材料幼苗葉片的APX活性在CK條件下與其余Cd2+脅迫濃度處理間差異均達顯著水平。
2.5 可溶性蛋白(Pro)含量
可溶性蛋白是植物體內重要的滲透調節物質和營養物質,其含量越高說明植物體的代謝活動越強[15 - 17 ]。可溶性蛋白增加對細胞的生命物質及生物膜起到保護作用,可溶性蛋白(Pro)含量的高低常被用作篩選植物抗性指標之一[12 - 14 ]。當Cd2+脅迫濃度為0 mg/L(CK)時,以野生甜瓜幼苗葉片的Pro含量最低,薄皮甜瓜幼苗葉片的Pro含量最高。隨Cd2+脅迫濃度的變大,野生甜瓜幼苗葉片的Pro含量變化趨勢不明顯,薄皮甜瓜的Pro含量呈逐步遞減趨勢,厚皮甜瓜呈先降后升又略下降趨勢,這可能是甜瓜幼苗抵抗鎘損傷能力差異造成的。Cd2+脅迫濃度達到100 mg/L時,薄皮甜瓜、厚皮甜瓜幼苗葉片的Pro含量均較CK明顯下降,而野生甜瓜的Pro含量較CK略有上升。由此可見,高濃度鎘脅迫嚴重抑制了薄皮甜瓜、厚皮甜瓜幼苗葉片的可溶性蛋白(Pro)的合成(圖1 E)。薄皮甜瓜幼苗葉片的Pro含量在CK處理下與其余Cd2+脅迫濃度處理間差異均達顯著水平,厚皮甜瓜和野生甜瓜的不同Cd2+脅迫濃度處理間差異均不顯著。
2.6 丙二醛(MDA)含量
MDA是膜脂過氧化作用的主要產物之一,通常利用它的含量作為脂質過氧化指標,反映細胞膜脂過氧化的程度。低濃度(25 mg/L)Cd2+處理下,以野生甜瓜幼苗葉片MDA含量最低,中高濃度(50~100 mg/L)Cd2+處理下,以薄皮甜瓜幼苗葉片MDA含量最低;厚皮甜瓜MDA含量在各處理條件下均最大,且基本處于恒定的水平。隨Cd2+脅迫濃度的變大,野生甜瓜幼苗葉片的MDA含量呈先降后升趨勢,薄皮甜瓜呈逐步遞減趨勢,厚皮甜瓜呈先升后降又略上升趨勢(圖1 F)。薄皮甜瓜MDA含量CK與其他處理間差異顯著,其余材料各處理間差異不顯著。
2.7 根系活力
根是植物體中最重要的絡合重金屬的部位,也是最易受重金屬毒性影響的部位。在整個培養過程中觀察發現厚皮甜瓜植株生長健壯,根密且粗壯,生物產量最大,充滿了整個培養空間;野生甜瓜整體長勢最弱,根系生長量小(圖2)。
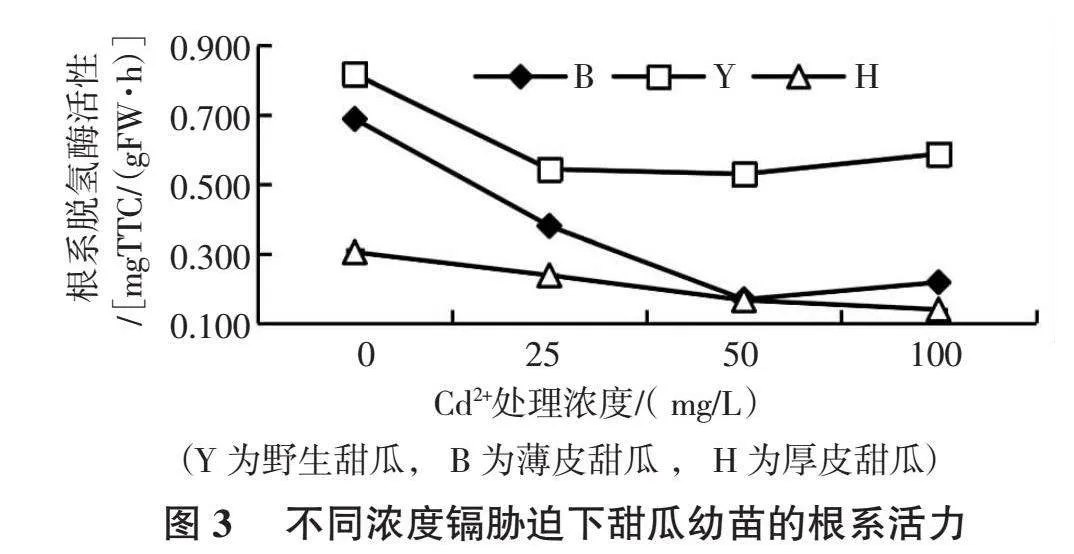
在同等濃度Cd2+處理下,野生甜瓜根系活力最強,約為厚皮甜瓜的3倍,隨 Cd2+濃度加大,野生甜瓜根系活力先大幅下降而后略有上升;薄皮甜瓜根系活力的降幅最大,100 mg/L Cd2+濃度下其活力僅為對照的1/6;厚皮甜瓜根系活力整體呈緩慢遞減趨勢(圖3)。高濃度鎘脅迫下甜瓜幼苗根系變黃、變褐、根尖壞死等脅迫癥狀,厚皮甜瓜表現尤為突出。甜瓜幼苗根系活力同一材料各處理間差異均顯著。
3 結論與討論
植物抗氧化酶主要包括 SOD、POD、APX 和 CAT,能相互協助清除重金屬脅迫下產生的活性氧,減輕細胞結構受損,從而緩解植物受毒害癥狀。本研究中,3種類型甜瓜試材抗氧化酶活性隨著鎘處理濃度的增加整體呈下降趨勢,其中SOD活性和POD活性在低濃度鎘脅迫下稍有提升,這表明甜瓜植株葉片細胞自我保護機制啟動。而后隨鎘脅迫濃度增大持續下降。而APX、CAT活性呈持續下降趨勢,這與張永平等[9 ]的研究結果鎘脅迫下甜瓜幼苗抗氧化酶活性整體先升后降的變化趨勢不一致。高濃度鎘脅迫下,抗氧化酶活性整體下降,嚴重時影響瓜苗正常生長,這可能是O2-和H2O2在植物體內的積累速度大于抗氧化酶的清除速度,活性氧持續積累,加劇了膜脂的過氧化[12 - 13 ]。不同作物乃至同一作物不同基因型在鎘脅迫下其抗氧化性酶活性變化規律均有差異[14, 18 - 19 ]。野生甜瓜幼苗長勢弱,葉片Pro含量最小,代謝活動最弱,而薄皮甜瓜和厚皮甜瓜Pro含量降幅較大,高濃度鎘嚴重抑制了可溶性蛋白的合成。MDA含量反映細胞膜脂過氧化的程度,厚皮甜瓜MDA含量在各處理下均最高,證明其在鎘脅迫下細胞膜脂過氧化程度重,植株生長勢強,代謝旺盛,鎘脅迫產生的膜損傷也重。
本試驗結果表明,脅迫下甜瓜幼苗葉片中SOD、POD、CAT、APX活性均低于對照不添加Cd2+。隨著Cd2+濃度增大,薄皮甜瓜和厚皮甜瓜幼苗葉片的SOD活性大幅下降,而野生甜瓜幼苗葉片的SOD活性略有回升,且在Cd2+中高濃度(50~100 mg/L)時均以野生甜瓜幼苗葉片的SOD活性最高,薄皮甜瓜的幼苗葉片的SOD活性最低。各處理下野生甜瓜幼苗葉片的POD活性均最高,薄皮甜瓜幼苗葉片的POD活性均最低,隨Cd2+脅迫濃度的增大,不同甜瓜材料幼苗葉片的POD活性均下降,但降幅不大。Cd2+脅迫濃度為0 mg/L時,薄皮甜瓜幼苗葉片的CAT活性最高,野生甜瓜幼苗葉片的CAT活性最低;隨Cd2+濃度的增大,薄皮甜瓜幼苗葉片的CAT活性降幅最大。各處理下野生甜瓜幼苗葉片的APX活性均最高,薄皮甜瓜幼苗葉片的APX活性最低,而野生甜瓜幼苗葉片的APX活性隨Cd2+濃度增大降幅較大。當Cd2+脅迫濃度為0 mg/L時,野生甜瓜幼苗葉片的Pro含量最低,薄皮甜瓜幼苗葉片的Pro含量最高;隨Cd2+脅迫濃度的變大,野生甜瓜幼苗葉片的Pro含量呈變化不明顯,薄皮甜瓜則逐步遞減趨勢,厚皮甜瓜呈先降后升又略下降趨勢。各處理下均以厚皮甜瓜幼苗葉片的MDA含量最高,隨Cd2+脅迫濃度的變大,野生甜瓜幼苗葉片的MDA含量呈先降后升趨勢,薄皮甜瓜的MDA含量呈逐步遞減趨勢,厚皮甜瓜的MDA含量呈先升后降又略上升趨勢。在相同Cd2+脅迫濃度處理下,野生甜瓜根系活力最強,薄皮甜瓜根系活力降幅最大。由此可見,鎘脅迫下薄皮甜瓜葉片的酶活性降幅較大,厚皮甜瓜次之,野生甜瓜葉片的酶活性降幅相對較小,且各處理下均以野生甜瓜根系活力最強,這說明參試的薄皮甜瓜對鎘脅迫較敏感,野生甜瓜抗鎘性較厚皮甜瓜、薄皮甜瓜強,可進一步開展野生材料抗鎘分子機理研究,為轉育出抗鎘核心甜瓜種質奠定基礎。
參考文獻:
[1] 張 軍,束文圣. 植物對重金屬鎘的耐受機制[J]. 植物生理與分子生物學學報,2006,32(1):1-8.
[2] 趙菲佚,翟祿新,陳 荃,等. Cd Pb復合處理下對植物膜的傷害初探[J]. 蘭州大學學報,2002,38(2):115-120.
[3] 史 靜, 潘根興, 夏運生, 等. 鎘脅迫對兩品種水稻生長及抗氧化酶系統的影響[J]. 生態環境學報, 2013,22(5): 832-837.
[4] 柯玉琴,潘廷國. NaCl 脅迫對甘薯葉片葉綠體超微結構及一些酶活性的影響[J]. 植物生理學報,1999,25(3):229-233.
[5] 張涵洋,李 愛,張衛華,等. 鎘脅迫對西瓜幼苗生長及其葉片解剖結構和生理特性的影響[J]. 西北植物學報,2023,43(3):401-409
[6] 李貞霞,任秀娟,孫涌棟. 南瓜對鎘的吸收積累特性研究[J]. 生態科學,2006,25(5):423-425.
[7] 姜曉君,蔣 英. 鎘脅迫對黃瓜種子萌發及幼苗生理指標的影響[J]. 天津農業科學,2019,25(10):27-30
[8] 張永平,范紅偉,楊少軍,等. 外源水楊酸對鎘脅迫甜瓜幼苗生長與光合氣體交換和葉綠素熒光特性的影響[J]. 西北植物學報2014,34(4):0778-0785.
[9] 張永平,沈若剛,姚雪琴,等. 鎘脅迫對甜瓜幼苗抗氧化酶活性和光合作用的影響[J]. 中國農學通報 2015,
31(34):82-88.
[10] 李合生. 植物生理生化實驗原理與技術[M]. 北京: 高等教育出版社,2000.
[11] 鄒 琦. 植物生理學實驗指導[M]. 北京:中國農業出版社,2000.
[12] 任艷芳,何俊瑜,劉 暢,等. 鎘脅迫對萵苣幼苗生長及抗氧化酶系統的影響[J]. 生態環境學報,2009,
18(2):494-497
[13] 汪 洪,趙士誠,夏文建,等. 不同濃度鎘脅迫對玉米幼苗光合作用、脂質過氧化和抗氧化酶活性的影響[J]. 植物營養與肥料學報,2008, 14(1):36-42.
[14] 孫光聞,朱祝軍,方學智. 不同鎘水平對白菜生長及抗氧化酶活性的影響[J]. 園藝學報,2004,31(3):378-380.
[15] 權建華,劉玉玲,許可紅,等. 辣椒根系分泌物對辣椒種子萌發和幼苗生長的影響[J]. 寒旱農業科學,2023,2(4):334-338.
[16] 白銳琴. 2,4-表油菜素內酯對干旱脅迫下辣椒葉片生理指標及果實品質的影響[J]. 寒旱農業科學,2024,
3(2):141-145.
[17] 牛軍強,尹曉寧,董 鐵,等. 基于主成分分析評價間伐改形對蘋果葉片生理指標的影響[J]. 寒旱農業科學,2023,2(7):631-638.
[18] YANG J X, GUO H T, MA Y B, et al. Genotypic variations in the accumulation of Cd exhibited by different vegetables[J]. Journal of Environmental Sciences, 2010, 22(8) : 1246-1252.
[19] MOHAPATRAL S, DEY S K. Physiological response of Hydrilla verticillata(l.f.) royle exposed to cadmium stress[J]. Journal of life sciences,2012(6): 55-60.
