氮素形態對穿心蓮氮吸收、分配和內酯成分積累的影響
2024-12-31 00:00:00簡少芬林楊萬斯鐘楚劉寒
廣西植物 2024年11期


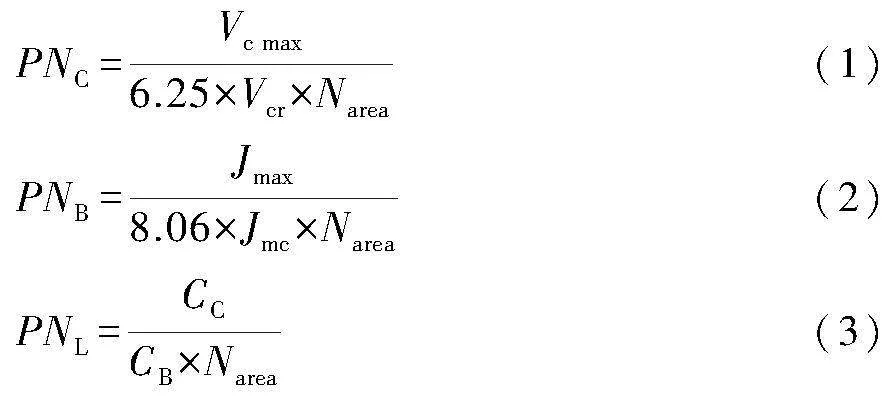


摘 要: "為了明確藥用植物穿心蓮(Andrographis paniculata)對不同氮素形態的利用特征及其與穿心蓮內酯成分積累的關系,以硝態氮(NN)、銨態氮(AN)、酰胺態氮(UN)和氨基酸態氮(GN)為單一氮源,采用15N同位素示蹤和生理生化分析,研究了氮素形態對穿心蓮不同生長時期(快速生長期、拔節期、現蕾期、開花期)氮素吸收、分配和穿心蓮內酯成分含量的影響。結果表明:(1)葉片和根中氮含量隨生長時期逐漸下降,NN處理氮含量較低。(2)穿心蓮氮吸收速率在營養生長期較高,生殖生長期急速下降,穿心蓮對AN、UN和GN的吸收速率較高。(3)現蕾期葉片氮分配比例減少,莖氮分配比例增加,與NN處理相比,該時期AN、UN和GN處理降低了葉片氮分配比例,而提高了莖和根氮分配比例。(4)快速生長期NN處理的光合作用最大羧化速率和最大電子傳遞速率較低,葉片氮在羧化系統和生物能學組分中的分配比例也較低;UN和AN處理分別在現蕾期和開花期降低了葉片氮在羧化系統中的分配。(5)AN、UN和GN提高了穿心蓮內酯和脫水穿心蓮內酯含量,降低了現蕾期和開花期14-去氧穿心蓮內酯含量,不同氮素形態對新穿心蓮內酯的影響較小。(6)穿心蓮內酯和新穿心蓮內酯含量與葉、莖、根氮含量,氮吸收速率及葉和根中氮分配比例顯著負相關,而與莖中氮分配比例顯著正相關,14-去氧穿心蓮內酯則相反。綜上認為,營養生長期是穿心蓮氮吸收的主要時期,穿心蓮能更好地利用銨態氮、酰胺態氮和氨基酸態氮,并通過優化氮分配促進穿心蓮內酯成分積累。
關鍵詞: 氮素形態, 氮吸收, 氮分配, 穿心蓮內酯, 穿心蓮
中圖分類號: "Q945.1
文獻標識碼: "A
文章編號: "1000-3142(2024)11-2076-13
Effects of nitrogen forms on nitrogen uptake and
allocation and andrographolide components
accumulation in Andrographis paniculata
Abstract: "Nitrogen (N) is a mineral nutrient with the highest demand for plants, and it is also a vital abiotic factor that limits plant yield and quality.
In order to clarify the utilization characteristics of different N forms of Andrograhis paniculata and its relationship with the accumulation of andrographolide content, with nitrate nitrogen (NN), ammonium nitrogen (AN), amide nitrogen (urea, UN) and amino acid nitrogen (glycine, GN) as sole nitrogen source and combining 15N isotope tracing and physiological and biochemical analysis, the effects of nitrogen form on N uptake and distribution and andrographolide content at different growth stages (rapid growth stage, jointing stage, budding stage, flowering stage) of A. paniculata were studied.
The results were as follows: (1) N contents in leaf and root decreased gradually with the growth stage, and it was lower in the NN treatment. (2) The uptake rate of N was higher in the vegetative growth stage, and rapidly decreased in the reproductive growth stage. The uptake rates of AN, UN and GN were higher than that of NN in A. paniculata. (3) The allocation ratio of N in leaf was decreased, while the allocation ratio of N in stem was increased at budding stage. In comparison to the NN treatment, AN, UN, and GN treatments reduced N allocation ratio in leaf, but increased the ratios in stem and root at this growth stage. (4) At rapid growth stage, the maximum carboxylation rate and maximum electron transport rate of photosynthesis were lower in the NN treatment, as well as the proportion of leaf N allocation in the carboxylation system and bioenergetics components. The allocation of leaf N in the carboxylation system in UN and AN treatments were reduced at budding and flowering stages, respectively. (5) AN, UN and GN increased andrographolide and dehydroandrographolide contents, and decreased 14-deoxyandrographolide content at budding and flowering stages, while neoandrographolide was slightly affected by N forms. (6) The contents of andrographolide and neoandrographolide were significantly negatively correlated with N contents in leaf, stem and root, N uptake rate, and N allocation ratios in leaf and root, while they were significantly positively correlated with N allocation ratio in stem. The opposite was true for 14-deoxyandrographolide. Taken together, the results indicate that the vegetative growth stage is the main stage for N uptake in "A. paniculata, and that "A. paniculata can better utilize AN, UN, and GN, and promote the accumulation of andrographolide components by optimizing N allocation. The results provide a theoretical reference for N management of A. paniculata.
Key words: nitrogen form, nitrogen uptake, nitrogen allocation, andrographolide, Andrographis paniculata
氮是植物需求量最大的礦質營養元素,也是限制植物產量和品質的關鍵非生物因子。植物所需的氮主要來源于土壤,以無機態的硝酸根(NO3-)和銨根(NH4+)離子為主,也包括一些小分子的氨基酸和多肽等有機態氮(曹小闖等,2015)。不同的氮素形態不僅對植物生長和生理特征有較大影響(Guo et al., 2007),也是影響藥用植物次生代謝物積累的關鍵因子。不同氮素形態對植物次生代謝物積累的影響有很大差別,與次生代謝物種類、植物對氮素形態的偏好性及氮形態的組成有關。如蕁麻多酚物質(Biesiada et al., 2009)、甜葉菊的甜菊苷(Sun et al., 2021)、醫用大麻的大麻素和萜類物質(Saloner amp; Bernstein, 2022)等在硝態氮條件下積累較銨態氮條件下高,而在茶樹(Camellia sinensis)(Yang et al., 2018)、天門冬(梁娟等,2018)、菘藍(唐曉清等,2017)等植物中,較高比例的銨態氮更有利于次生代謝物質的積累。不同氮素形態可能通過調控藥用植物初生代謝(如氨基酸合成、三羧酸循環和糖酵解)和次生代謝的關系以及次生代謝通路酶編碼基因的表達影響次生代謝物的積累(張強等,2018;羅佳琪和付立忠,2020;Chen et al., 2023)。然而,其具體的影響機制還缺乏系統研究。不同氮形態對植物體內活性成分影響的差異在一定程度上與植物對不同形態氮的吸收利用差異有關(晏楓霞等,2010)。但目前為止,關于藥用植物對不同氮素形態的吸收利用特征及其對次生代謝物積累的影響還鮮有報道。
穿心蓮(Andrographis paniculata)是我國重要的南藥大宗藥材之一,在全球草藥市場占有重要地位。它起源于南亞的印度和斯里蘭卡,廣泛應用于南亞、東南亞和東亞傳統醫藥,具有祛熱解毒,消炎止痛等功效,是“天然抗生素藥物”。筆者前期研究表明,氮素形態影響穿心蓮主要活性成分穿心蓮內酯的積累,銨態氮和有機態的酰胺態氮和氨基酸態氮相對于硝態氮能提高穿心蓮內酯含量(Zhong et al., 2021)。鑒于植物對不同氮素形態的吸收利用普遍存在差異,因此推測不同氮素形態對穿心蓮內酯成分積累的影響可能與穿心蓮對不同形態氮源的吸收、分配差異有關。本研究以4種氮素形態作為唯一氮源,采用無土基質培育穿心蓮植株,通過生理生化研究方法和15N穩定同位素示蹤技術,擬探討穿心蓮不同生長時期對各氮素形態的吸收利用特征及其與穿心蓮內酯成分積累的關系,以期為穿心蓮氮素養分的科學管理提供理論依據。
1 材料與方法
1.1 試驗材料和處理
以藥用植物穿心蓮為材料,原始種子由廣西藥用植物園種子庫提供,由鐘楚博士鑒定為爵床科穿心蓮屬植物穿心蓮。經本課題組篩選和純化后獲得性狀一致、穩定的株系,用于后續研究。在培養室晝夜恒溫(約28 ℃)條件下,將穿心蓮種子放在裝有濕濾紙的培養皿中發芽,過程中保持濾紙完全濕潤。光照周期為14 h光照/10 h黑暗,200 μmol·m-2·s-1的光合有效輻射(photosynthetically active radiation, PAR),相對濕度60%。大約10 d后,將發芽的幼苗移植到裝有蛭石、珍珠巖和品氏泥炭土(體積比約為4∶1∶1)混合基質的育苗盤中繼續生長,定期澆水防止幼苗干旱,直到5對葉齡。選取健康、長勢一致的幼苗移植到裝有蛭石和珍珠巖(4∶1)的花盆中進行N形態處理。每盆1株,移栽前抖凈根上附著的基質,每處理48盆(株)。將植物置于光溫培養室中進行生長,光、溫環境同上。
分別以2 mmol·L-1 KNO3和2 mmol·L-1 Ca(NO3)2、6 mmol·L-1 NH4Cl、3 mmol·L-1 尿素 [CO(NH2)2]以及6 mmol·L-1甘氨酸提供氮源,分別作為硝態氮(nitrate nitrogen,NN)、銨態氮(ammonium nitrogen,AN)、酰胺態氮(urea nitrogen,UN)和氨基酸態氮(glycine nitrogen,GN)處理。營養液中的其他礦物質營養素含有1 mmol·L-1 NaH2PO4、2 mmol·L-1 KCl、2 mmol·L-1 CaCl2、1 mmol·L-1 MgSO4、18 μmol·L-1 H3BO3、0.15 μmol·L-1 ZnSO4、0.15 μmol·L-1 CuSO4、0.52 μmol·L-1 (NH4)6Mo7O24、9.5 μmol·L-1 MnSO4和36 μmol·L-1 Fe-EDTA。為了維持各處理中K+和Ca2+濃度一致,NN處理不加KCl和CaCl2,K+和Ca2+由KNO3和Ca(NO3)2提供。將溶液的pH值調至5.8~6.0,并在營養液中加入10 mg·L-1氨芐青霉素以抑制微生物繁殖。每周澆營養液2次,每盆100 mL。每處理長勢一致的植株有40株左右,可滿足取樣的需求。處理30、50、70、85 d后取樣分析。
1.2 測定方法
1.2.1 光合作用CO2響應曲線 分別在移栽后30 d(快速生長期)、50 d(拔節期)、70 d(現蕾期)和85 d(開花期)選取長勢一致的植株3株,采用LI-6400XT便攜式光合測定系統,測定主莖上第9至第12葉位(不同時期葉位不同,但同一時期各處理葉位相同)葉片光合作用CO2響應特征。使用內置LED光源,紅藍光比例為9∶1,光合有效輻射(PAR)為1 500 μmol·m-2·s-1,葉室溫度25 ℃,相對濕度60%~70%,CO2濃度梯度為50、100、150、200、300、400、600、800、1 000、1 200、1 500 μmol·mol-1。
將葉片置于葉室中,在1 500 μmol·m-2·s-1 PAR、400 μmol·mol-1 CO2濃度下適應至少20 min,待各參數穩定后,開始測量各CO2濃度下的光合氣體交換參數。根據FvCB模型(Farquhar et al., 1980)對CO2響應曲線進行擬合,計算最大羧化速率(Vcmax)和最大電子傳遞速率(Jmax)。
1.2.2 葉綠素含量、比葉重和比葉氮測定 測定完光合作用之后,用直徑6 mm的打孔器在葉片中部主脈兩側各取4片葉圓片置于10 mL離心管中,加入5 mL丙酮∶乙醇(1∶1,體積比)混合液,在室溫暗處浸提24 h,直至葉片完全變白。測定浸提溶液在663 nm和645 nm處的吸光值,計算葉綠素a、b和總葉綠素含量(張憲政,1986)。
另用打孔器在葉片兩側各取5片葉圓片,70 ℃烘干至恒重,稱重后,葉片用H2SO4-H2O2在260 ℃消煮,靛酚藍比色法測定總氮含量(呂偉仙等,2004)。比葉重(specific leaf weight,SLM)和比葉氮(specific leaf nitrogen,SLN)分別為單位葉片面積的重量和氮含量。
1.2.3 15N同位素標記和測定 分別在快速生長期、拔節期、現蕾期和開花期各處理選取長勢一致的植株4株,取樣前6 d,將營養液中的氮源分別換成15N標記的相應氮源,15N原子豐度均為10%。每3 d處理1次,處理結束后,將植株根系完整取出,先用自來水沖洗干凈附著的蛭石和珍珠巖,再用1 mmol·L-1 CaCl2沖洗1 min,最后用去離子水沖洗干凈。把植株根、莖、葉各部位分開,分別裝入牛皮紙袋后105 ℃殺青30 min,75 ℃烘干至恒重,稱重后將各部位研磨成細粉。采用連續流動同位素比值質譜儀偶聯碳氮元素分析儀(ANCA-MS,PDZ-Eropa)測定各部位15N原子豐度,并計算15N含量和吸收速率。
1.2.4 植株生物量和總氮含量測定 生物量和植株總氮含量采用1.2.3樣品進行測定。總氮含量采用靛酚藍比色法測定(呂偉仙等,2004)。
1.2.5 穿心蓮內酯成分含量 葉片干樣中穿心蓮內酯、脫水穿心蓮內酯、14-去氧穿心蓮內酯和新穿心蓮內酯含量的測定參考王雨霞等(2022)的方法,采用1.2.3樣品進行測定。
1.3 數據處理與統計
光合氮(photosynthetic nitrogen,PN)分配:將光合機構分為羧化系統(PNC)、生物能學(PNB)和集光系統(PNL)3部分(史作民等,2015)。各部分氮的分配比例按下式計算:
式中:Vc max為最大羧化速率(μmol·m-2·s-1);Jmax為最大電子傳遞速率(μmol·m-2·s-1),由CO2響應曲線根據FvCB模型擬合得到;CC為葉綠素含量(mmol·m-2);Narea為單位葉片面積氮含量(SLN,g·m-2);Vcr為Rubisco比活,取值為20.78 μmol CO2·g-1 Rubisco·s-1;Jmc為單位Cyt f的最大電子傳遞速率,取值156 μmol e-·μmol-1 Cyt f·s-1;CB為捕光系統中葉綠素與氮的比值,取值2.15 mmol·g-1 N(史作民等,2015)。
采用單因素方差分析和LSD法多重比較處理間差異顯著性。數據以平均值±標準差(n=3或4)表示,數據統計分析在SPSS 19.0數據處理系統中進行,以Microsoft Excel 2023進行繪圖。
2 結果與分析
2.1 不同氮源對穿心蓮生長的影響
如圖1所示,用不同氮源處理5對葉齡穿心蓮植株30 d(圖1:A,S1)、50 d(圖1:B,S2)、70 d(圖1:C,S3)和85 d(圖1:D,S4)的表型沒有太大差別,硝態氮(NN)處理的植株進入開花期稍晚。各處理前3個時期株高差異較小,而在開花期NN處理株高顯著高于其他3個處理(圖1:E)。各處理植株生物量在S1時期沒有顯著差異,而S2時期NN處理顯著高于其他3個處理,S3時期AN和UN的生物量較低,而S4時期以AN的生物量最低,顯著低于其他3個處理(圖1:F)。葉片生物量占地上部生物量的比例(葉重比)隨生長時期變化而逐漸下降,S1時期以UN處理最低,而S2和S3時期以AN處理最低,S4時期各處理沒有顯著差異(圖1:G)。
2.2 氮源對穿心蓮氮吸收、積累和分配的影響
各處理植株葉片氮含量隨生長時期呈逐漸下降的變化。NN處理各時期葉片氮含量相對較低,在S1和S3時期顯著低于其他3個處理(圖2:A)。莖中氮含量在整個生育期變化不大。S1時期NN和AN相對較低,與UN差異顯著;S2時期AN最低,與NN和GN差異顯著;S3時期AN最低,與UN差異顯著;S4時期各處理差異不顯著(圖2:B)。根中氮含量隨生長時期呈逐漸下降的變化。UN處理在各時期均最高,除S2時期,NN處理在其他3個時期均最低;NN和AN處理在S1、S2和S4時期顯著低于UN和GN(S4時期AN除外),而在S3時期NN和GN顯著低于AN和UN(圖2:C)。植株總氮含量隨生長時期呈逐漸下降變化,NN處理在各時期普遍較低(圖2:D)。
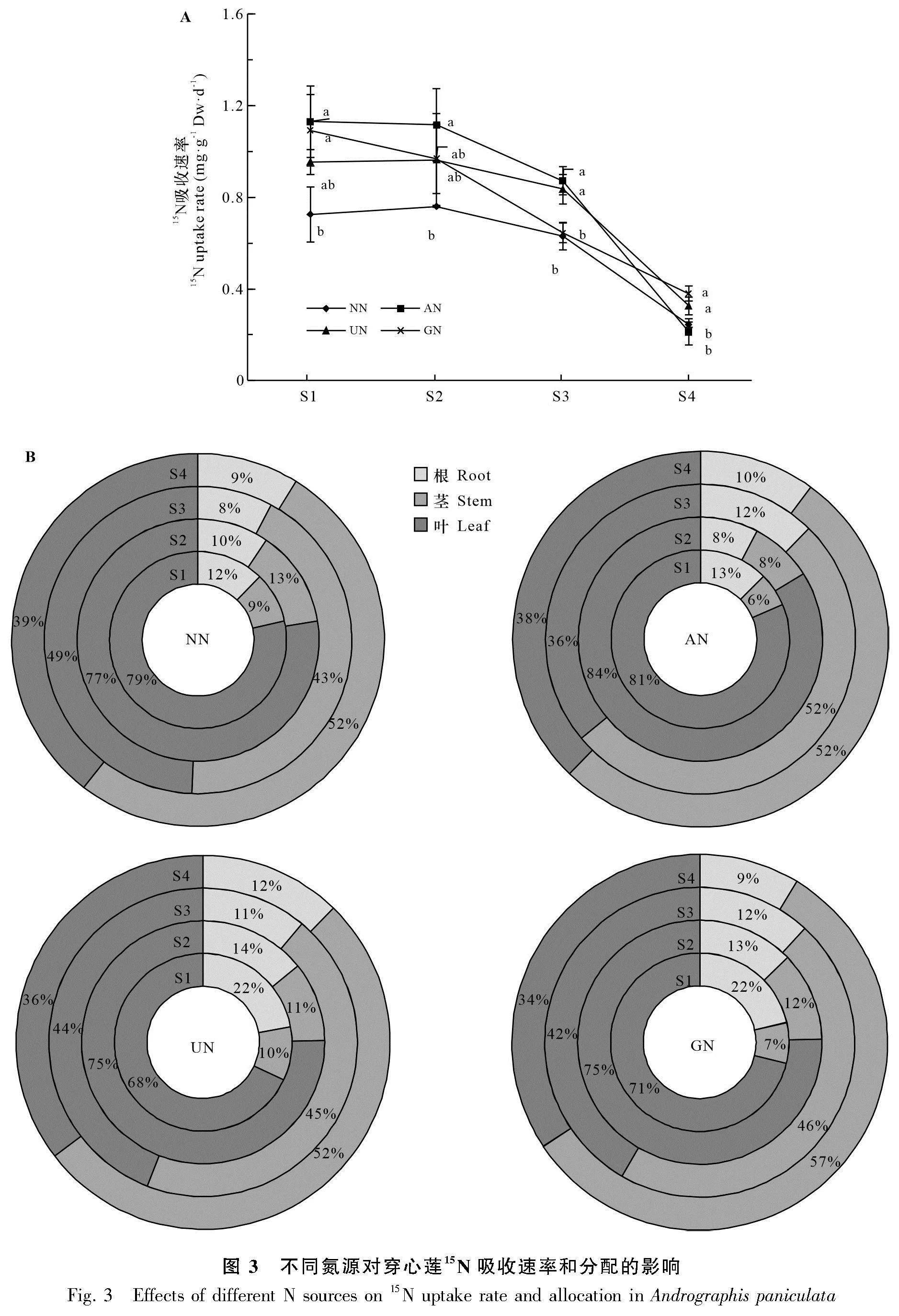
以15N穩定同位素進行示蹤,測定了不同氮源處理下穿心蓮的氮吸收速率。如圖3:A所示,在前2個生長時期穿心蓮對氮的吸收速率較高, 隨后急劇下降。各生長時期穿心蓮對硝態氮的15N吸收速率始終最低,S1-S3時期顯著低于AN處理,而S3和S4時期顯著低于UN處理,S1和S4時期顯著低于GN處理。結果說明,穿心蓮可以更好地利用銨態氮、酰胺態氮和氨基酸態氮。
通過計算15N在各器官中的分配比例,結果表明,S1和S2時期各氮源處理穿心蓮吸收的15N主要分布在葉片中,而S3和S4時期在莖中的分配比例大幅增加,甚至高于葉片中分配的比例(圖3:B)。S1時期AN處理與NN處理的15N分配較為相似,而UN與GN處理相似。NN和AN處理穿心蓮葉片中15N的分配比例明顯高于UN和GN處理,而根中15N的分配比例則明顯低于UN和GN處理。S2時期NN和AN各器官15N的分配比例沒有太大變化,而UN和GN處理根中15N的分配比例明顯下降,葉片和莖中15N的分配比例有所增加。S3時期AN、UN和GN處理葉片中15N的分配比例明顯較NN處理低,而根中分配比例則明顯較高。S4時期各處理各器官中15N的分配比例沒有明顯差異。
2.3 氮源對穿心蓮光合氮分配的影響
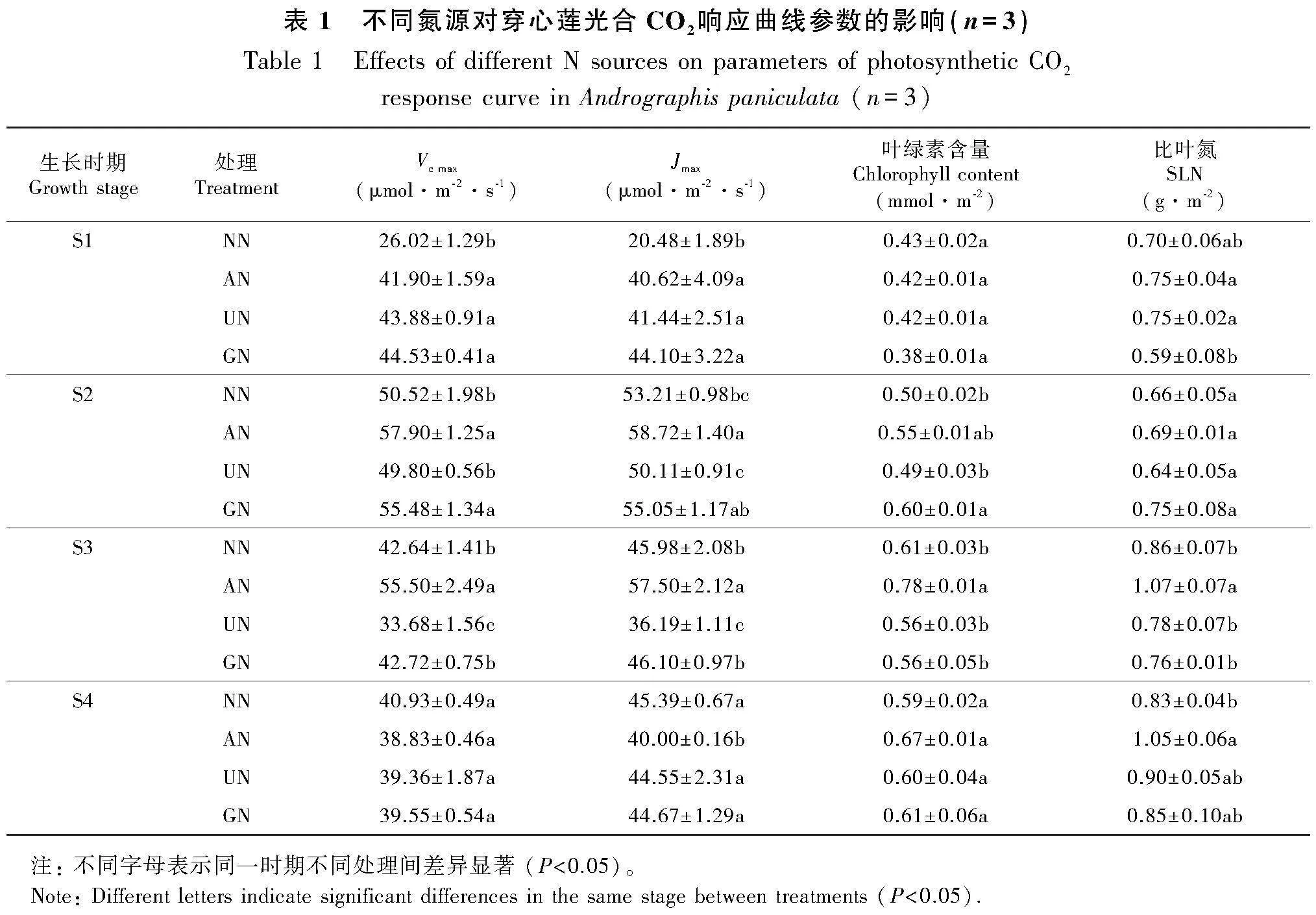
不同氮源處理對不同生長時期穿心蓮光合作用CO2響應特征有明顯影響。由表1可知:AN、UN和GN處理在S1時期Vc max和Jmax均顯著高于NN處理,S2時期AN處理Vc max和Jmax最高,顯著高于UN和NN處理,S3時期AN處理的Vc max和Jmax仍顯著高于其他3個處理,而S4時期各處理之間沒有顯著差異;各處理葉綠素含量在生長前期較低,后期較高, S1和S4時期處理間差異不顯著,S2時期GN處理葉綠素含量最高,顯著高于NN和UN處理,S3時期AN處理葉綠素含量顯著高于其他處理;各處理比葉氮(SLN)在S1和S2時期相對較低,而S3和S4時期相對較高,S1時期AN 和UN處理顯著高于GN處理,S3和S4時期AN處理最高。
由圖4的葉片氮在光合機構中的分配情況可知,分配于光合機構中的葉片氮比例為52.8%~88.7%,隨生長時期呈先升高后降低的趨勢,S2時期比例最高,與該時期有較高的Vc max和Jmax結果一致。葉片氮在光合機構3個組分中分配的比例以生物能學(PNB)最低,不到10%。S1和S2時期各處理葉片氮在集光系統(PNL)和羧化系統(PNC)中分配的比例相當,但S1時期NN處理PNC中氮的分配比例較其他處理低。S3和S4時期各處理葉片氮主要分配于PNL中,處理間相差不大。S3時期UN處理和S4時期AN處理的PNC氮分配比例明顯低于同時期其他處理。
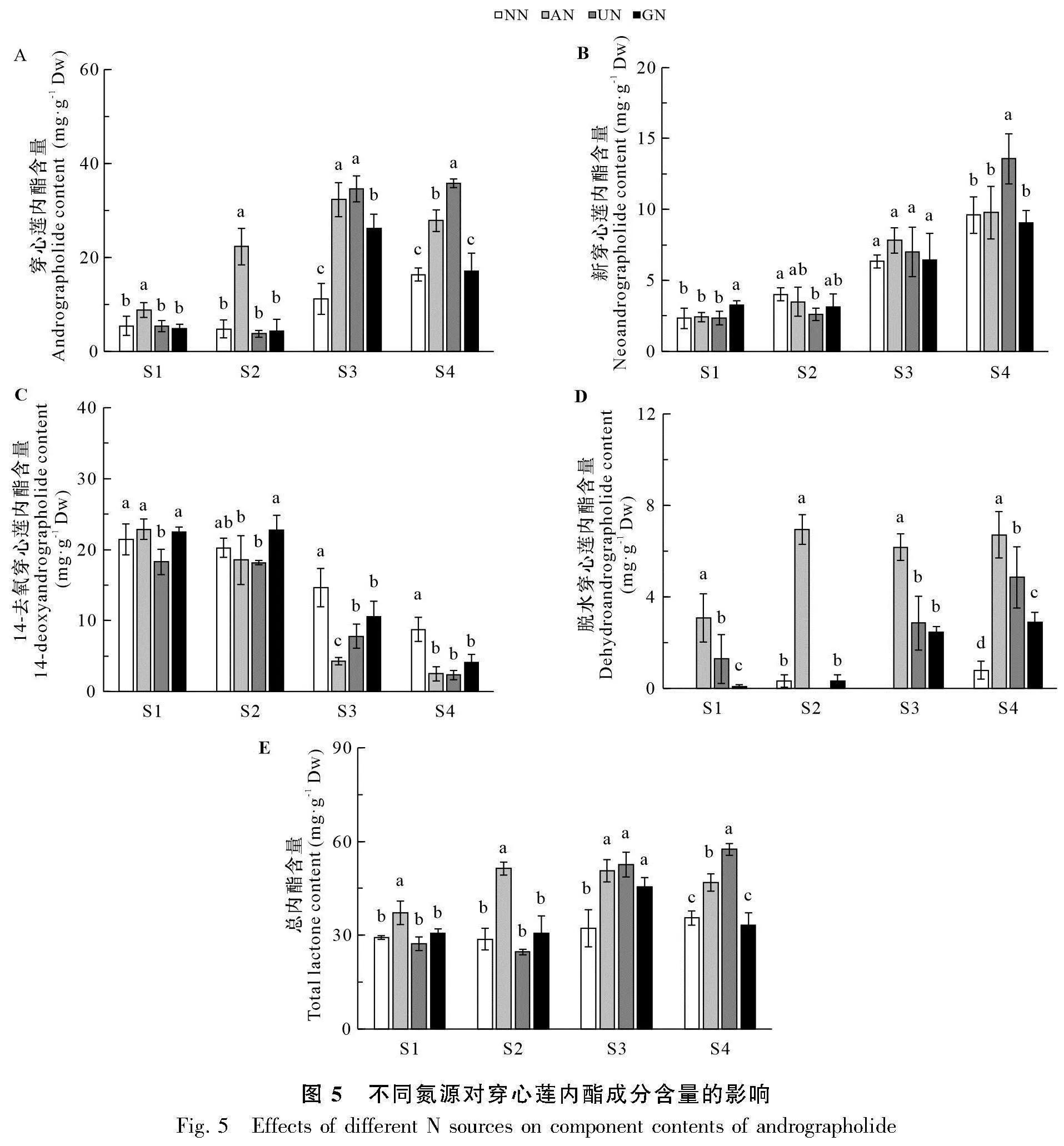
2.4 氮形態對穿心蓮內酯成分積累的影響
如圖5:A所示,AN處理穿心蓮內酯含量在各時期均較高,其中S1和S2時期顯著高于其他3個處理,而其他3個處理差異不顯著。尤其是S2時期,AN處理的穿心蓮內酯含量超過其他處理的4倍。S3時期UN和GN處理的穿心蓮內酯含量迅速上升,其中UN與AN差異不顯著,GN處理顯著低于AN和UN,但都顯著高于NN。該時期AN和UN處理的穿心蓮內酯含量是NN處理的3倍左右。S4時期UN處理的穿心蓮內酯含量仍維持較高水平,顯著高于其他處理,而GN處理的穿心蓮內酯含量下降,與NN差異不顯著,二者顯著低于AN處理。
各處理新穿心蓮內酯含量隨生長時期逐漸升高。S1時期以GN最高,顯著高于其他處理;S2時期UN最低,顯著低于NN處理;S3時期各處理沒有顯著差異,而S4時期UN處理最高,顯著高于其他3個處理(圖5:B)。14-去氧穿心蓮內酯含量隨生長時期逐漸下降,NN處理各時期相對較高,尤其在S3和S4時期顯著高于其他3個處理(圖5:C)。各時期NN處理的脫水穿心蓮內酯含量都較低,在S1和S3時期未檢出;AN處理各時期脫水穿心蓮內酯含量都最高,顯著高于其他3個處理;UN和GN處理的脫水穿心蓮內酯含量在前2個時期較低,而后2個時期相對較高(圖5:D)。AN處理總內酯含量在S1和S2時期顯著高于其他3個處理,而其他3個處理之間沒有顯著差異;S3時期AN與UN和GN差異不顯著,三者顯著高于NN處理;S4時期以UN處理最高,其次為AN,NN和GN處理最低(圖5:E)。總體來看,銨態氮、酰胺態氮和氨基酸態氮有利于提高穿心蓮內酯及總內酯含量。
2.5 穿心蓮內酯成分含量與氮吸收和分配的相關性分析
相關性分析結果(表2)表明,穿心蓮內酯和新穿心蓮內酯含量與根、莖、葉氮含量以及葉片和根中氮分配比例、氮吸收速率負相關,而與莖中氮分配比例極顯著正相關。14-去氧穿心蓮內酯含量與穿心蓮內酯含量和新穿心蓮內酯含量極顯著負相關,因此其與葉和根的氮含量、氮分配比例、氮吸收速率顯著或極顯著正相關,而與莖中氮分配比例極顯著負相關。總內酯含量與穿心蓮內酯含量相似,與莖和根中氮含量及葉中氮分配比例極顯著負相關,與莖中氮分配比例極顯著正相關。結果說明,降低植株氮含量,增加氮在莖中的分配比例可以有效提高穿心蓮內酯及總內酯含量。
各器官氮含量之間相互呈極顯著正相關,而莖中氮分配比例與葉和根中氮分配比例均呈極顯著負相關,尤其與葉中氮分配比例的相關性最強。葉和根中較高的氮含量也會提高氮在它們當中的分配比例,而不利于莖中氮分配比例的增加。
3 討論與結論
3.1 穿心蓮對不同氮源的吸收和利用特征
植物可以直接利用無機態氮和有機態氮,但對不同氮素形態的吸收利用存在較大差異,大部分陸地植物以吸收硝態氮為主,只有少數植物如水稻、茶樹、馬鈴薯等以利用銨態氮為主(郝凱等,2020)。本研究中通過15N示蹤發現,穿心蓮對銨態氮、酰胺態氮和氨基酸態氮的吸收速率要明顯高于對硝態氮的吸收速率,尤其對銨態氮的吸收速率最高,表現出對銨態氮有一定的偏好性。同時也發現,銨態氮及酰胺態氮、氨基酸態氮處理穿心蓮葉片在現蕾期以前均有較高的氮含量。以上結果說明,穿心蓮能更好地利用這3種氮形態。值得注意的是,銨態氮處理植株雖然生物量相對較少,但并未出現如葉片黃化、植株生長受抑制,甚至致死等典型的銨毒癥狀(Jian et al., 2018; Hachiya et al., 2021)。銨抑制穿心蓮生物量積累的原因仍有待進一步研究。
利用15N同位素示蹤比較不同生長時期氮素
吸收速率的變化可知,拔節期(S2)之前是穿心蓮氮素吸收的主要時期,進入現蕾期(S3)之后氮素吸收速率開始大幅下降。結果表明,拔節期之前的營養生長期是穿心蓮氮需求的關鍵時期,而進入生殖生長后對氮素的需求急劇減少。15N同位素示蹤結果還表明,現蕾期是植株水平上氮素重新分配的關鍵時期。因氮吸收減少,葉片中氮分配比例下降,植株將氮轉移至莖和根,尤以銨態氮、酰胺態氮和氨基酸態氮處理的植株該時期葉片中的氮向根和莖中轉運更加明顯。此外,這3種氮源處理的植株根中氮含量也普遍較硝態氮處理高,與15N測定結果一致。綜上認為,穿心蓮生產上氮肥應主要在營養生長期施用,生殖生長期因氮吸收速率下降,植株體內氮以重新分配為主,應避免在生殖生長期施用氮肥。
3.2 氮源對穿心蓮內酯成分的影響
本研究表明,不同氮素形態對穿心蓮內酯成分有較大影響。氮素形態對藥用植物不同活性成分的影響差異已有廣泛報道,但關于其機理還缺乏深入研究。根據碳/養分平衡假說,較高的氮含量通常導致藥用植物富含碳的次生代謝物質積累下降(羅佳琪和付立忠,2020)。本研究中穿心蓮內酯、新穿心蓮內酯和總內酯含量與葉、莖、根中的氮含量及氮吸收速率負相關。然而, 銨態氮、酰胺態氮和氨基酸態氮處理下植株葉片氮含量和氮吸收速率較硝態氮處理植株高且穿心蓮內酯含量也相對較高。相關性分析結果反映了各生長時期氮含量變化與穿心蓮內酯成分變化有關,而不同氮素形態處理之間葉片氮含量和氮吸收速率的差異不是造成內酯含量差異的主要原因。
光合產物是植物次生代謝的最初物質來源,維持較高的光合能力可以為次生代謝提供更多的同化物。銨態氮、酰胺態氮和氨基酸態氮處理的植株具有較強的光合能力,可能是導致穿心蓮內酯含量較高的原因之一。植物葉片中大部分的氮都存在于光合器官中參與光合作用(Makino et al., 2003)。因此,葉片氮含量及其在光合機構各組分中的分配對光合作用有重要影響(史作民等,2015)。營養生長期是植物干物質積累的重要時期。本研究中,銨態氮、酰胺態氮和氨基酸態氮處理的穿心蓮葉片始終具有相對較高的氮含量,而且在營養生長時期(S1和S2)具有較高的光合能力以及較高的Vc max和Jmax,表明這3種氮源處理的植株將更多的氮投入到光合作用中,光合作用受到的化學限制也較小。從氮在光合機構組分中的分配可以看出,S1時期硝態氮處理的植株光合氮在羧化系統和生物能學組分中分配的比例較其他處理低,可能是導致其光合能力較低的重要原因。
植物由營養生長向生殖生長的轉化期是氮素轉移的關鍵時期,也是穿心蓮內酯成分積累的重要時期(陳娟等,2014)。本研究中現蕾期(S3)穿心蓮內酯和新穿心蓮內酯含量開始大量積累。該時期銨態氮、酰胺態氮和氨基酸態氮處理葉片氮的分配比例較硝態氮處理明顯下降,而根和莖中的氮分配比例增加。相關性分析也表明,穿心蓮內酯和新穿心蓮內酯含量與葉片氮分配比例顯著負相關而與莖中氮分配比例顯著正相關,說明該時期葉片氮向莖中轉移增加對穿心蓮內酯積累有促進作用。葉片是氮同化的主要器官,尤其生長后期大量氮積累將增加碳水化合物消耗而不利于次生代謝物質積累(Zhong et al., 2021)。本研究結果在一定程度上符合碳/養分平衡假說的預測。氮在光合機構各組分中的分配也影響植物對氮的再利用(劉濤,2018)。水溶性蛋白較其他類型蛋白更易被降解而再利用。羧化系統中的氮主要以羧化酶為主,占水溶性蛋白的一半以上(Carmo-Silva et al., 2015)。羧化系統中維持較高的氮比例不僅降低植物光合效率(Mu et al., 2016),也降低葉片中氮的轉移和再利用。生長后期(S3和S4)銨態氮和酰胺態氮處理植株羧化系統氮的比例下降,可能有利于葉片中氮的轉移。
綜合以上分析,銨態氮、酰胺態氮和氨基酸態氮影響穿心蓮植株和葉片氮分配,提高穿心蓮營養生長期光合能力,促進生殖生長期植株葉片氮素向莖中的轉移,為葉片中穿心蓮內酯成分的合成與積累提供了物質基礎。
3.3 本研究的潛在意義
氮素形態通過調控藥用植物次生代謝物合成通路中酶基因的表達而影響活性成分積累(張強等,2018)。穿心蓮內酯屬于半日花烷型二萜內酯,其合成通路還尚未被解析。本研究中發現,14-去氧穿心蓮內酯含量與穿心蓮內酯含量極顯著負相關,銨態氮、酰胺態氮和氨基酸態氮不僅提高了穿心蓮內酯含量,還特異性地提高了脫水穿心蓮內酯含量,尤以銨態氮的效果更加明顯。結果說明,穿心蓮內酯含量的提高與14-去氧穿心蓮內酯和脫水穿心蓮內酯含量的變化有密切聯系。本研究結果支持了任俊澤(2023)最近提出的穿心蓮內酯合成的推測路徑,即由14-去氧穿心蓮內酯和脫水穿心蓮內酯通過單加氧反應形成穿心蓮內酯。細胞色素氧化酶P450(CYP450)家族蛋白酶催化單加氧反應,在藥用植物化學成分的修飾方面起到重要作用(Xu et al., 2015)。可以推測,銨態氮、酰胺態氮和氨基酸態氮可能誘導了某個(或某幾個)CYP450基因的上調表達,促進了14-去氧穿心蓮內酯和脫水穿心蓮內酯向穿心蓮內酯的轉化。長期以來,雖然有較多關于穿心蓮 CYP450基因的研究(Garg et al., 2015;Liang et al., 2020;孫銘陽等,2022),但催化穿心蓮內酯合成的關鍵CYP450基因還未見報道。本研究為穿心蓮內酯合成通路中CYP450基因的挖掘奠定了基礎。
參考文獻:
BIESIADA A, WOOSZCZAK E, SOKETOWSKA A, et al., 2009. The effect of nitrogen form and dose on yield, chemical composition and antioxidant activity of stinging nettle (Urtica dioica L.) [J]. Herba Pol, 55(3): 84-91.
CAO XC, WU LH, MA QX, et al., 2015. Advances in studies of absorption and utilization of amino acids by plants: A review" [J]. Chin J Appl Ecol, 26(3): 919-929." [曹小闖, 吳良歡, 馬慶旭, 等, 2015. 高等植物對氨基酸態氮的吸收與利用研究進展 [J]. 應用生態學報, 26(3): 919-929.]
CARMO-SILVA E, SCALES JC, MADGWICK PJ,et al., 2015. Optimizing rubisco and its regulation for greater resource use efficiency [J]. Plant Cell Environ, 38: 1817-1832.
CHEN J, GU W, DUAN JA, et al., 2014. Study on active components of Andrographis paniculata at various growth periods and differential expression of key enzyme gene [J]. Chin Tradit Herbal Drugs, 45(21): 3149-3152." [陳娟, 谷巍, 段金廒, 等, 2014. 不同生長期穿心蓮活性成分及關鍵酶基因差異表達研究 [J]. 中草藥, 45(21): 3149-3152.][JP2]
CHEN Y, BAI Y, ZHANG ZR, et al., 2023. Transcriptomics and metabolomics reveal the primary and secondary metabolism changes in Glycyrrhiza uralensis with different forms of nitrogen utilization [J]. Front Plant Sci, 14: 1229253.
FARQUHAR GD, VON CAEMMERER S, BERRY JA, 1980. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species [J]. Planta, 149: 78-90.
GARG A, AGRAWAL L, MISRA RC, et al., 2015. Andrographis paniculata transcriptome provides molecular insights into tissue-specific accumulation of medicinal diterpenes [J]. BMC Genomics, 16(1): 659.
GUO SW, ZHOU Y, GAO YX, et al., 2007. New insights into the nitrogen form effect on photosynthesis and photorespiration [J]. Pedosphere, 17(5): 601-610.
HACHIYA T, INABA J, WAKAZAKI M, et al., 2021. Excessive ammonium assimilation by plastidic glutamine synthetase causes ammonium toxicity in Arabidopsis thaliana [J]. Nat Commun, 12: 4944.
HAO K, JIA LG, QIN YL, et al., 2020. Research progress about nitrogen effects on potato source-sink relationship [J]. Crops, 36(3): 22-26." [郝凱, 賈立國, 秦永林, 等, 2020. 氮素對馬鈴薯源-庫關系影響研究進展 [J]. 作物雜志, 36(3): 22-26.]
JIAN SF, LIAO Q, SONG HX, et al., 2018. NRT1.1-related NH4+ toxicity is associated with a disturbed balance between NH4+ uptake and assimilation" [J]. Plant Physiol, 178(4): 1473-1488.
LIANG J, YE Y, YANG W, 2018. Effects of different nitrogen form ratios on growth and quality of Asparagus cochinchinensis (Lour.) Merr." [J]. Soil Fert Sci China (1): 28-31." [梁娟, 葉漪, 楊偉, 2018. 不同氮素形態及配比對天門冬生長和品質的影響 [J]. 中國土壤與肥料 (1): 28-31.]
LIANG Y, CHEN SS, WEI KH, et al., 2020. Chromosome level genome assembly of Andrographis paniculata" [J]. Front Genet, 11: 701.
LIU T, 2018. Mechanisms underlying the effects of nitrogen supply on leaf nitrogen-forms partitioning and photosynthetic nitrogen use efficiency of Brassica napus" [D]. Wuhan: Huazhong Agricultural University" [劉濤, 2018. 氮供應對油菜葉片氮形態分配與光合氮利用效率的影響機理 [D]. 武漢: 華中農業大學.]
LUO JQ, FU LZ, 2020. Research progress on the effect of nitrogen on the accumulation of active ingredients in medicinal plants" [J]. J Zhejiang Agric Sci, 61(6): 1046-1049." [羅佳琪, 付立忠, 2020. 氮素對藥用植物有效成分累積的影響研究進展 [J]. 浙江農業科學, 61(6): 1046-1049.][JP2]
L WX, GE Y, WU JZ, et al., 2004. Study on the method for the determination of nitric nitrogen, ammoniacal nitrogen and total nitrogen in plant" [J]. Spectrosc Spectr Anal,24(2): 204-206." [呂偉仙, 葛瀅, 吳建之, 等, 2004. 植物中硝態氮、氨態氮、總氮測定方法的比較研究 [J]. 光譜學與光譜分析, 24(2): 204-206.]
MAKINO A, SAKUMA H, SUDO E, et al., 2003. Differences between maize and rice in N-use efficiency for photosynthesis and protein allocation [J]. Plant Cell Physiol, 44(9): 952-956.
MU XH, CHEN QW, CHEN FJ, et al., 2016. Within-leaf nitrogen allocation in adaptation to low nitrogen supply in maize during grain-filling stage" [J]. Front Plant Sci, 7: 699-709.
REN JZ, 2023. Biosynthetic pathway elucidation and microbial cell factory construction of active diterpenoids of Andrographis panucilata" [D]. Shanghai: Naval Medical University." [任俊澤, 2023. 穿心蓮活性二萜的生物合成途徑解析及微生物細胞工廠構建 [D]. 上海: 中國人民解放軍海軍軍醫大學.]
SALONER A, BERNSTEIN N, 2022. Nitrogen source matters: high NH4+/NO3- ratio reduces cannabinoids, terpenoids, and yield in medical cannabis [J]. Front Plant Sci, 13: 830224.
SHI ZM, TANG JC, CHENG RM, et al., 2015. A review of nitrogen allocation in leaves and factors in its effects [J]. Acta Ecol Sin, 35(18): 5909-5919." [史作民, 唐敬超, 程瑞梅, 等, 2015. 植物葉片氮分配及其影響因子研究進展 [J]. 生態學報, 35(18): 5909-5919.]
SUN MY, XU SQ, LI JY, et al., 2020.Transcriptome analysis and discovery of genes related to andrographolide synthesis of Andrographis paniculata under UV-C treatment" [J]. J S Agric, 53(3): 618-627." [孫銘陽, 徐世強, 李靜宇, 等, 2020. UV-C處理穿心蓮轉錄組分析及穿心蓮內酯合成相關基因挖掘" [J]. 南方農業學報, 53(3): 618-627.]
SUN Y, ZHANG T, XU X, et al., 2021. Transcriptomic characterization of nitrate-enhanced stevioside glycoside synthesis in stevia (Stevia rebaudiana) Bertoni [J]. Int J Mol Sci, 22: 8549.
TANG XQ, YANG Y, L TT, et al., 2017. Effects of nitrogen form and ratio on growth and active component content in summer-planted Isatis indigotica" [J]. J Plant Resour Environ, 26(1): 21-29." [唐曉清, 楊月, 呂婷婷, 等, 2017. 氮素形態及配比對夏播菘藍生長及活性成分含量的影響 [J]. 植物資源與環境學報, 26(1): 21-29.]
WANG YX, ZOU XZ, LI CY, et al., 2023. Investigation of yield traits and medicinal components of Andrographis paniculata germplasm" [J]. Acta Sci Nat Univ Sunyat, 62(3): 100-108." [王雨霞, 鄒秀崽, 李春雨, 等, 2023. 穿心蓮種質產量性狀與藥用成分分析 [J]. 中山大學學報(自然科學版), 62(3): 100-108.]
XU J, WANG XY, GUO WZ, 2015. The cytochrome P450 superfamily: Key players in plant development and defense" [J]. J Integr Agric, 14(9): 1673-1686.
YAN FX, WANG KC, TANG XQ, et al., 2010. Effects of NH4+-N/NO3--N ratio on the active component and mineral elements contents of Isatis indigotica Fort." [J]. Jiangsu J Agric Sci, 26(4): 735-739." [晏楓霞, 王康才, 唐曉清, 等, 2010. 氮素形態對菘藍活性成分和礦質元素含量的影響 [J]. 江蘇農業學報, 26(4): 735-739.]
YANG YY, WANG F, WAN Q, et al., 2018. Transcriptome analysis using RNA-Seq revealed the effects of nitrogen form on major secondary metabolite biosynthesis in tea (Camellia sinensis) plants [J]. Acta Physiol Plant, 40: 127.
ZHANG Q, LIU Y, WU XM, et al., 2018. Preliminary study on the effects of two different nitrogen forms on flavonoids accumulation and related molecular mechanism in medicinal plant Tetrastigma hemsleyanum" [J]. Bull Bot Res, 38(3): 367-376." [張強, 劉一, 吳曉敏, 等, 2018. 兩種氮素形態對藥用植物三葉青中黃酮類活性成分積累的影響及相關分子機制的初步探討 [J]. 植物研究, 38(3): 367-376.]
ZHANG XZ, 1986. Determination of chlorophyll content in plants — Acetone ethanol mixture method" [J]. Liaoning Agric Sci (3): 26-28." [張憲政, 1986. 植物葉綠素含量測定——丙酮乙醇混合液法 [J]. 遼寧農業科學 (3): 26-28.]
ZHONG C, JIAN SF, CHEN DL, et al., 2021. Organic nitrogen sources promote andrographolide biosynthesis by reducing nitrogen metabolism and increasing carbon accumulation in Andrographis paniculata" [J]. Plant Physiol Biochem, 164: 82-91.
