水分和土壤微生物對干熱河谷山合歡和銀合歡生長性狀的影響
2024-12-31 00:00:00石磊琦劉梓鈺王超俊王垠昊杜訓秋王雪梅
廣西植物 2024年11期



摘 要: "植被恢復是干熱河谷自然植被恢復的有效途徑,干熱河谷自然植被修復的同時也引進了很多外來種。為比較本地種和外來引進種對水分與微生物的響應差異,該文選取當地鄉土種山合歡(Albizia kalkora)和外來種銀合歡(Leucaena leucocephala),采用氣候箱模擬云南干熱河谷地區條件,通過土壤水分控制和微生物接種研究了水分和各自的土壤微生物對兩種豆科植物生長發育的影響。結果表明:(1)山合歡的種子發芽率、根生物量、根冠比和植株氮磷比較銀合歡高58.82%、76.13%、90.59%和18.42%(Plt;0.05)。(2)干旱條件下山合歡的根生物量和植株氮含量比濕潤條件低33.67%和8.65%(Plt;0.05),銀合歡的株高、地上生物量、根生物量在干旱條件下較濕潤條件低39.32%、40.06%和39.61%(Plt;0.05),表明干旱抑制了兩種植物生長和養分吸收。(3)山合歡在干旱條件下的根瘤數量多于濕潤條件(Plt;0.05),而銀合歡在濕潤條件與干旱條件下的根瘤數量沒有顯著性差異。(4)除枯葉比例外,土壤微生物接種、土壤微生物與水分的交互作用對山合歡和銀合歡的生長并沒有造成顯著影響。以上研究結果表明山合歡較銀合歡可能更適宜干旱環境,為干熱河谷植被恢復物種選擇提供了一定的理論依據,但土壤微生物對植物生長的影響還值得進一步探索和研究。
關鍵詞: 干熱河谷, 山合歡, 銀合歡, 土壤水分, 土壤微生物
中圖分類號: "Q945
文獻標識碼: "A
文章編號: "1000-3142(2024)11-2125-16
Effects of water and soil microorganisms on the growth
traits of Albizia kalkora and Leucaena
leucocephala in a dry-hot valley
Abstract: "Vegetation restoration is an effective way for ecological restoration in the dry-hot valley, and many alien species are introduced along with natural vegetation restoration. In order to compare the responses of native and alien species to water and microorganisms, the native Albizia kalkora and alien Leucaena leucocephala were selected as our research objects, and the effects of water and soil microorganisms on the growth and development of the two plants were studied by soil water control and soil microbial inoculation under climate chamber conditions. The results were as follows: (1) Compared with Leucaena leucocephala, the seed germination rate, root biomass, root-shoot ratio and plant N∶P ratio of Albizia kalkora were significantly increased by 58.82%, 76.13%, 90.59% and 18.42%, respectively (Plt;0.05). (2) Under drought conditions, the root biomass and plant nitrogen content of Albizia kalkora were significantly lower than those under moist conditions by 33.67% and 8.65% (Plt;0.05), and the plant height, aboveground biomass and root biomass of Leucaena leucocephala under drought conditions were 39.32%, 40.06% and 39.61% lower than those under moist conditions (Plt;0.05), indicating that drought inhibited the growth and nutrient uptake of both plants. (3) The quantity of root nodules in Albizia kalkora under drought conditions was higher than that under moist conditions (Plt;0.05), and the quantity of root nodules in "Leucaena leucocephala under moist conditions was higher than that under drought conditions, but no significant difference. (4) Except for the litter proportion, soil microbial inoculation and the interaction between soil microorganisms and water had no significant effect on the growth of Albizia kalkora and Leucaena leucocephala. The above results suggest that Albizia kalkora is more suitable for drought environment than Leucaena leucocephala, which provides a theoretical basis for the selection of species for vegetation restoration in dry-hot valley, but the effects of soil microorganisms on plant growth are worthy of further exploration and research.
Key words: dry-hot valley, Albizia kalkora, Leucaena leucocephala, soil water, soil microorganisms
云南元謀干熱河谷鑲嵌在云南高原亞熱帶濕潤與半濕潤區中的高熱多光照干旱河谷區,地處滇中高原北部的金沙江一級支流——龍川江下游(趙琳等,2006)。該區全年高溫、多光照且干旱,晝間日照時長接近16 h,年平均氣溫不低于20 ℃,為各種植物的生長提供了充足的熱量條件。但與此同時,該地區環境與氣候條件惡劣,表現在氣候干旱、土壤貧瘠,植被覆蓋率低,水土流失嚴重,生態環境極其脆弱。植被恢復是干熱河谷區生態恢復的有效途徑,1992年至今,眾多學者從物種篩選和恢復技術兩方面開展了大量研究(楊振寅等,2007)。在物種選擇上,主要從植物的耐旱性出發對物種進行篩選,并引進了很多外來物種,如桉樹、銀合歡、辣木、印楝、相思樹等;在恢復技術上,主要從整地、育苗、抗旱保水技術等方面進行研究(周麗麗等,2021;趙保榮和楊春鳳,2023)。干旱是干熱河谷首要的環境脅迫因子,土壤水分條件的改善是干熱河谷退化生存系統恢復與重建的關鍵問題(鐘祥浩,2000)。同時,該區土壤類型主要為燥紅土,具有缺氮、少磷、貧有機質的特點(張燕平等,2005)。在這種貧瘠的環境條件下,植物與土壤微生物間的關系也顯得十分緊密(Sardans amp; Peuelas, 2013)。目前,關于水分脅迫對干熱河谷植物生理生長特征的影響已有較多研究,但該區土壤微生物對植物生長的影響還鮮有報道。
土壤中的水分含量會對植物的生長發育產生直接影響。水分是植物體的重要組成部分,植物的生長一般情況下是依靠吸收土壤中的水來伸長或膨大細胞來實現,膨壓降低,即土壤水分過低時,植物的生長發育就會減緩甚至停止(雷麗等,2009)。同時,土壤含水量也會通過影響土壤肥力來影響植物的生長發育(李連智和韓琳,2019)。因此,對植物抗旱性的研究應運而生,不同植物適應干旱環境的能力一定程度上可以通過測定植物的生長性狀來實現(Kudoyarova et al., 2013)。研究也證明了在干旱脅迫下,植物的地上生物量和地下生物量均會有所下降,但地上生物量降低程度要明顯高于地下生物量的降低程度(Haffani et al., 2014)。土壤微生物是土壤活有機體的重要組成部分,與植物生長、養分吸收等密切相關,其種類、數量、活性都會影響植物的生長發育。土壤微生物對植物的影響分為直接影響和間接影響。其中,直接影響包括菌根真菌對植物的作用、固氮共生體對植物的影響、病原菌對植物的影響等;而土壤微生物對植物的間接影響是通過游離微生物來實現(王雪梅等,2017)。同時,土壤微生物對植物的影響也可分為正和負兩方面,正效應通常是因為微生物群落中含有對植物生長的有益菌,如根瘤菌的固氮作用、腐生微生物對土壤中有機質分解產生利于植物生長的無機營養;而負效應則主要是由于病原菌所引起。Lau和Lennon(2012)研究表明,植物對干旱的適應性響應主要取決于其土壤微生物的響應,暗示了植物面臨環境脅迫不只限于適應或遷移途徑,也可以得益于其周圍生物群落的快速響應。但是,關于土壤微生物與水分對植物生長的交互作用并不十分清楚。
山合歡(Albizia kalkora)為豆科、含羞草亞科、合歡屬植物,生長于溪溝邊、路旁和山坡上,是干熱河谷地區常見的野生鄉土種,也是云南省中部高原荒山河谷造林先鋒樹種(崔永忠等,2010)。銀合歡(Leucaena leucocephala)為豆科、含羞草亞科、銀合歡屬植物,原產于美洲熱帶地區,自20世紀60年代以來,我國開始大規模引種銀合歡。因為銀合歡具有較強的環境適應性、對水分要求低等特點,所以被列為干熱河谷地區造林的優良樹種。同時研究表明,銀合歡具有高蒸騰、低水分利用效率等特征,具有潛在的入侵植物危害屬性,因此也被列為入侵植物(趙廣等,2019)。段愛國等(2013)分別對山合歡和銀合歡的水分利用效率進行了動態分析,從生長情況上來看,山合歡的地徑略大于銀合歡,但株高和冠幅均小于銀合歡,從不同條件下植株的水分利用效率上來看,在干旱季,山合歡的水分利用效率要大于銀合歡,而在濕潤季,山合歡的水分利用效率要小于銀合歡,兩者對干旱脅迫的響應特征尚不明晰。此外,豆科植物與土壤微生物關系十分密切,可形成叢枝菌根真菌(arbuscular mycorrhizal fungi, AMF)-豆科植物-根瘤菌三重共生體,是植物與微生物共生中的一種特殊類型(Igiehon amp; Babalola, 2018),對土壤微生物變化十分敏感(Yang et al., 2020)。目前,山合歡和銀合歡對土壤微生物及其與水分交互作用的響應也沒有相關研究。
基于此,本文以干熱河谷本地豆科山合歡和外來豆科銀合歡為研究對象,依托當地特有的燥紅土為試驗土壤,采用室內控制和氣候箱培養方法,通過研究水分、土壤微生物及其交互作用對山合歡和銀合歡生長的影響,擬探討以下問題:(1)山合歡和銀合歡對水分脅迫的響應是否存在差異;(2)土壤微生物是否影響植物生長性狀并影響植物對水分脅迫的響應。本研究的山合歡和銀合歡均為干熱河谷優勢豆科植物,形態特征十分相似,但山合歡為本地種,銀合歡為外來種,研究材料具有對比性的特色,可在植被恢復物種選擇方面提供理論依據;同時,研究過程中不僅關注水分對植物生長的影響,還關注土壤微生物對植物生長的作用,擬從植被恢復技術上為該區土壤微生物功能的調節和利用提供科學支撐。
1 材料與方法
1.1 試驗材料
1.1.1 植物種子 本試驗采用生長于干熱河谷地區山合歡和銀合歡植物的種子,種子于成熟季節從干熱河谷地區采集,并經風干后保存于布袋中。山合歡種子的百粒重為(8.689±0.003) g,銀合歡種子的百粒重為(5.460±0.016) g。挑選飽滿、大小基本一致的種子進行試驗。
1.1.2 盆栽土壤 試驗用土選自野外山合歡和銀合歡生長區的燥紅土。經野外調研,在元謀干熱河谷典型區域(101°48′36″ E、25°40′12″ N)進行土壤樣品采集,此處同時生長著山合歡和銀合歡植株,環境差異較小。采樣時,分別選擇5~10個山合歡植株采樣點和銀合歡植株采樣點,各樣點山合歡和銀合歡植株大小基本一致,用鏟子和鋤頭在緊鄰植株主干周圍按十字交叉法挖取4個方位土壤約 50 cm 深。為消除土壤中本身存在的微生物,將采集的山合歡和銀合歡土壤樣品分別置于高溫高壓滅菌鍋中濕熱滅菌(121℃、103.5 kPa)1 h,并重復滅菌3次,滅菌間隔時間24 h。高溫高壓滅菌是土壤滅菌的有效方法,并且對土壤理化性質的影響較小(Berns et al., 2008)。鄭嘉慧等(2017)研究也表明采用高溫高壓蒸汽法對土壤滅菌3次,每次間隔24 h可以達到很好的滅菌效果。滅菌后,為了控制非試驗因素的一致性,使所有處理的土壤理化性質一致,并避免山合歡和銀合歡對土壤的適應偏好,試驗以滅菌混合后的山合歡土壤和銀合歡土壤為基質,即取等質量的山合歡滅菌土和銀合歡滅菌土混合裝入滅菌的盆缽中(121 ℃、20 min),作為基質土壤。基質土的基本理化性質如下:pH為(6.20±0.01),有機碳含量為(1.55±0.47) g·kg-1,全氮含量為(0.43±0.05) g·kg-1,全磷含量為(0.01±0.00) g·kg-1,全鉀含量為(0.58±0.02)" g·kg-1,有效氮含量為(15.16±3.31) mg·kg-1,有效磷含量為(0.34±0.02) mg·kg-1,速效鉀含量為(57.26±2.29) mg·kg-1。
1.1.3 微生物接種用土 2022年1月5日,采集已經分別種植了80 d山合歡和80 d銀合歡的新鮮土壤用作微生物接種用土,經高通量測序,山合歡和銀合歡土壤的微生物群落組成和多樣性如表1所示。將采集的新鮮土壤保存于-20 ℃冰箱中備用。
1.2 試驗設計
整個試驗包括植物、水分、土壤微生物3個因素。其中,植物分為山合歡和銀合歡。水分處理采用傳統的稱重法進行控制,通過控制不同的土壤田間持水量(field capacity, FC)水平設置水分梯度(Wang et al., 2016;劉愛林等,2023)。試驗前測出該區燥紅土的FC為12%,據此設置濕潤(W)和干旱(D)兩個梯度,濕潤為80%~85% FC,干旱為40%~45% FC,對應的土壤質量含水量分別為9.6%~10.2%和4.8%~5.4%。微生物處理分為未接種(M0)和接種土壤微生物(M)兩種。由于不同植物下的土壤微生物群落具有差異,對本地種和外來種的反饋效應也不同,本研究主要關注植物生長過程中自身的土壤微生物對其生長發育的反饋作用,因此山合歡處理接種山合歡土壤微生物,銀合歡處理接種銀合歡土壤微生物,最終形成山合歡(DM0、WM0 、DM 、WM)和銀合歡(DM0、WM0 、DM、WM)共8種處理,每個處理設置4個重復。
本研究采用室內培養法,利用氣候箱模擬干熱河谷的溫濕度條件,未考慮當地地形的影響。試驗于2022年2月底開始,試驗時,先稱量空盆缽重量,將盆缽滅菌(121 ℃,20 min)后根據盆缽體積取等質量的山合歡滅菌土(400 g)和銀合歡滅菌土(400 g)混勻后裝入滅菌的盆缽中,用作盆栽基土,同時澆適量滅菌水保持土壤濕潤。挑選健康、飽滿的植物種子,經過濃硫酸浸泡攪拌5 min后破除休眠,用無菌水反復沖洗干凈后播種于盆缽中,每盆播10顆銀合歡或山合歡種子,播種后再在表面覆蓋等質量的滅菌基質,放入氣候培養箱(晝夜:16 h/8 h,35 ℃/25 ℃,光照度15 000 lx)中培養。35 ℃/25 ℃的晝夜溫度接近于干熱河谷區植物生長季的空氣溫度(王雪梅等,2017)。前期保持等量足夠的澆水量促使種子發芽,待所有盆缽均出現4株幼苗時間苗,保持每盆有4株幼苗。繼續培養一段時間后,開始進行微生物接種處理。
土壤微生物接種參照van de Voorde等(2012)的方法,以接種土壤微生物懸浮液的方式進行,具體方法如下:分別以山合歡和銀合歡的新鮮土壤為接種源,取質量為土壤基質10%的新鮮土壤(即80 g新鮮土),按1∶1的土水比加入滅菌水中,混合并攪拌2 min。先將懸浮液放置15 min后,再攪拌2 min并靜置15 min,最后篩分上清液,將上清液過0.5 mm 篩,過濾液即為接種液,并用滅菌水沖洗至相同的體積,為一個盆缽接種量。對照處理添加等量的滅菌后的土壤懸浮液。微生物接種1個月后開始進行水分脅迫,通過稱重法控制不同的田間持水量進行澆水。每天進行稱重并補充滅菌水以維持含水量,并記錄各盆缽每次的澆水量。同時,及時收集落葉,觀察植物變化。水分脅迫72 d后收獲植物和土壤樣品進行指標測定。
1.3 指標測定方法
(1)植物生長和生物量指標:用直尺測量植株株高;將各盆中植株地上部分和根系部分分開取樣,將根系和地上部分于65 ℃烘干至恒重后稱量每盆中根系生物量和地上生物量。同時,將試驗過程中收集的枯葉于65 ℃烘干稱重。每盆隨機選擇 20 片長勢良好的葉片,首先將采集的葉片迅速稱鮮重(WF),然后在5 ℃的黑暗環境中用去離子水浸泡 12 h 后用吸水紙吸去葉片表面水分,迅速稱其飽和鮮重(WR),最后將葉片放入 65 ℃烘箱中烘干至恒重后稱干重(WD),計算相對含水量(%)= [(WF-WD)/(WR-WD)] ×100。
(2)植物養分指標:將植株地上部分烘干后磨細,用于測定植株氮磷含量。運用凱氏定氮法測量植物氮含量,釩鉬黃吸光光度法測量磷含量。
(3)土壤指標:每盆中取一定量的、同樣深度的土壤,烘干稱量土壤重量含水量。通過稀釋平板法測定土壤中細菌、霉菌、放線菌數量,細菌采用牛肉膏瓊脂培養基,濃度梯度為10-4,放線菌采用高氏一號培養基,濃度梯度為10-2,霉菌采用孟加拉紅固體培養基,濃度梯度為10-2。土壤有效氮含量采用堿解擴散法,土壤有效磷含量采用鉬銻抗比色法。
(4)共生體指標:根系取樣時通過計數法統計根瘤數量。采用醋酸墨水染色法(楊亞寧等,2010)在高倍顯微鏡下觀察菌絲、泡囊和叢枝的交叉點數,最后計算叢枝菌根真菌的侵染情況。
1.4 數據分析
本試驗采用Excel和SPSS 22.0軟件對數據進行統計分析。首先,用獨立樣本T檢驗分析山合歡和銀合歡是否存在顯著性差異。然后, 由于接種微生物時接種的是各自的土壤微生物,需要進行分開比較,所以要對山合歡和銀合歡分開處理。最后,分別采用雙因素方差分析法檢驗水分和土壤微生物及其相互作用對山合歡和銀合歡的影響。通過多重比較檢驗不同處理下各指標的差異顯著性,各個處理之間的差異性用Turkey HSD檢驗法進行多重比較,顯著性水平為0.05。
2 結果與分析
2.1 水分控制情況與植物水分特征
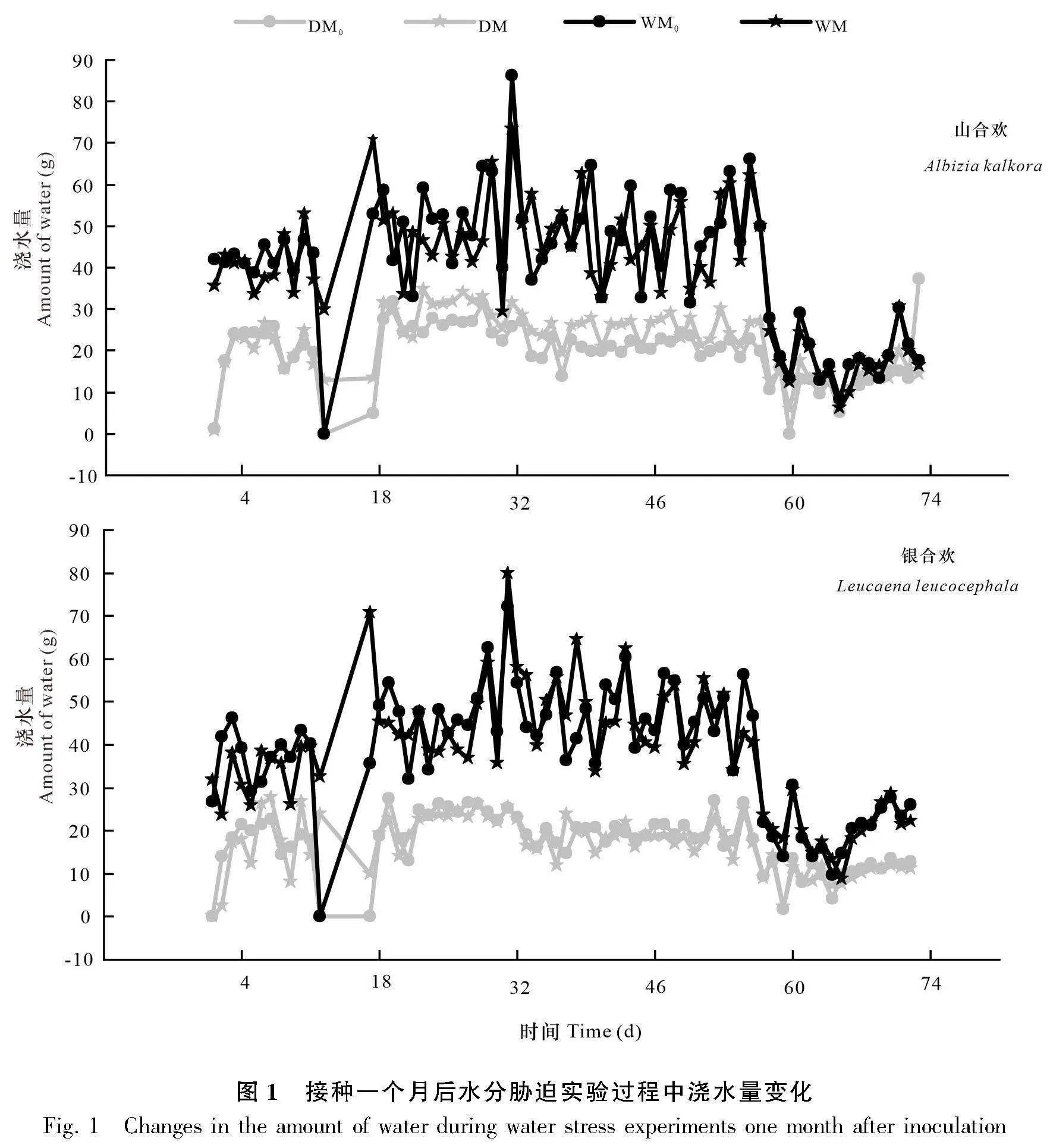
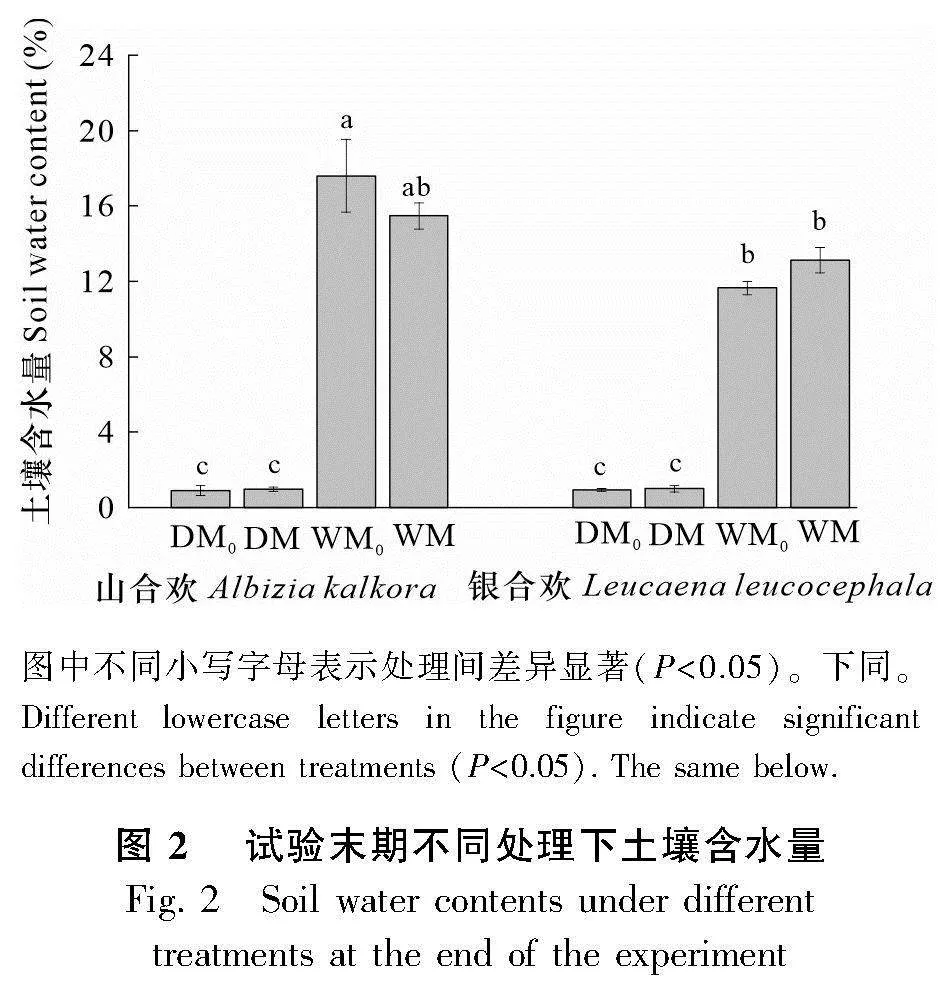
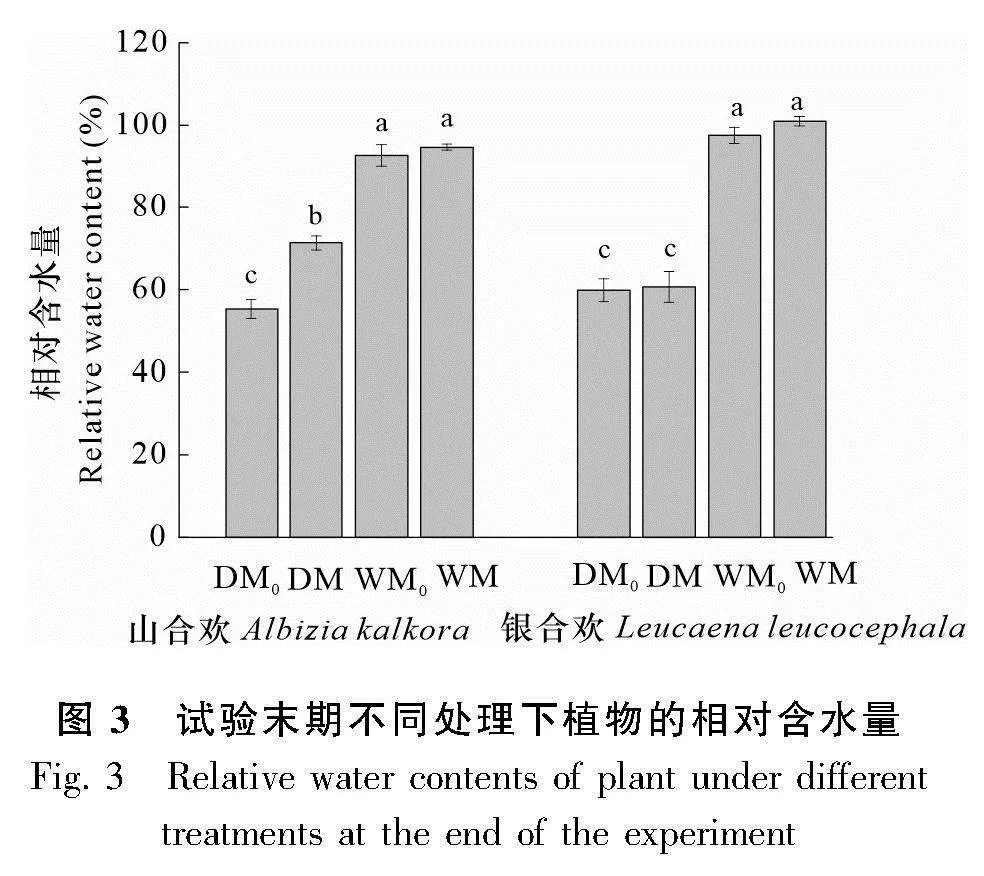
經過試驗設計,最終確定了干旱(DM0、DM)與濕潤(WM0、WM)兩種水分情況,并且以重量含水量的方式來控制干旱與濕潤的情況。每天都通過重量來控制水分使其處于干旱或者濕潤的條件,具體澆水量結果如圖1所示,干旱和濕潤處理在澆水量上有著明顯差異,這說明干旱和濕潤處理界限明確。在試驗末期(即水分處理72 d后收獲樣品期間),我們對干旱與濕潤處理條件下的土壤進行了土壤含水量的測定(圖2)。通過圖2也可以看出干旱與濕潤處理的界限明確,干旱組土壤含水量平均值為0.95%,濕潤組土壤含水量平均值為14.46%,存在顯著差異,表明干旱和濕潤處理的土壤水分梯度明顯。由圖3可知,在經過水分處理后,試驗末期干旱與濕潤處理下植物的相對含水量具有明顯的梯度差異,濕潤條件下植物的相對含水量比干旱條件下的相對含水量高了56.00%。
2.2 植物生長特征
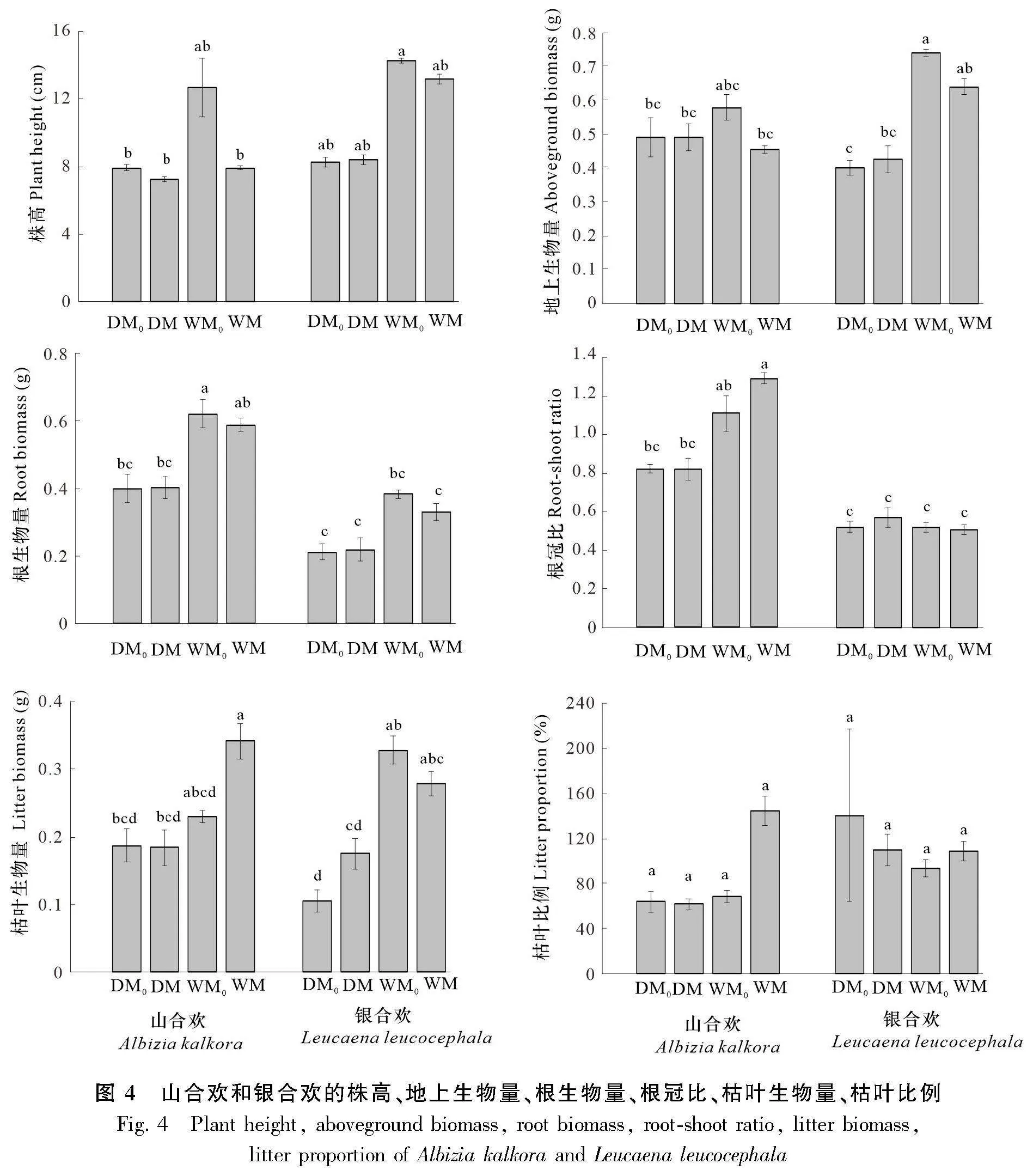
結合統計分析結果(表2)與山合歡和銀合歡各項指標(圖4)可以看出,山合歡和銀合歡在發芽率、根生物量、根冠比上有顯著性差異(Plt;0.01)。山合歡的發芽率為54.76%,銀合歡的發芽率為34.48%;山合歡的根生物量和根冠比分別比銀合歡高76.13%和90.59%。就山合歡而言,不同的水分條件對其根生物量、根冠比有顯著性影響(Plt;0.01),對枯葉生物量、枯葉比例也有影響(Plt;0.05)。濕潤條件下山合歡的根生物量、根冠比、枯葉生物量和枯葉比例分別比干旱條件高50.76%、46.25%、54.13%和70.27%。就銀合歡而言,不同的水分條件對株高、地上生物量、根生物量、枯葉生物量也有顯著性影響(Plt;0.01),濕潤條件下,銀合歡的株高、地上生物量、根生物量和枯葉生物量分別比干旱條件高64.79%、66.83%、65.59%和117.27%。從微生物處理方面來看,其并未對山合歡與銀合歡各項指標產生顯著性影響,但微生物及水分與微生物的交互作用對山合歡的枯葉比例有影響(Plt;0.05),濕潤條件下接種微生物處理顯著增加了山合歡的枯葉比例。
2.3 不同處理下的植物養分特征
不同處理情況下植物體地上部的氮含量、磷含量和氮磷比結果如圖5所示。結合表2與圖5分析可知,山合歡和銀合歡的磷含量和氮磷比有顯著性差異(Plt;0.05)。山合歡磷含量為0.487 2 g·kg-1,銀合歡磷含量為0.590 3 g·kg-1且比山合歡高21.2%;山合歡的氮磷比(45)要顯著高于銀合歡(38)。水分對山合歡的氮含量具有顯著性影響(Plt;0.05),濕潤條件下山合歡的氮含量(23.13 g·kg-1)比干旱條件下的氮含量(21.13 g·kg-1)高9.47%。同樣,微生物及微生物與水分的交互作用對氮磷養分指標沒有造成顯著影響(Pgt;0.05)。
2.4 共生體特征
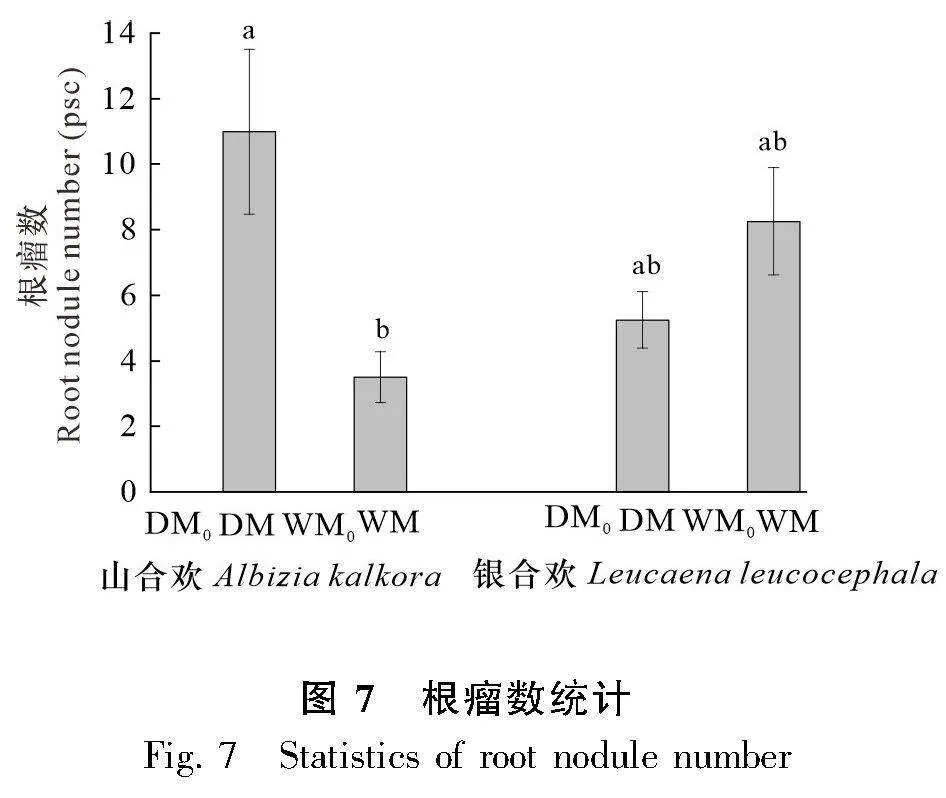
在收集處理植物根系的過程中,通過統計微生物處理條件下的根瘤數量(圖6),發現在接種微生物的條件下,山合歡在干旱條件下的根瘤數目(11)比濕潤條件下的根瘤數目(3.5)多(Plt;0.05),而銀合歡在濕潤與干旱條件下的根瘤數量沒有顯著性差異(圖7)。
此外,使用顯微鏡對植物根系進行觀察,發現只有個別樣品中存在共生微生物AMF侵染(圖8),絕大部分山合歡和銀合歡根系中未觀察到AMF侵染。
2.5 不同處理下的土壤特征
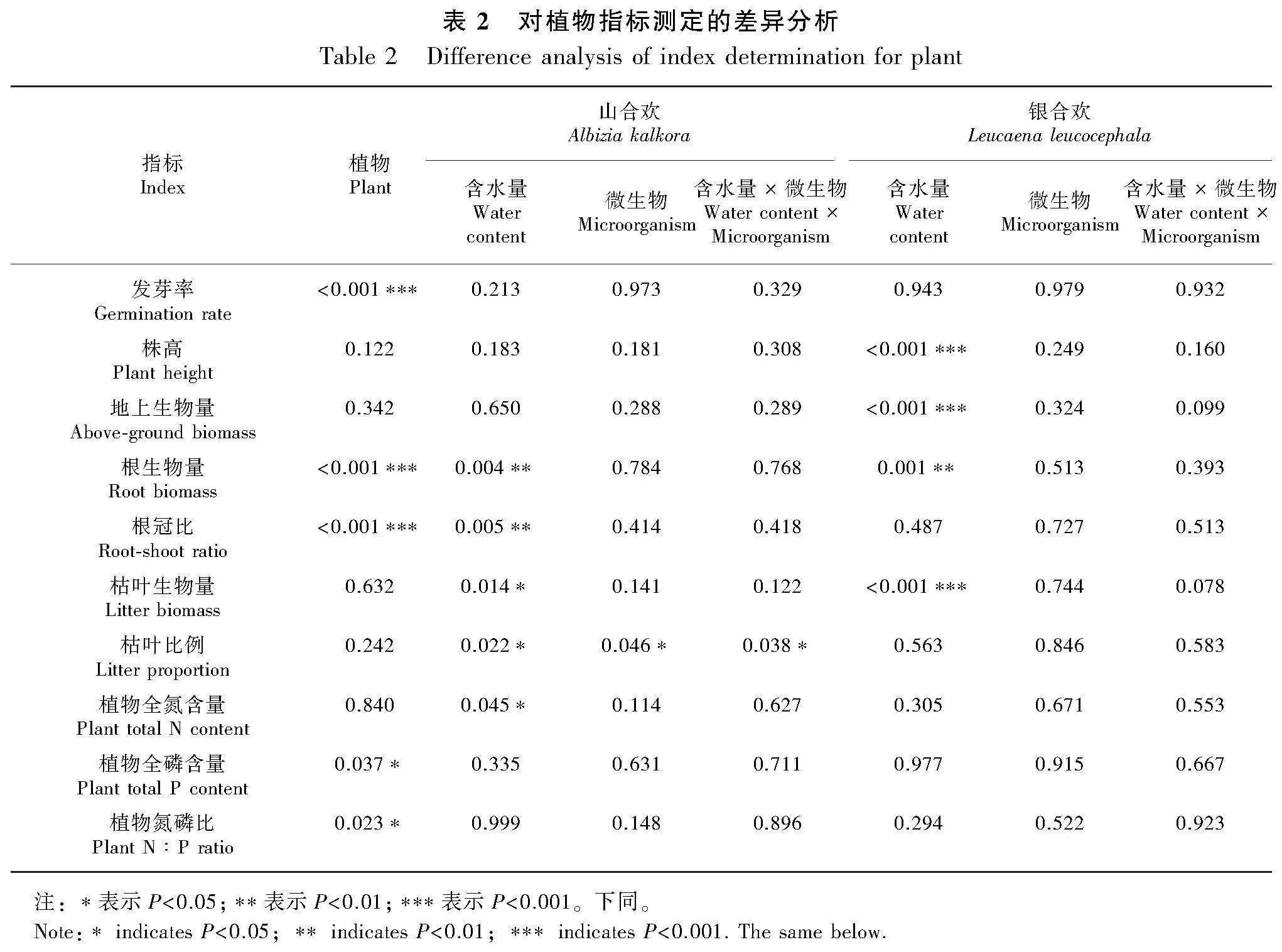
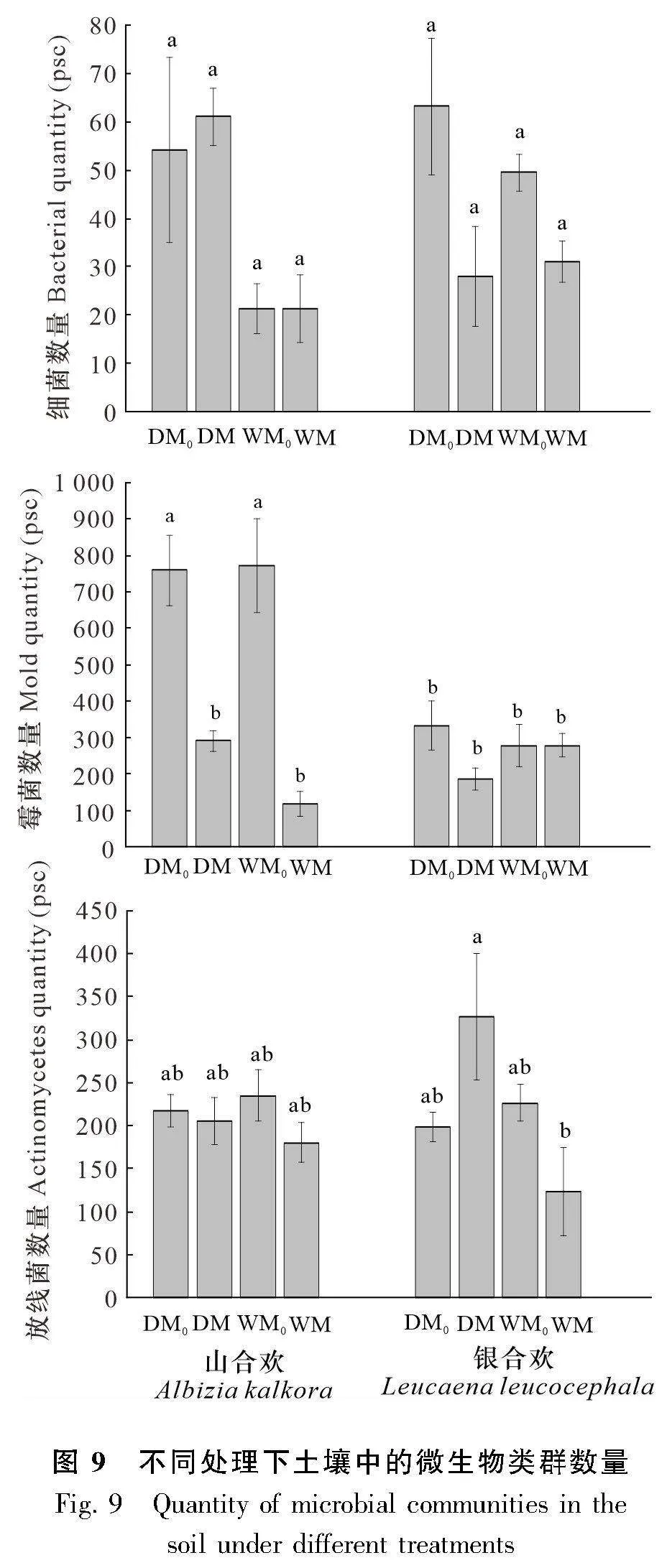
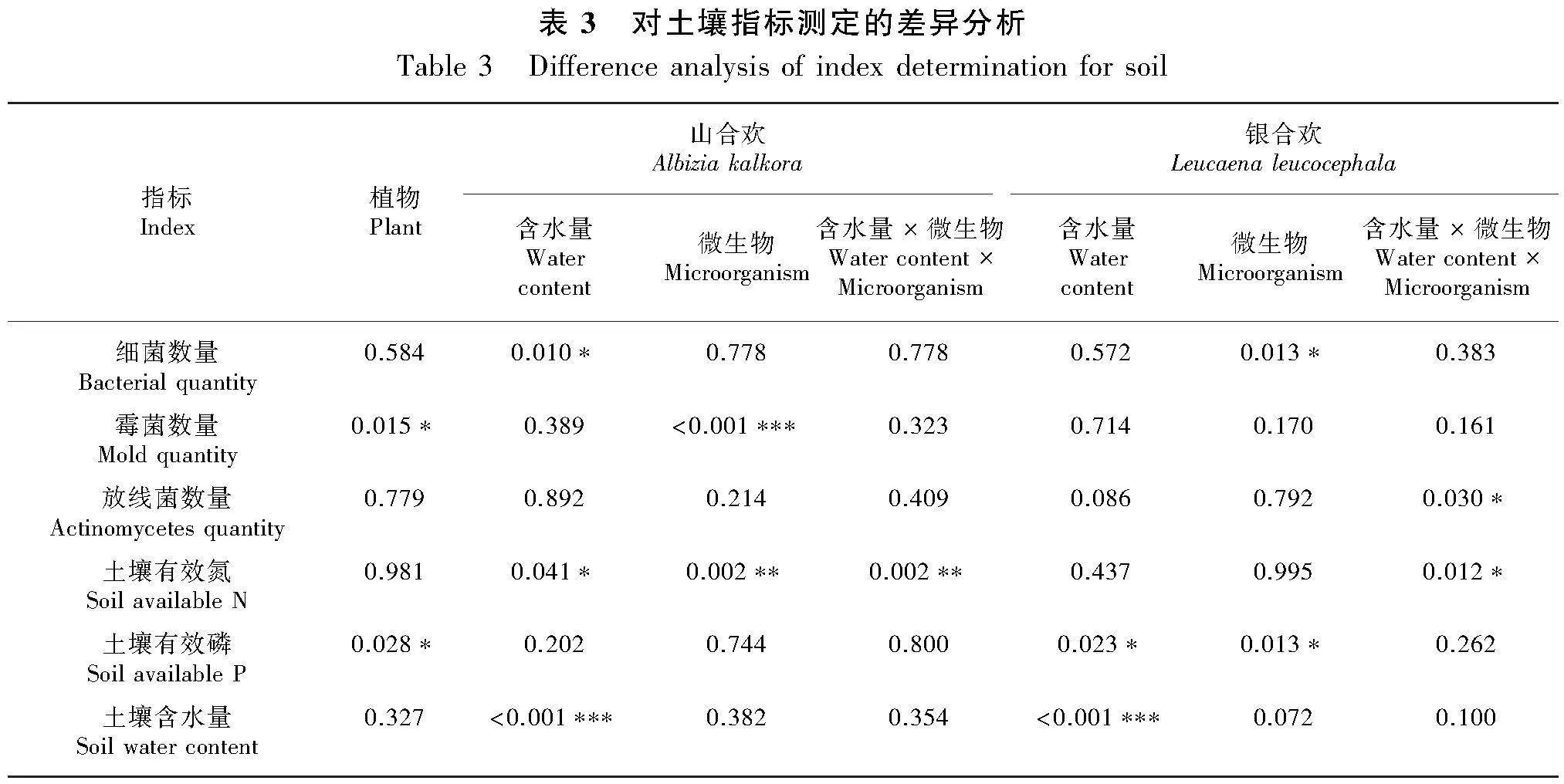
結合方差分析結果的顯著性P值表(表3)與分析土壤微生物類群的柱狀圖(圖9)可以看出,培養不同植物的土壤在霉菌數量上存在差異性(Plt;0.05 ),表現為種植山合歡的土壤霉菌數顯著高于銀合歡80.81%。就山合歡而言,水分對其細菌數量有顯著性影響(Plt;0.01),干旱條件下的土壤細菌數比濕潤條件多171.18%;微生物處理對其霉菌數量有顯著性影響(Plt;0.01),不接種微生物的處理下土壤霉菌數比接種微生物的處理高272.84%。就銀合歡而言,微生物處理對其細菌數量有影響(Plt;0.05),不接種微生物的處理其土壤中的細菌比接種微生物的處理高91.10%。
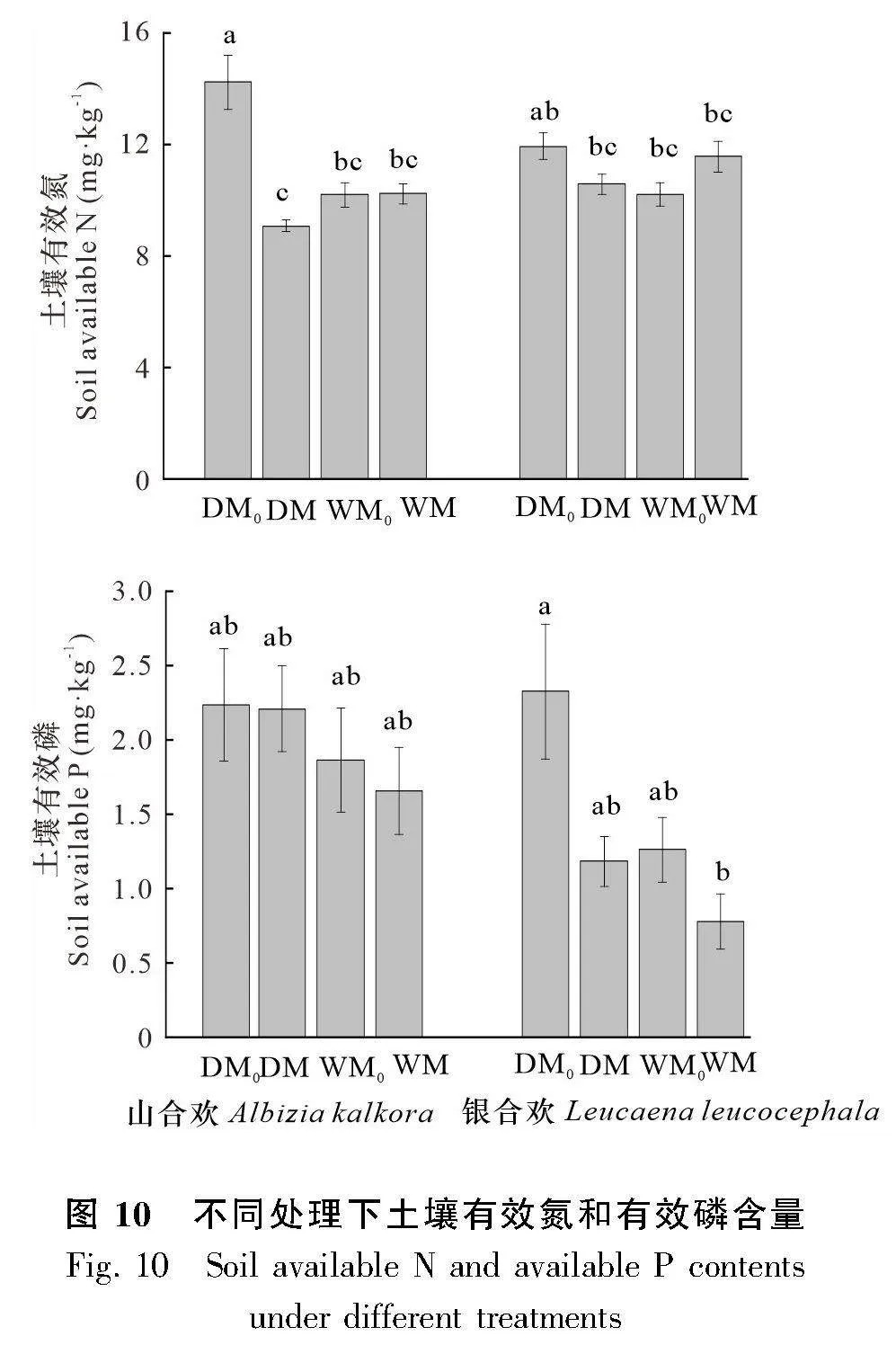
由表3與分析土壤養分的柱狀圖(圖10)結果可知,兩種植物的土壤有效磷含量有顯著性差異(Plt;0.05),山合歡的土壤有效磷含量比銀合歡顯著高43.50%。就山合歡而言,水分與微生物對土壤有效氮含量有顯著的交互作用(Plt;0.01),干旱條件下不接種處理的土壤有效氮含量顯著高于其他處理。就銀合歡而言,水分條件和微生物處理均對土壤有效磷含量有顯著影響(Plt;0.05),干旱條件下土壤有效磷含量比濕潤條件下高72.04%,而接種微生物處理的土壤有效磷含量比不接種微生物處理低45.32%。與山合歡一致,水分與微生物對銀合歡的土壤有效氮含量具有顯著的交互作用 (Plt;0.05)。此外,和種植前的培養基土相比,種植山合歡和銀合歡后,土壤的有效氮含量降低了26.98%,但土壤有效磷含量提高了391.18%。
3 討論
3.1 水分對植物的影響
根據本研究數據可知,濕潤條件下的植物體在株高、生物量、植物氮濃度等方面的表現要強于干旱條件下的植物體,說明干旱抑制了植物生長發育。研究表明,金沙江干熱河谷區旱季土壤含水率常降至5%以下(魏漢功和葉厚源,1991),本研究中干旱水平的土壤含水量基本達到旱季的土壤含水量水平,濕潤與干旱處理的含水量與銀合歡林地雨季和旱季的土壤含水量也比較接近(韓姣姣等,2019)。水分條件對本地種與外來入侵種的影響,也就是本地種與外來入侵種在抗旱性上的差異,已有部分學者做了相關研究。李玲等(2008)的研究發現,外來入侵種金鐘藤(Merremia boisiana)在入侵地對不同水分環境的生理適應較本地早期入侵種葛藤(Pueraria lobata)占優勢。陳明林和王友寶(2008)的研究發現,本地種酢漿草(Oxalis corniculata)比外來種銅錘草(O. corymbosa)對水分脅迫更為敏感。周蛟和張明友(1998)通過比較銀合歡、念珠相思(Acacia torulosa)、黃豆樹(Albizia procera)、加勒比松(Pinus caribaea)、山合歡5種抗旱耐熱樹種,認為外來種銀合歡、黃豆樹和加勒比松是適合元謀干熱河谷的造林樹種。這與本研究結論不同,本研究通過對山合歡和銀合歡的對比分析發現,本地種山合歡的發芽率、根生物量、根冠比都要高于外來種銀合歡,在干旱條件下山合歡的根瘤數也更多。植物將更多的生物量投資于地下有助于滿足植物本身對養分和水分的強烈需求,這也是植物在水分虧缺時的重要生存對策之一,有助于植物抗旱(Zhang et al., 2004;Zhou et al., 2014)。本研究山合歡根生物量和根冠比大于銀合歡,從這個方面來說,本地種山合歡相比于外來種銀合歡可能更適應干旱脅迫環境。此外,水分是影響植物吸收礦質元素的最活躍因素,礦質營養問題往往成為干旱脅迫的次生效應(da Silva et al., 2011)。本研究干旱條件下山合歡的根瘤數比銀合歡更多,較多的根瘤有利于固定更多的氮,這對緩解當地的氮限制是有利的。段愛國等(2013)研究發現山合歡在干旱季的水分利用效率要大于銀合歡,從這個角度來講也是本地種山合歡更適宜當地干旱環境,也更適合用于當地植被恢復。楊濟達等(2016)提出干熱河谷地區已經成為外來植物入侵的重災區;干熱河谷植被恢復應該以本地樹種為主(楊振寅等,2007)。趙廣等(2019)研究認為,考慮到干熱河谷區銀合歡具有高蒸騰、低水分利用效率特征以及其潛在的入侵植物危害屬性,今后干熱河谷植被恢復物種應謹慎選擇銀合歡。本研究結果與之相符,在干熱河谷植被恢復過程中我們更建議使用本地種山合歡。
3.2 土壤微生物對植物的影響
經微生物處理后土壤微生物對土壤養分及植物的生物量、土壤中的微生物數量的影響可能是通過以下兩種方式:(1)接種的土壤微生物中含有能對植物養分吸收產生促進作用的細菌,如根瘤菌。(2)土壤微生物既可以通過改良土壤養分的有效性提高植物對養分的吸收來促進植物的生長,也可以通過生物化學作用使土壤中的養分流失或揮發到大氣中,從而在一定程度上抑制植物的生長。沈仁芳和趙學強(2015)研究認為,有些土壤微生物能夠與植物根系形成共生關系,或者分布在植物根際,影響植物獲得土壤養分的能力。這類微生物主要有共生固氮菌、叢枝菌根真菌、植物根際促生菌等。畢銀麗等(2014)的研究也表明接種微生物顯著提高了植物的生物量以及植物體內氮、磷、鉀等營養元素的含量,促進了植物的生長,同時提高了植物對土壤礦質元素尤其是磷元素的吸收和利用率。此外,土壤微生物對本地種和外來種的影響具有差異性。一般而言,相比于滅菌(不接種)處理,當地的土壤微生物群落對外來種具有顯著的正效應,而對本地種具有負效應。張令等(2012)的研究結果表明,土壤微生物對入侵種烏桕(Sapium sebiferum)具有正效應;與本地種相比,入侵種烏桕幼苗總生物量、相對生長速度、根冠比以及總葉面積、葉面積比、比葉面積等生物學指標均在新鮮土壤中表現出明顯優勢。梁作盼等(2016)的研究結果顯示,土壤微生物對紫莖澤蘭(Ageratina adenophora)與本地植物香茶菜(Rbdosia amethystoides)的生長均具有顯著的正反饋效應,并且土壤微生物有助于增強紫莖澤蘭對本地植物的競爭優勢。然而,本研究并未出現類似結果,土壤微生物對山合歡和銀合歡的生長并未造成顯著影響。對于產生這種現象的原因我們從以下幾個方面分析。
第一,試驗樣品可能受到了來自于空氣中微生物的污染。已有研究表明空氣中的微生物以氣溶膠的形式存在,具有6大特性,即來源的多相性、種類的多樣性、活性的易變性、播散的三維性、沉積的再生性、感染的廣泛性(孫平勇等,2010)。因而,對于未經微生物處理的樣品,空氣中的微生物可直接附著于樣品土壤中進行生長繁殖,爭奪植物生長可利用的各種營養物質,尤其以營腐生生長的微生物為主。試驗結果也表明,不接種微生物處理的土壤霉菌數顯著高于接種微生物處理,其產生的霉菌可能來自空氣微生物。對于經微生物處理后的樣品,空氣中的各種微生物在附著于樣品土壤上之后,與接種的微生物之間可能形成競爭關系,從而既影響植物對營養物質的利用,也可能抑制有益菌種對植物生長的促進作用。
第二,接種土壤經低溫凍存可能導致微生物活性降低。研究表明,土壤樣品經4個月、-20 ℃的低溫凍存顯著降低了微生物群落的代謝活性(周楊等,2015)。本研究中,試驗所用的接種土壤經過了大約兩個月的冷凍保藏,其中的微生物數量和活性可能已經減少了很多,從而導致微生物接種沒有發揮出應有的效果。
第三,微生物接種方式的影響。土壤微生物對植物的影響主要通過土壤微生物滅菌和土壤微生物接種的方法進行研究。由于土壤滅菌會造成土壤理化性質的改變,因此Shaw 等(1999)認為滅菌土上再接種新鮮土壤可作為非滅菌處理,并且接種密度在1%~15%之間能對生物量產生效應(Pernilla et al., 2010),具體操作上包括接種過1 mm篩的新鮮土壤、過1 mm篩的土壤懸液與過20 μm篩的土壤懸液,以接種過1 mm篩的新鮮土壤對植物的影響最大(van de Voorde et al., 2012)。本研究中,為了不影響土壤中 AMF,同時減少土壤動物如線蟲的影響,以接種土壤懸浮液的方式接種土壤微生物,接種后靜置一段時間就將盆缽放回氣候箱中。由于懸浮液并未完全下滲,還有一部分殘留在表層土壤,因此在氣候箱的風力和光照影響下懸浮液蒸干可能導致部分微生物未接種成功,造成接種的微生物的量并不是很充足,從而導致微生物接種處理和對照處理間無顯著差異。
本研究接種的土壤微生物群落中含有根瘤菌類群,但試驗結束時山合歡和銀合歡根系中根瘤數量還比較少。根據我們前期的試驗培養結果(數據未發表),在同樣的氣候培養箱條件和植株條件下,山合歡和銀合歡在燥紅土上培養80 d后分別產生了30個和80個根瘤,比本研究高了接近10倍。這可能是由于接種的微生物數量較少,根瘤菌少,從而使得根系中結瘤較少。此外,豆科植物不僅能與根瘤菌形成聯合共生體,而且形成AMF-豆科植物-根瘤菌三重共生體。AMF 是干熱河谷生態系統的重要組成成分,干熱河谷自然植被的建群種大多有豐富的AMF(Li amp; Zhao, 2005;Li et al., 2010),Li 和 Zhao(2005)研究發現山合歡和銀合歡的根際土壤中也都被檢測到豐富的 AMF 孢子。但是,我們的試驗基本沒有觀察到AMF的侵染。基于 FUNGuild 軟件對接種土的真菌功能類群進行分析發現,接種用土的真菌群落中共生營養型所占比例不到1%,在共生營養型中,山合歡和銀合歡土壤中的AMF相對豐度分別為4.2%和13.8%(數據未發表)。這說明,在接種用土的真菌群落中,AMF的相對豐度本身比較低,加之試驗所用的接種用土經過了一定時間的冷凍保存,使得進一步減少了AMF的數量,因此AMF侵染很少。
3.3 植物生長對土壤有效氮磷的影響
本研究中,種植山合歡和銀合歡降低了培養基土的有效氮含量,但提高了基土的有效磷含量。土壤有效氮含量降低主要與植物吸收氮有關,而土壤有效磷含量的提高可能與植物的根系分泌物有關。吳愛姣(2021)的研究發現,在低磷脅迫下豆科植物根際可以通過分泌磷酸酶和有機酸從而使其適應低磷環境;酸性磷酸酶可以活化有機磷,釋放出磷酸根離子,實現對土壤磷營養的改良(肖曉明等,2014);有機酸可以促進土壤中難溶性磷的活化,從而導致土壤有機磷含量增加(石貴陽,2021)。因此,豆科植物在低磷條件下能產生大量有機酸,從而促進根際酸化,提高土壤中磷的有效性(趙文杰等,2011)。干熱河谷燥紅土屬強固磷土壤,其磷素有效性低。本研究中,培養前基土的有效磷含量只有0.34 mg·kg-1,可能對植物生長造成了低磷脅迫。從山合歡和銀合歡的氮磷比來看,其氮磷比平均值為42,較高的氮磷比(N∶Pgt;20)通常暗示著磷限制(Zhang et al., 2015),表明山合歡和銀合歡確實存在低磷脅迫。因此,山合歡和銀合歡可能通過根系分泌磷酸酶和有機酸活化土壤中的有機磷,分泌質子和小分子有機酸,加速被吸附的磷溶解為有效磷而提高土壤有效磷含量。
4 結論
綜上所述,干旱對植物生長性狀具有一定的抑制作用,但山合歡較高的根生物量和根冠比以及干旱條件下較多的根瘤數量有利于植物對干旱條件的適應。因此,我們更推薦以本地種山合歡作為干熱河谷植被恢復樹種。另外,本研究中山合歡和銀合歡種植明顯提高了土壤有效磷含量,利用豆科植物的作用來提高燥紅土磷素有效性是值得探索的新途徑。
本試驗雖然取得了一定的顯著性結果,但仍然存在一些不足之處。本研究中,土壤微生物接種對山合歡和銀合歡生長都沒有造成顯著影響,這可能與本研究的微生物接種源和試驗處理方式有關,在后續研究中,可從以下兩點改進試驗:(1)確保微生物接種用土的有效性,采集新鮮土壤及時開展試驗。(2)更換微生物的接種方式。在樣品中接種培養基質10%的新鮮土壤,增加微生物數量,充分發揮接種土壤的微生物效益。另外,本研究結果主要是基于氣候箱模擬條件得出,與實際干熱河谷條件可能存在一定的差異,后續研究可在原位條件下進行以便更真實地反映山合歡和銀合歡的適應性差異。
參考文獻:
BERNS AE, PHILIPP H, NARRES HD, et al., 2008. Effect of gamma-sterilization and autoclaving on soil organic matter structure as studied by solid state NMR, UV and fluorescence spectroscopy" [J]. Eur J Soil Sci, 59: 540-550.
BI YL, CHEN SL, KONG WP, et al., 2014. Effects of microorganism inoculation on growth of soybean and its rhizosphere soil" [J]. Ecol Sci, 33(1): 121-126." [畢銀麗, 陳書琳, 孔維平, 等, 2014. 接種微生物對大豆生長及其根際土壤的影響 [J]. 生態科學, 33(1): 121-126. ]
CHEN ML, WANG YB, 2008. A comparative study on the physiological properties of the exotic species Oxalis corymbosa and the native species Oxalis corniculata under water stress" [J]. Acta Pratacul Sin, 17(6): 52-59." [陳明林, 王友保, 2008. 水分脅迫下外來種銅錘草和本地種酢漿草的生理指標比較研究 [J]. 草業學報, 17(6): 52-59.]
CUI YZ, LI K, SUN YY, et al., 2010. Phenotypic variation of geographic provenance of Albizia kalkora in Yunnan Province" [J]. Guizhou Agric Sci, 38(11): 98-101." [崔永忠, 李昆, 孫永玉, 等, 2010. 云南省山合歡地理種源變異性研究 [J]. 貴州農業科學, 38(11): 98-101.]
DA SILVA EC, NOGUEIRA R, DA SILVA MA, et al., 2011. Drought stress and plant nutrition" [J]. Plant Stress, 5(Special Issue 1): 32-41.
DUAN AG, ZHANG JG, HE CY, et al., 2013. Regional effect of photosynthesis and transpiration of the main tree species for vegetation restoration in dry season of the dry-hot river valleys" [J]. J Arid Land Resour Environ, 27(7): 148-154." [段愛國, 張建國, 何彩云, 等, 2013. 金沙江干熱河谷主要造林樹種干季光合與蒸騰作用區域效應分析 [J]. 干旱區資源與環境, 27(7): 148-154.]
HAFFANI S, MEZNI M, SLAMA I, et al., 2014. Plant growth, water relations and proline content of three vetch species under water-limited conditions" [J]. Grass Forage Sci, 69: 323-333.
HAN JJ, DUAN X, ZHAO YY, 2019.Spatial and temporal variability of soil moisture on slope land of different vegetation of dry-hot valley in Jinsha River" [J]. Arid Land Geogr, 42(1): 123-131." [韓姣姣, 段旭, 趙洋毅, 2019. 金沙江干熱河谷不同植被坡面土壤水分時空分布特征 [J]. 干旱區地理, 42(1): 123-131.]
IGIEHON NO, BABALOLA OO, 2018. Below-ground-above-ground plant-microbial interactions: focusing on soybean, rhizobacteria and mycorrhizal fungi" [J]. Open Microbiol J, 12: 261-279.
KUDOYAROVA GR, KHOLODOVA VP, VESELOV DS, 2013. Current state of the problem of water relations in plants underwater deficit" [J]. Russ J Plant Physiol, 60(2): 165-175.
LAU JA, LENNON JT, 2012. Rapid responses of soil microorganisms improve plant fitness in novel environments" [J]. Proc Natl Acad Sci, 109: 14058-14062.
LEI L, CHENG X, CAI XF, 2009. The space-time distribution of water content of desertification soil and their relationship with plant growth in karst rocky Guizhou" [J]. Guizhou Sci, 27(2): 50-54." [雷麗, 程星, 蔡雄飛, 2009. 貴州巖溶山區土壤含水量時空分布與植物生長關系研究 [J]. 貴州科學, 27(2): 50-54.]
LI L, XU ZF, WEI X, et al., 2008. Physiological compare Merremia boisiana with Pueraria lobata under drought stress and rewatering conditions" [J]. Guihaia, 28(6): 806-810." [李玲, 徐志防, 韋霄, 等, 2008. 金鐘藤和葛藤在干旱與復水條件下的生理比較 [J]. 廣西植物, 28(6): 806-810.]
LI LZ, HAN L, 2019. Effects of soil organic matter and moisture on greenhouse soil characteristics and plant growth" [J]. S China Agric, 13(14): 188-189." [李連智, 韓琳, 2019. 土壤有機質和水分對溫室土壤特性及植物生長的影響 [J]. 南方農業, 13(14): 188-189. ]
LI T, LI LF, SHA T, et al., 2010. Molecular diversity of arbuscular mycorrhizal fungi associated with two dominant xerophytes in a valley-type savanna, southwest China" [J]. Appl Soil Ecol, 44: 61-66.
LI T, ZHAO ZW, 2005. Arbuscular mycorrhizas in a hot and arid ecosystem in southwest China" [J]. Appl Soil Ecol, 29: 135-141.
LIANG ZP, LI LQ, WAN FH, et al., 2016. Feedback of soil biota on Ageratina adenophora growth and competitiveness with native plant: a comparison of different sterilization methods" [J]. Chin J Eco-Agric, 24(9): 1223-1230." [梁作盼, 李立青, 萬方浩, 等, 2016. 土壤微生物對紫莖澤蘭生長與競爭的反饋: 不同滅菌方法的比較 [J]. 中國生態農業學報, 24(9): 1223-1230. ]
LIU AL, FU CM, LUO ZY, et al., 2023. Comparative analysis of physiological and biochemical characteristics of six native grass species in dry and hot river valley of Yunnan under drought stress" [J]. J W China For Sci, 52(6): 47-54." [劉愛林, 伏春美, 羅中陽, 等, 2023. 干旱脅迫下云南干熱河谷區6個鄉土草種生理生化特性的比較分析 [J]. 西部林業科學, 52(6): 47-54.]
PERNILLA BE, PUTTEN WH, BAKKER EJ, et al., 2010. Plant-soil feedback: experimental approaches, statistical analyses and ecological interpretations" [J]. J Ecol, 98: 1063-1073.
SARDANS J, PENUELAS J, 2013. Plant-soil interactions in mediterranean forest and shrublands: impacts of climatic change" [J]. Plant Soil, 365: 1-33.
SHAW LJ, BEATONB Y, GLOVER LA, et al., 1999. Re-inoculation of autoclaved soil as a non-sterile treatment for xenobiotic sorption and biodegradation studies" [J]. Appl Soil Ecol, 11: 217-226.
SHEN RF, ZHAO XQ, 2015. Role of soil microbes in the acquisition of nutrients by plants" [J]. Acta Ecol Sin, 35(20): 6584-6591." [沈仁芳, 趙學強, 2015. 土壤微生物在植物獲得養分中的作用 [J]. 生態學報, 35(20): 6584-6591.]
SHI GY, 2021. Physiological mechanism and proteomics of soybean root response to low phosphorus stress" [D]. Guiyang: Guizhou University: 9-33." [石貴陽, 2021. 大豆根系響應低磷脅迫的生理機制及蛋白組學初探 [D]. 貴陽: 貴州大學: 9-33.]
SUN PY, LIU XL, LIU JL, et al., 2010. Research progress on air microorganism" [J]. Chin Agric Sci Bull, 26(11): 336-340." [孫平勇, 劉雄倫, 劉金靈, 等, 2010. 空氣微生物的研究進展 [J]. 中國農學通報, 26(11): 336-340.]
VAN VOORDE TFJ, PUTTEN WH, MARYIJIN BT, 2012. Soil inoculation method determines the strength of plant-soil interactions" [J]. Soil Biol Biochem, 55: 1-6.
WANG XM, YAN BG, ZHAO G, et al., 2017. Effects of microorganism on carbon, nitrogen and phosphorus of Dodonaea viscosa and the soils from different elevations in Yuanmou, Yunnan, China" [J]. Chin J Plant Ecol, 41(3): 311-324." [王雪梅, 閆幫國, 趙廣, 等, 2017. 云南元謀不同海拔土壤微生物對車桑子碳、氮、磷化學計量特征及土壤特性的影響 [J]. 植物生態學報, 41(3): 311-324.]
WANG XM, ZHAO L, YAN BG, et al., 2016. Morphological and physiological responses of Heteropogon contortus to drought stress in a dry-hot valley" [J]. Bot Stud, 57(1): 1-12.
WEI HG, YE HY, 1991. Study on the evaluation of site quality of soil moisture content in dry season in the dry-hot valley of Jinsha River" [J]. J West China For Sci (2): 48-51." [魏漢功, 葉厚源, 1991. 金沙江干熱河谷旱季土壤含水率評價立地質量的研究 [J]. 云南林業科技 (2): 48-51.]
WU AJ, 2021. Mechanisms of root responses to low phosphorus stress in different crop species/genotypes with contrasting root system" [D]. Xianyang: Research Center of Soil and Water Conservation and Ecological Environment, Chinese Academy of Sciences and Ministry of Education: 51-91. [吳愛姣, 2021. 不同根系類型作物/品種的根系對低磷脅迫的響應機制 [D]. 咸陽: 中國科學院教育部水土保持與生態環境研究中心: 51-91.]
XIAO XM, LIU JS, ZHOU C, et al., 2014. Difference of root exudates from macadamia seedlings under different phosphorus supply" [J]. Chin J Trop Crop, 35(2): 261-265." [肖曉明, 劉軍生, 周程, 等," 2014. 不同磷水平下澳洲堅果幼苗根系分泌物的差異 [J]. 熱帶作物學報, 35(2): 261-265.]
YANG ZY, SU JR, LUO D, et al., 2007. Progress and perspectives on vegetation restoration in the dry-hot valley" [J]. For Res, 20(4): 563-568." [楊振寅, 蘇建榮, 羅棟, 等, 2007. 干熱河谷植被恢復研究進展與展望 [J]. 林業科學研究, 20(4): 563-568.]
YANG GW, ROY J, VERESOGLOU SD, et al., 2020. Soil biodiversity enhances the persistence of legumes under climate change" [J]. New Phytol, 229(5): 2945-2956.
YANG JD, ZHANG ZM, SHEN ZH, et al., 2016. Review of research on the vegetation and environment of dry-hot valleys in Yunnan" [J]. Biodivers Sci, 24(4): 462-474." [楊濟達, 張志明, 沈澤昊, 等, 2016. 云南干熱河谷植被與環境研究進展 [J]. 生物多樣性, 24(4): 462-474.]
YANG YN, BA L, BAI XN, et al., 2010. An improved method to stain arbuscular mycorrhizal fungi in plant roots" [J]. Acta Ecol Sin, 30(3): 774-779." [楊亞寧, 巴雷, 白曉楠, 等, 2010. 一種改進的叢枝菌根染色方法 [J]. 生態學報, 30(3): 774-779.]
ZHANG L, WANG H, CHEN NN, et al., 2012. Effects of soil biotic communities on the seedling performance of native and invasive provenances of Triadica sebifera" [J]. J Biosaf, 21(1): 41-45." [張令, 王泓, 陳楠楠, 等, 2012. 土壤微生物對不同種源烏桕生長的影響 [J]. 生物安全學報, 21(1): 41-45. ]
ZHANG W, ZHAO J, PAN F, et al., 2015. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China" [J]. Plant Soil, 391: 77-91.
ZHANG X, ZANG R, LI C, 2004. Population differences in physiological and morphological adaptations of Populus davidiana seedlings in response to progressive drought stress" [J]. Plant Sci, 166: 791-797.
ZHANG YP, ZHAO FX, LIU XX, et al., 2005. Relationship between growth increment of Azadirachta indica and site condition in hot and arid valley" [J]. For Res, 18(1): 74-79." [張燕平, 趙粉俠, 劉秀賢, 等, 2005. 干熱河谷印楝生長與立地條件關系 [J]. 林業科學研究, 18(1): 74-79.]
ZHAO BR, YANG CF, 2023. Vegetation restoration techniques for difficult sites in dry and hot valley of Yuanjiang River" [J]. For Invent Plan, 48(6): 48-52." [趙保榮, 楊春鳳, 2023. 元江干熱河谷困難立地植被恢復技術 [J]. 林業調查規劃, 48(6): 48-52.]
ZHAO G, JIN J, HAN XQ, et al., 2019. Photosynthetic characteristics of three typical trees for vegetation restoration in dry season of dry-hot valley" [J]. SW China J Agric Sci, 32(1): 81-86." [趙廣, 金杰, 韓學琴, 等, 2019. 干熱河谷旱季3種典型植被恢復樹種光合生理特征 [J]. 西南農業學報, 32(1): 81-86.]
ZHAO L, LANG NJ, ZHENG K, et al., 2006. A study on eco-environmental characteristics of dry and hot valley of Yunnan Province" [J]. For Invent Plan, 31(3): 114-117. [趙琳, 郎南軍, 鄭科, 等, 2006. 云南干熱河谷生態環境特性研究 [J]. 林業調查規劃, 31(3): 114-117.]
ZHAO WJ, ZHANG LJ, CHANG Q, et al., 2011. The research progress of organic acids metabolism in leguminous plant under phosphorus deficiency" [J]. Pratacul Sci, 28(6): 1207-1213."" [趙文杰, 張麗靜, 暢倩, 等, 2011. 低磷脅迫下豆科植物有機酸分泌研究進展 [J]. 草業科學, 28(6): 1207-1213.]
ZHENG JH, CHEN HY, LI JQ, et al., 2017. The effects of different soil sterilization treatments on soil microbial activity" [J]. J Fudan Univ (Nat Sci), 56(6): 681-691." [鄭嘉慧, 陳鴻洋, 李金全, 等, 2017. 不同土壤滅菌方法對土壤微生物活性的影響 [J]. 復旦學報(自然科學版), 56(6): 681-691.]
ZHONG XH, 2000. Degradtion of ecosystem and ways of its rehabilitation and reconstruction in dry and hot valley" [J]. Resour Environ Yangtze Basin, 9(3): 376-383." [鐘祥浩, 2000. 干熱河谷區生態系統退化及恢復與重建途徑 [J]. 長江流域資源與環境, 9(3): 376-383.]
ZHOU J, ZHANG MY, 1998. Study on quantitative selection of drought resistant and heat resistant afforestation on species in Yuanmou" [J]. Yunnan For Sci Technol (3): 33-37. [周蛟, 張明友, 1998. 元謀抗旱耐熱造林樹種的定量選擇研究 [J]. 云南林業科技 (3): 33-37.]
ZHOU LL, LI SB, PAN H, et al., 2021. Characteristics of soil nutrient and enzyme activities in plantations of Eucalyptus urophylla × E. grandis and five Acacia species" [J]. J Trop Subtrop Bot, 29(5): 483-493." [周麗麗, 李樹斌, 潘輝, 等, 2021. 5種相思樹和尾巨桉人工林土壤養分和酶活性特征 [J]. 熱帶亞熱帶植物學報, 29(5): 483-493.]
ZHOU P, AN Y, WANG Z, et al., 2014. Characterization of gene expression associated with drought avoidance and tolerance traits in a perennial grass species" [J]. PLoS ONE, 9: e103611.
ZHOU Y, CUI H, ZHU HH, et al., 2015. Effects of long-term soil storage on the metabolic activity and functional groups of soil microbial community" [J]. Microbiol China, 42(6): 1017-1024." [周楊, 崔航, 朱紅惠, 等, 2015. 土壤樣品長期保存對微生物群落代謝活性和功能類群的影響 [J]. 微生物學通報, 42(6): 1017-1024.]
