單寧種類和劑量效應在反芻動物生產中的應用
2024-12-31 00:00:00張家軒葛振影劉汝杰仲慶振孫澤威
草地學報 2024年8期

摘要:單寧是一類廣泛分布于植物體內的多酚類物質,是植物自身產生的一種次級代謝產物。因其具有結合蛋白質和碳水化合物的特性,適宜劑量的單寧反而有利于反芻動物對營養物質的利用。但因不同植物分子量以及分子結構存在的差異導致不同單寧的特性也不盡相同。本文總結了單寧的化學性質以及結合蛋白質和碳水化合物的特性,重點闡明了不同特性及劑量的單寧在瘤胃內作用效果的差異,為日后合理利用單寧提供科學參考,為未來精準調控反芻動物營養提供新思路。
關鍵詞:植物單寧;反芻動物;瘤胃;蛋白質降解;碳水化合物發酵;微生物區系;生物氫化
中圖分類號:S816.7""" 文獻標識碼:A"""" 文章編號:1007-0435(2024)08-2337-09
Tannin Species and Dose Effects in Ruminant Production
ZHANG Jia-xuan1, GE Zhen-ying2, LIU Ru-jie2, ZHONG Qing-zhen2, SUN Ze-wei2*
(1. College of Forestry and Grass Science, Jilin Agricultural University, Changchun, Jilin Province 130118, China;
2. College of Animal science and technology, Jilin Agricultural University, Changchun, Jilin Province 130118, China)
Abstract:Tannins are a class of polyphenols widely distributed in plants and are secondary metabolites produced by plants. Because of their ability to bind proteins and carbohydrates,appropriate doses of tannins are beneficial to the utilization of nutrients by ruminants. The different properties of different tannins are mainly due to different plant sources as well as differences in the chemical formulas and molecular weights of the tannins. This paper summarized the chemical properties of tannins and the characteristics of binding proteins and carbohydrates,and focused on clarifying the differences of the effects of tannins with different characteristics and doses in the rumen,which could put forward a scientific reference for the rational utilization of tannins and provide new ideas for the future precise regulation of ruminant nutrition.
Key words:Plant tannins;Ruminant;Rumen;Protein degradation;Carbohydrate fermentation;Microflora;Biohydrogenation
單寧因其結合蛋白質,阻礙酶的消化以及獨特的苦澀味而影響適口性等因素,通常被認為是飼料中的抗營養因子[1-2]。但越來越多的研究表明,植物單寧中的水解單寧可替代單胃動物飼料中的抗生素和促生長添加劑,起到改善動物健康和提高動物生長性能的作用[3-5];也可用于抑制反芻動物瘤胃對營養物質的過度發酵,減少反芻動物飼料的營養損失和甲烷排放[6-7]。圍繞單寧的研究重心也逐漸由抗營養作用轉移到營養作用上。但不同種類、不同劑量單寧對反芻動物瘤胃內蛋白質的降解以及碳水化合物發酵的作用不同,適宜種類和劑量的單寧可有效調控飼料蛋白質和碳水化合物在瘤胃內的發酵降解,進而提高蛋白質和碳水化合物的利用效率,反之,則會降低飼料養分的利用率。因此,本文在簡述單寧的種類和化學性質的基礎上,重點闡述單寧在反芻動物生產中的利用方式,單寧在反芻動物瘤胃蛋白質降解、碳水化合物發酵、微生物區系變化以及生物氫化過程中的種類和劑量效應,為單寧在反芻動物生產中的科學應用和反芻動物營養的精準調控提供參考。
1 單寧的分類及化學性質
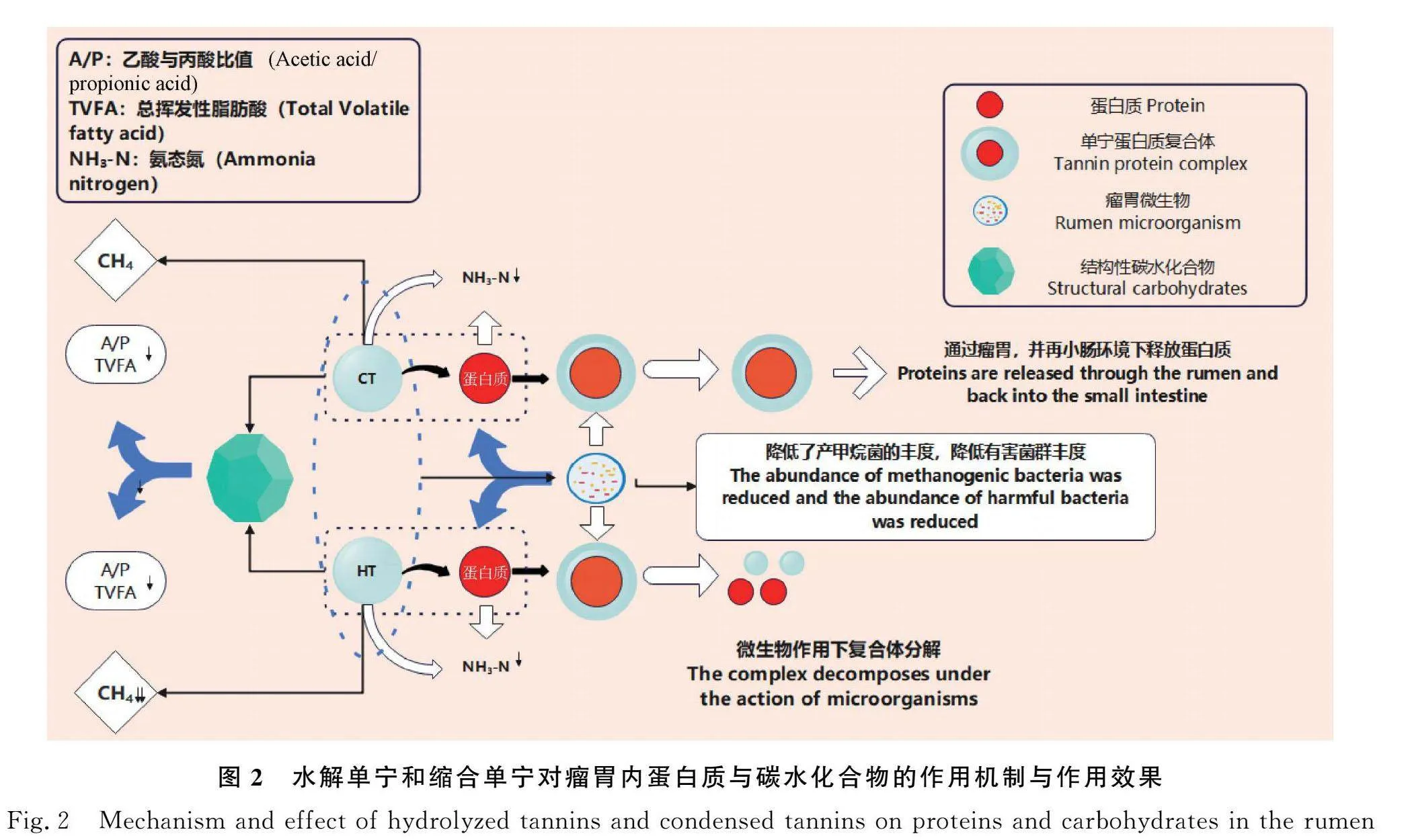
單寧又被稱為鞣酸、單寧酸,是植物體自身產生的一種植物次級代謝產物[8]。不同植物產生的單寧其組成和化學結構也有所不同,如圖1所示[14]。植物單寧通常可分為縮合單寧(Condensed tannins,CT)和水解單寧(Hydrolysable tannins,HT)[9]。縮合單寧又稱原花青素,是黃烷醇類植物多酚,是分子量最大的單寧,最大可達 20 000 Da[10],主要由兒茶素、表兒茶素、沒食子兒茶素和表沒食子兒茶素為結構單元聚合而成[11];水解單寧是以多元醇為核心的復雜分子,其分子量較小,僅有500~3000 Da[12],易被降解生成次級產物多元醇和酚酸等[6],其經過弱酸、弱堿、單寧酶或者微生物作用下會分解生成小分子有毒代謝產物[13]。
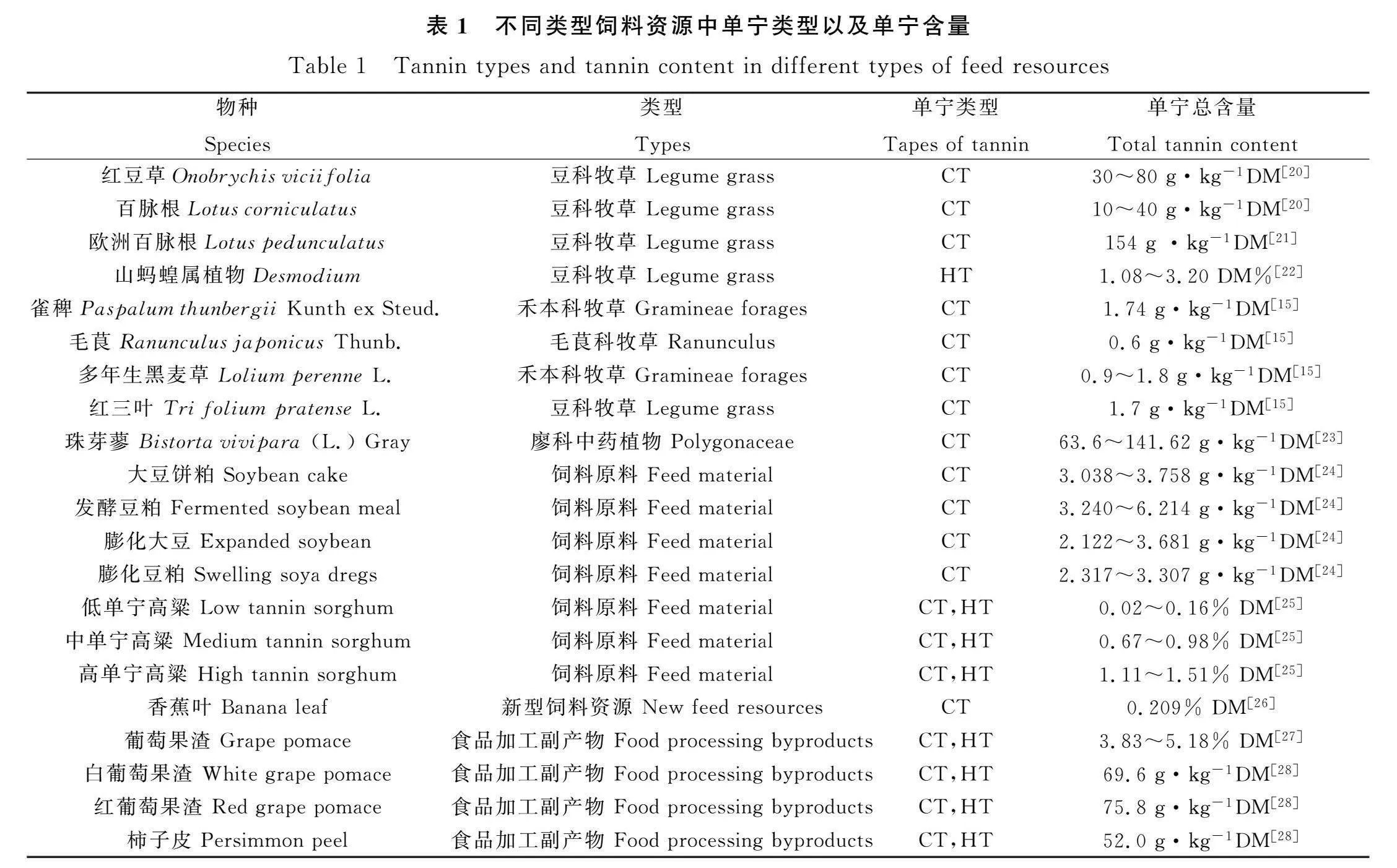
2 不同類型飼料資源中單寧種類和含量的差異
不同類型的飼料資源中含單寧的種類和含量也不盡相同(如表1所示),有些植物僅含有縮合單寧,有些植物僅含有水解單寧,也有些植物同時含有縮合單寧和水解單寧。牧草中單寧的主要類型為縮合單寧,但也有部分牧草富含水解單寧,同種屬的植物中單寧種類基本相同,但生長地區和環境等因素也會導致其單寧含量有較大差異[15],甚至植物不同器官單寧的種類和含量也有巨大的差異[16]。比如,美國山核桃果仁中既有水解單寧又有縮合單寧,含量分別為0.22 mg·g-1和4.66 mg·g-1;而美國山核桃皮中僅存在縮合單寧,含量在0.5~876 mg·g-1[17-18]。來自北美的三種朔樹雖屬于同屬植物,且所含單寧均為水解單寧,但單寧的含量差異巨大[19]。
3 單寧結合蛋白質和碳水化合物特性
3.1 單寧結合蛋白質特性
早在1962年,單寧被當時的學者定義為一類可以沉淀蛋白質的水溶性多酚類物質[29]。不同類型的單寧因其分子結構以及分子量大小不同,在瘤胃中對蛋白質的吸附程度以及復合體的類型也表現出較大的差異[30]。縮合單寧因其相對分子質量較大,同時通過共價鍵和非共價鍵與蛋白質結合,在此過程中發生電子的偏移,這種結合過程是不可逆的[31]。縮合單寧-蛋白質復合體在瘤胃內很難被瘤胃微生物降解;在進入真胃和小腸中可以大量地解體釋放蛋白質,供小腸吸收(如圖2所示)[6];水解單寧相對分子質量較小,并以共價鍵與蛋白質結合,在這個過程中,不會發生電子的偏移,因此與蛋白質結合后形成的水解單寧-蛋白質復合體可能會被瘤胃微生物所降解。
單寧與不同來源的蛋白質結合可以發揮不同作用。單寧可以通過競爭性結合菌體和寄生蟲營養所需的蛋白質抑制其生長;單寧也可以抑制或結合寄生蟲分泌的酶,進而直接影響到其機體代謝[32]。Villanuev等[33] 利用天然提取的五種單寧提取物作用于鼠傷寒沙門氏菌、銅綠假單胞菌、大腸桿菌和金黃色葡萄球菌,并觀察細菌膜的影響,研究結果表明,單寧對測定細菌表現出特異性的抗生物膜活性。Blomstrand等[34]研究表明,利用云杉樹皮提取縮合單寧并添加至奶牛日糧中,具有極強的抗球蟲作用。
在眾多常規飼料原料中也存在單寧[35],例如大豆、蠶豆[36]等。研究發現,高單寧飼料適口性下降的本質是單寧結合了口腔中唾液蛋白后,在吞咽時增加了摩擦性[37];高單寧飼料蛋白質消化率低主要歸結于單寧結合消化酶降低了動物的消化能力,單寧結合飼料中的蛋白質降低了飼料蛋白質的可消化性。
而在反芻動物中,單寧的蛋白質結合特異性使其可作為瘤胃蛋白質降解的有效調控物質。有研究發現,在日糧中添加低劑量的單寧有利于反芻動物瘤胃氮素的利用[38-40]。反芻動物日糧中存在大量可溶性蛋白質,日糧中單寧可于瘤胃內與可溶性蛋白形成單寧蛋白質復合體,其本質是單寧與蛋白質以氫鍵和疏水鍵的方式進行聚合反應[41]。由于單寧蛋白質復合體在瘤胃很難被降解,以過瘤胃蛋白的形式,到達真胃和小腸。而在真胃和小腸的酸性環境下,單寧蛋白質復合體解離釋放出蛋白質,從而增加了過瘤胃蛋白比例,提高了反芻動物蛋白質利用效率。
3.2 單寧結合碳水化合物特性
非結構性碳水化合物,如淀粉,在瘤胃中的消化不受單寧的影響,而高濃度的單寧降低了結構性碳水化合物和半纖維素的瘤胃消化[42]。在單寧與蛋白質結合后保持游離的部分單寧可通過與木質素纖維素絡合從而阻止微生物消化[43],亦或是單寧通過直接抑制調控纖維素分解的微生物[44]和纖維分解酶[45]的活性從而抑制纖維消化。適宜類型和劑量的單寧對碳水化合物在瘤胃中的發酵有一定積極的效應。有研究報道,單寧可改變瘤胃的發酵類型[3],減少反芻動物甲烷的排放[46]。目前有關單寧與碳水化合物結合的機理性文章較少,因此對于具體單寧與碳水化合物的結合方式以及規律仍需進一步地探討與研究。
4 單寧在反芻動物生產上的應用
目前為止,單寧在反芻動物上的應用主要分三方面:一是將富含單寧的植物及植物加工副產物添加至反芻動物日糧中;二是在粗飼料青貯過程中添加單寧;最后一方面是直接將單寧以添加劑的形式混合至反芻動物日糧中。
4.1 富含單寧的植物及加工副產物添加至反芻動物日糧中的應用
因存在飼料資源短缺的問題,新型飼料資源開發和利用成為了學者們研究的重點。大多數植物中富含天然單寧成分,開發此類植物作為新型飼料資源既有利于節約飼料,又對反芻動物生產具有積極的作用。隨著社會發展,例如葡萄渣、蘋果渣等這一類食品加工副產物的產量也逐年遞增,極大影響環境。因此將加工副產物添加至動物日糧中同樣也可以緩解飼料資源短缺[47-48],并改善環境。
向綿羊日糧中添加富含單寧的葡萄渣可顯著降低綿羊瘤胃氨態氮(NH3-N)的含量[49]。使用6%的印度楝葉替代部分日糧飼喂山羊,與未使用印度楝葉替代日糧組相比,顯著降低了瘤胃內NH3-N的濃度[50]。但是也有研究表明,利用富含單寧的牧草替代部分日糧飼喂動物顯著的增加了瘤胃內NH3-N的含量[51]。其本質原因可能在于不同植物中單寧的類型不同以及替代日糧比例的不同。在影響碳水化合物發酵方面,日糧中添加部分富含單寧的牧草可以減少甲烷(CH4)排放,并且隨著替代比例的增加,CH4的產量也顯著下降[52],揮發性脂肪酸(Volatile fatty acid,VFA) 的含量顯著增加,其中丙酸和丁酸的比例會顯著增加[53],但也有研究表明添加富含單寧的牧草會導致瘤胃內乙酸與丙酸比值顯著上升[54]。其原因可能在于植物內纖維性物質含量的差異。利用加工副產物替代動物日糧也得到相似的結論,將野櫻桃果渣按照500 g·kg-1DM的添加量替換部分日糧時顯著降低了干物質的瘤胃降解率,以500 g·kg-1DM的添加量將富含水解單寧的紅石榴果渣替換部分動物日糧時,顯著降低了甲烷的排放量[55]。這些研究表明,在利用了新型飼料資源以及加工副產物的同時,其內部的單寧發揮功效巨大,減緩了蛋白質的瘤胃降解率,同時也抑制了甲烷的排放,但不同植物對反芻動物瘤胃內蛋白質和碳水化合物的影響存在一定差異,這種差異歸根結底來源于植物中單寧的類型和含量不同。
4.2 單寧青貯的粗飼料對反芻動物瘤胃發酵的影響
單寧是一種優秀的青貯添加劑,在青貯的過程中,飼料中的真蛋白質會隨著微生物的生命活動分解成非蛋白氮,因此單寧的添加可以有效的結合真蛋白質,防止其過度降解,進一步提高青貯飼料品質[56-57]。同時,這種方式青貯的飼料中的蛋白質同樣也難以在瘤胃中降解。有研究表明,經過單寧青貯的飼料被反芻動物攝入后,飼料中蛋白質的瘤胃降解率顯著下降[4,58],這由于單寧在青貯的過程中結合蛋白,導致蛋白質組分發生了改變,最終導致瘤胃快速降解蛋白占比顯著下降,瘤胃慢速降解蛋白以及瘤胃不可降解蛋白的比例顯著升高[59]。陳雷等人的研究表明,在青貯過程中添加4% DM的單寧酸,與對照組相比顯著的增加了干物質量和粗蛋白質含量,顯著抑制了真蛋白質的降解。在后續的體外瘤胃發酵試驗中,添加單寧組顯著增加了瘤胃菌體蛋白的數量,同時NH3-N的含量顯著下降[60],這可能由于單寧抑制了瘤胃中蛋白質分解菌的菌群數量所導致[61]。但也有研究表明,與對照組相比添加單寧青貯葛藤莖葉對粗蛋白質含量無顯著影響,但是顯著的降低了青貯過程中黃曲霉毒素、嘔吐毒素以及玉米赤霉烯酮的含量。這證明,單寧在青貯的過程中抑制了各種雜菌的生長[62]。
反芻動物在進食添加單寧青貯的粗飼料時同樣會改變瘤胃發酵模式[4],瘤胃中乙酸的含量顯著降低,丙酸含量顯著上升,但對于產氣量的影響有著很大的差異。謝小來等[63]研究表明,分別以0.2%,0.4%,0.6% DM的單寧酸青貯紫花苜蓿,體外產氣量隨著單寧酸添加劑量的增加而增加;但也有研究表明,添加10 g·kg-1DM,20 g·kg-1DM,40 g·kg-1DM和60 g·kg-1DM的五倍子鞣酸青貯紫花苜蓿,各處理組中性洗滌纖維瘤胃降解率隨著五倍子鞣酸劑量的增加而顯著降低,同時各處理組的體外瘤胃產氣量也隨著五倍子鞣酸劑量的增加而顯著下降。
4.3 直接添加單寧至反芻動物日糧中的應用
單寧對反芻動物瘤胃內的影響主要包括對蛋白質的降解、碳水化合物的發酵、瘤胃內微生物區系以及瘤胃內生物氫化的影響。
4.3.1 單寧種類和劑量效應對瘤胃蛋白質降解的影響 目前直接添加單寧至反芻動物日糧中對瘤胃蛋白質降解影響的研究主要表現在對單寧種類效應或劑量效應的單一層面,對于單寧的種類以及劑量的混合效應研究較少。
單一使用一種單寧時無論使用何種單寧,隨著劑量的增加,瘤胃中NH3-N均顯著下降;對比兩種單寧對蛋白質降解的影響,縮合單寧對日糧中瘤胃慢速降解蛋白組分具有更強的保護性[64];還有研究表明,單寧可以減緩支鏈氨基酸脫氨,進而導致異VFA的濃度下降[65]。這些研究結果充分的表明單寧會與飼料中的蛋白質結合,可以減少蛋白質在瘤胃中的過度降解,增加過瘤胃蛋白的比例。使用混合單寧的研究結果表明,隨著混合單寧添加劑量的增加,NH3-N濃度和異揮發性脂肪酸含量均顯著降低[66]。Mezzomo等人[67]研究表明,將縮合單寧和水解單寧混合后添加至日糧中降低了飼料蛋白質在瘤胃中的降解率。
不同類型的單寧對蛋白質的結合能力也不同,混合單寧相對結合效果會更強。但是,蛋白質對單寧的親和力由分子量的大小決定,蛋白質分子量越大、其三級結構越復雜則表現出來的親和性越強[68]。因此,日糧中快、中、慢速降解蛋白的比例可能影響單寧的結合。目前的研究結果僅從NH3-N含量的下降,瘤胃蛋白質的降解率下降體現出單寧的積極意義。但目前的研究結果并未明確單寧對瘤胃內氨態氮含量影響的動態范圍,以及對反芻動物機體健康是否存在負面影響。混合使用單寧對瘤胃內蛋白質具體組分的影響也并未有很深入的研究。
4.3.2 單寧種類和劑量效應對瘤胃碳水化合物發酵的影響 單寧的種類和劑量效應對于瘤胃碳水化合物發酵也存在一定影響。在甲烷產量方面,縮合單寧和水解單寧均能顯著降低CH4產量,這意味著減少了能量的損失,因此單寧可以增加動物攝入日糧中能量的利用效率[69],但不同種類的單寧對于CH4產量的影響是有顯著差異的,水解單寧只降低了24~48 h產氣量;而縮合單寧影響了整個發酵時期的CH4產量[70]。但也有研究表明,以15 g·kg-1DM,30 g·kg-1DM和60 g·kg-1DM為劑量添加至以豆粕為發酵底物的體外發酵體系中,水解單寧組15 g·kg-1DM條件下24小時產氣量顯著下降,而縮合單寧組在60 g·kg-1DM劑量下24小時產氣量顯著下降[71]。水解單寧與縮合單寧對于產氣量的影響是不同的,相比較于單一使用水解單寧,混合使用單寧對產氣的抑制性更強[72]。
在影響瘤胃內VFA層面,縮合單寧的添加均會導致TVFA含量顯著降低,丙酸的含量顯著上升,乙酸的含量顯著下降,由乙酸發酵轉變為丙酸發酵[70,73],在抑制碳水化合物發酵的層面更加有效;但使用水解單寧添加至動物日糧中,會顯著提高VFA的含量[74],這是因為水解單寧在被微生物代謝后生成了鞣花酸,鞣花酸進一步代謝可以生成乙酸[75]。Menci等人[66]對比了混合單寧和單一縮合單寧對野豌豆鮮草和干草兩種基質體外發酵的差異。試驗結果表明,作用于干草基質時,縮合單寧相較于混合單寧組,乙酸含量顯著下降,乙酸與丙酸的比值顯著下降。這表明,混合單寧中存在水解單寧的組分,因此增加了乙酸的組分。
單寧在反芻動物瘤胃中對碳水化合物的影響是由多方面因素限制的。不僅取決于單寧的類型、劑量以及反芻動物日糧的組成結構,還取決于動物的個體以及種屬差異。前人的研究中僅對單一單寧和混合單寧進行比較,有關不同單寧混合比例對碳水化合物發酵影響的文章相對較少,同時,單寧對瘤胃發酵參數的影響并未有明確的參考范圍,因此不能確定單寧是否對動物機體產生影響。
4.3.3 單寧種類和劑量效應對瘤胃微生物的影響 瘤胃內相互共生的微生物發揮著豐富而強大的功能,對于調控瘤胃能氮平衡起重要的作用。不同單寧因其分子結構不同對微生物的群落和類別也有著顯著不同的影響。且單寧的比例效應和劑量效應對于瘤胃內微生物的影響也同樣具有差異。有研究表明,縮合單寧對于蛋白質水解細菌溶纖維丁酸弧菌(B.fibrisolvens)、嗜淀粉瘤胃桿菌(Ruminobacter amylophilus)以及牛鏈球菌(Streptococcus bovis)的菌群豐度影響巨大,但對棲瘤胃普雷沃氏菌(Prevotella ruminicola)的影響較小[76]。這表明,單寧對蛋白質瘤胃降解率的影響與蛋白質水解菌的菌群豐度有著密切的聯系。研究證實,幾乎所有類型的單寧添加至反芻動物日糧中均有效降低甲烷桿菌(Methanobacterium)以及產甲烷古菌(Methanogenic Archaea)的菌群豐度[77-78],這也從微生物的角度印證了單寧可以顯著降低瘤胃甲烷的產量。Carrasco等人按照1∶2的比例將栗木水解單寧和堅木縮合單寧進行混合添加至荷斯坦奶牛日糧中,研究結果表明,混合單寧增加了瘤胃內厚壁菌門(Phylum Firmicutes)以及擬桿菌門(Bacteroidetes)的菌群豐度,顯著的降低了淀粉分解菌(Amylolytic bacteria)以及纖維分解菌(Trichoderma harzianum)的豐度[79]。
4.3.4 單寧種類和劑量效應對瘤胃內生物氫化的影響 單寧以兩種途徑調節瘤胃內生物氫化,一是影響生物氫化相關的微生物,二是影響生物氫化中間體的影響。Mnica等人[74]研究結果表明,部分細菌可以參與瘤胃內脂肪酸的調控,縮合單寧相較于水解單寧顯著降低了黃色瘤胃球菌(Ruminococcus flavefaciens)、白色瘤胃球菌(Ruminococcus albus)、溶纖維丁酸弧菌的菌群豐度。溶解纖維丁酸弧菌對于生物加氫反應具有一定的影響[80],而黃色瘤胃球菌以及白色瘤胃球菌對于生物氫化過程中最后一步,由中間體轉化成C18∶0起到重要作用[81]。因此,在單寧種類層面,縮合單寧對于生物氫化的影響要高于水解單寧。縮合單寧和水解單寧均會降低RA/LA(瘤胃酸/亞油酸)的比值,即影響了生物氫化的第一步過程[66]。從劑量效應的層面來看,隨著單寧劑量的增加,顯著的提升了α-亞麻酸(ALA)、LA以及油酸(OA)的含量;降低了硬脂酸(SA),C15∶0,支鏈C15∶0,C17∶0以及支鏈C17∶0脂肪酸的含量[66]。在四個單寧種類(兩種縮合單寧和兩種水解單寧)和四個劑量的兩因素試驗中,僅橡樹水解單寧在20 g·kg-1DM的劑量下顯著提高了總多不飽和脂肪酸(tPUFA),C18∶3n-3,C18∶2n-6,t-11 C18∶1的含量,降低了t-10 C18∶1以及C18∶0的含量。這表明,即便單寧種類相同,但是植物來源不同,對于瘤胃生物加氫的影響也是不同的[82]。
5 小結與展望
單寧不僅能通過其抗氧化性、抗菌性和抗寄生蟲性來保護反芻動物腸道,從而保證動物機體健康,還因其結合蛋白質和碳水化合物的特性,在反芻動物生產中,可以減少蛋白質在瘤胃中的過度降解,減少甲烷的排放,改善瘤胃的發酵方式;同時單寧亦可以降低瘤胃中產甲烷類型的菌群豐度以及生物氫化相關菌群豐度,提高共軛亞油酸等脂肪酸含量,降低C18∶0的含量。但由于單寧種類、動物種類以及日糧結構的差異,使得研究結果具有一定的差異。在反芻動物生產過程中,應以地區的飼料資源特異性為主,使用當地富含單寧的特色飼料資源或提取的單寧提取物。隨著未來科技的進步,單寧將會被越來越廣泛的應用于健康養殖領域。
單寧的種類以及植物來源眾多,目前仍有很多問題亟待解決,現有研究對于單寧的種類和劑量效應仍不完善,未來可以從以下幾個方面進行深入研究:①完善不同種類動物飼養過程中最適宜的單寧種類和使用閾值;②完善單寧對不同品種粗飼料青貯過程中最適的青貯時間以及青貯工藝,最大限度的提升青貯飼料的品質;③深入研究單寧對反芻動物瘤胃代謝的機制,以及單寧在胃后消化道的作用機制,完善單寧對消化道微生物影響機制的研究。最終實現單寧在反芻動物生產上的廣泛應用,精準調控反芻動物營養以及反芻動物機體健康。
參考文獻
[1] 宋青龍,秦貴信. 飼料中的抗營養因子及其消除方法[J]. 國外畜牧學(豬與禽),2003(3):9-12
[2] 王沖,婁玉杰. 常見飼料中抗營養因子及對動物的影響[J]. 家畜生態,2000(4):39-43
[3] CHEN L,BAO X Y,GUO G. Effects of Hydrolysable Tannin with or without Condensed Tannin on Alfalfa Silage Fermentation Characteristics and In Vitro Ruminal Methane Production,Fermentation Patterns,and Microbiota[J]. Animals,2021,11(7):1967-1985
[4] LEI C,XUEYAN B,GANG G,et al. Treatment of alfalfa silage with tannin acid at different levels modulates ensiling characteristics,methane mitigation,ruminal fermentation patterns and microbiota[J]. Animal Feed Science and Technology,2021(278):114997
[5] SONG Y Y,LUO Y,YU B. Tannic acid extracted from gallnut prevents post-weaning diarrhea and improves intestinal health of weaned piglets[J]. Animal Nutrition,2021,7(4):1078-1086
[6] PATRA A K,SAXENA J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition[J]. Journal of the Science of Food Agricultural,2011,91(1):24-37
[7] WANG Y S,WANG L Z,WANG Z S. Recent advances in research in the rumen bloat of ruminant animals fed high-concentrate diets[J]. Frontiers in Veterinary Science,2023(10):339-347
[8] 李元恒,金龍,韓國棟,等. 植物單寧在反芻動物營養和健康養殖作用中的研究進展[J]. 草地學報,2013,21(6):1043-1051
[9] HUANG Q,LIU X,ZHAO G. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production[J]. Animal Nutrition,2018,4(2):137-150
[10]ZHANG L L,GUAN Q H,ZHANG H. Effect of metal ions on the interaction of condensed tannins with protein[J]. Foods,2023,12(4):829-829
[11]MCSWEENEY C S,PALMER B,MCNEILL D M. Microbial interactions with tannins:nutritional consequences for ruminants[J]. Animal Feed Science and Technology,2001,91(1-2):83-93
[12]ARAPITSAS P. Hydrolyzable tannin analysis in food[J]. Food Chemistry,2012,135(3):1708-1717
[13]BESHARATI M,MAGGIOLINO A,PALANGI V. Tannin in ruminant nutrition:Review[J]. Molecules,2022,27(23):8273-8299
[14]梁澤毅,張劍搏,鄭娟善,等. 單寧的生物學功能及其在反芻動物營養中的研究進展[J]. 動物營養學報,2020,32(11):5059-5068
[15]JACKSON F S,MCNABB W C,BARRY T N. The condensed tannin content of a range of subtropical and temperate forages and the reactivity of condensed tannin with ribulose- 1,5-bis-phosphate carboxylase(Rubisco)protein[J]. Journal of the Science of Food and Agriculture,1996,72(4):483-492
[16]ADAMCZYK B,SIMON J,KITUNEN V. Tannins and their complex interaction with different organic nitrogen compounds and enzymes:Old paradigms versus recent advances[J]. Chemistry Open,2017,6(5):610-614
[17]DANIEL E M,KRUPNICK A S,HEUR Y H. Extraction,stability,and quantitation of ellagic acid in various fruits and nuts[J]. Journal of Food Composition amp; Analysis,1989,2(4):338-349
[18]ALVAREZ-PARRILLA E,ROSA L,URREA-LPEZ R. Bioactive components and health effects of pecan nuts and their byproducts:a review[J]. Journal of Food Bioactives,2018(1):56-92
[19]PRIDA A,PUECH J L. Influence of geographical origin and botanical species on the content of extractives in American,French,and East European oak woods[J]. Journal of Agriculture and Food Chemistry,2006,54(21):8115-8126
[20]LAGRANGE S P,MACADAM J W,VILLALBA J J. The use of temperate tannin containing forage legumes to improve sustainability in forage-livestock production[J]. Agronomy-Basel,2021,11(11):18-28
[21]GEBREHIWOT L,BEUSELINCK P R,ROBERTS C A. Seasonal variations in condensed tannin concentration of three lotus species[J]. Agronomy Journal,2002,94(5):1059-1065
[22]陳艷琴. 山螞蝗屬植物單寧含量動態變化及其飼用價值研究[D]. 海口:海南大學,2011:18
[23]陳子凡. 不同生長期珠芽蓼縮合單寧的含量與結構及其對體外瘤胃發酵的影響[D]. 蘭州:蘭州大學,2022:46
[24]李丹,邱靜,谷旭,等. 不同大豆原料中單寧含量的差別分析[J]. 飼料工業,2016,37(1):38-41
[25]馮淦熠,李瑞,黃瑞林,等. 高粱的營養價值與抗營養因子及在豬和家禽生產中的應用[J]. 動物營養學報,2022,34(4):2073-2086
[26]劉中英,王春芳,曹兵海. 香蕉葉與香蕉全株營養成分和單寧含量的比較分析[J]. 飼料工業,2014,35(1):32-33
[27]張歡,王麗慧,李冰,等. 釀酒葡萄渣單寧及主要營養成分含量分析[J]. 飼料工業,2019,40(19):11-15
[28]FITRI A,OBITSU T,SUGINO T. Effect of ensiling persimmon peel and grape pomace as tannin-rich byproduct feeds on their chemical composition and in vitro rumen fermentation[J]. Animal Science Journal,2021,92(1):e13524
[29]KOCZWARA M. Recent studies on tannins of vegetable origin[J]. Farmacja Polska,1962(18):127-132
[30]SILVA S A,NABAVI F S,SAEEDI M. Recent advances in natural products analysis[M]. 1st edition. Amsterdam:Elsevier,2020:132-146
[31]MUELLER-HARVEY I. Unravelling the conundrum of tannins in animal nutrition and health[J]. Journal of the Science and Food Agricultural,2006,86(13):2010-2037
[32]FRAGA-CORRAL M,GARCIA-OLIVEIRA P,PEREIRA A G. Technological application of tannin-based extracts[J]. Molecules,2020,25(3):614-641
[33]VILLANUEVA X,ZHEN L,ARES J N. Effect of chemical modifications of tannins on their antimicrobial and antibiofilm effect against Gram-negative and Gram-positive bacteria[J]. Frontiers Microbiology,2023(13):4905
[34]BLOMSTRAND B M,ENEMARK H L,STEINSHAMN H. Administration of spruce bark (Picea abies) extracts in young lambs exhibits anticoccidial effects but reduces milk intake and body weight gain[J]. Acta Veterinaria Scandinavica,2022,64(1):629-639
[35]馮定遠,汪儆. 抗營養因子及其處理研究進展[C] //中國畜牧獸醫學會.2000動物營養研究進展—全國畜禽飼養標準學術討論會暨營養研究會成立大會論文集. 北京:中國農業出版社,2000:18
[36]夏艷秋,劉汝杰,趙雨菲,等. 蠶豆中非營養性化合物的生物活性及其加工鈍化研究進展[J]. 中國畜牧雜志,2023,7(59):54-58
[37]ISABELLE L,NOBLE C A. Polyphenols:factors influencing their sensory properties and their effects on food and beverage preferences[J]. The American Journal of Clinical Nutrition,2005,81(1):330-335
[38]ALIPOUR D,ROUZBEHAN Y. Effects of several levels of extracted tannin from grape pomace on intestinal digestibility of soybean meal[J]. Livestock Science,2010,128(1-3):87-91
[39]AVILA S C,KOZLOSKI G V,ORLANDI T. Impact of a tannin extract on digestibility,ruminal fermentation and duodenal flow of amino acids in steers fed maize silage and concentrate containing soybean meal or canola meal as protein source[J]. Journal of the Agriculture Science,2015,153(5):943-953
[40]DENTINHO M T P,BELO A T,BESSA R J B. Digestion,ruminal fermentation and microbial nitrogen supply in sheep fed soybean meal treated with Cistus ladanifer L. tannins[J]. Small Ruminant Research,2014,119(1-3):57-64
[41]FONSECA N V,CARDOSO A D,BAHIA A S. Additive tannins in ruminant nutrition:An alternative to achieve sustainability in animal production[J]. Sustainability,2023,15(5):4162
[42]BARRY T N,MANLEY T R,DUNCAN S J. The role of condensed tannins in the nutritional value of Lotus pedunculatus for sheep. 4. Sites of carbohydrate and protein digestion as influenced by dietary reactive tannin concentration[J]. The British Journal of Nutrition,1986,55(1):123-137
[43]BARRY T N,MANLEY T R. Interrelationships between the concentrations of total condensed tannin,free condensed tannin and lignin in Lotus sp. and their possible consequences in ruminant nutrition[J]. Journal of the Science and Food Agricultural,1986,37(3):248-254
[44]PATAR A K,JYOTISNA S. Dietary phytochemicals as rumen modifiers:a review of the effects on microbial populations[J]. Antonie Van Leeuwenhoek,2009,96(4):363-375
[45]BAE H D,MCALLISTER T A,YANKE J. Effects of condensed tannins on endoglucanase activity and filter paper digestion by fibrobacter succinogenes S85[J]. Applied and Environmental Microbiology,1993,59(7):2132-2138
[46]WITZIG M,ZEDER M,RODEHUTSCORD M. Effect of the ionophore monensin and tannin extracts supplemented to grass silage on populations of ruminal cellulolytics and methanogens in vitro[J]. Anaerobe,2018(50):44-54
[47]BARTEL I,KOSZARSKA M,WYSOCKI K. Effect of dried apple pomace (DAP) as a feed additive on antioxidant system in the rumen fluid[J]. International Journal of Molecular Sciences,2022,23(18):10475
[48]ISHIDA K,KISHI Y,OISHI K. Effects of feeding polyphenol-rich winery wastes on digestibility,nitrogen utilization,ruminal fermentation,antioxidant status and oxidative stress in wethers[J]. International Journal of Molecular Sciences,2015,86(3):260-269
[49]ABARGHUEI M J,ROUZBEHAN Y,ALIPOUR D. The influence of the grape pomace on the ruminal parameters of sheep[J]. Livestock Science,2010,132(1):73-79
[50]TAETHAISONG N,PAENGKOUM S,KAEWWONGSA W. The effect of neem leaf supplementation on growth performance,rumen fermentation,and ruminal microbial population in goats[J]. Animals,2023,13(5):890-904
[51]CARDOSO-GUTIERREZ E,ARANDA-AGUIRRE E,ROBLES-JIMENEZ L E. Effect of tannins from tropical plants on methane production from ruminants:A systematic review[J]. Veterinary and Animal Science,2021(14):100214
[52]VARGAS-ORTIZ L,CHAVEZ-GARCIA D,BARROS-RODRIGUEZ M. Rumen function and in vitro gas production of diets influenced by two levels of tannin-rich forage[J]. Fermentation,2022,8(11):607-618
[53]ALBORES-MORENO S,AlAVN-GAMBOA J A,MIRANDA-ROMERO L A. Effect of tree foliage supplementation of tropical grass diet on in vitro digestibility and fermentation,microbial biomass synthesis and enteric methane production in ruminants[J]. Tropical Animal Health and Production,2018,51(4):893-904
[54]SUYBENG B,CHARMLEY E,GARDINER C P. Supplementing northern australian beef cattle with desmanthus tropical legume reduces in-vivo methane emissions[J]. Animals,2020,10(11):2097-2114
[55]GILLER K,BOSSUT L,EGGERSCHWILER L. In vitro ruminal fermentation,methane production and nutrient degradability as affected by fruit and vegetable pomaces in differing concentrations[J]. Journal of Animal Physiology Animal Nutrition,2022,106(5):957-967
[56]JAYANEGARA A,YAMAN A,KHOTIJAH L. Reduction of proteolysis of high protein silage from Moringa and Indigofera leaves by addition of tannin extract[J]. Veterinary World,2019,12(2):211-217
[57]TABACCO E,BORREANI G,CROVETTO G M. Effect of chestnut tannin on fermentation quality,proteolysis,and protein rumen degradability of alfalfa silage[J]. Journal of Dairy Science,2006,89(12):4736-4746
[58]趙夢迪. 添加低水平縮合單寧對青貯野火球發酵品質及延邊黃牛瘤胃體外發酵影響[D]. 延邊:延邊大學,2020:24
[59]DENTINHO M T P,PAULOS K,PORTUGAL P V. Proteolysis and in situ ruminal degradation of lucerne ensiled with Cistus ladanifer tannins[J]. Grass and Forage Science,2019,74(1):78-85
[60]陳雷,暴雪艷,郭剛,等. 單寧酸和乳酸菌對紫花苜蓿青貯品質和體外瘤胃發酵的影響[J]. 草地學報,2021,29(8):1853-1858
[61]JOLAZADEH A R,DEHGHAN-BANADAKY M,REZAYAZDI K. Effects of soybean meal treated with tannins extracted from pistachio hulls on performance,ruminal fermentation,blood metabolites and nutrient digestion of Holstein bulls[J]. Animal Feed Science and Technology,2015,(203):33-40
[62]楊冬梅,陶雙倫,梁靜,等. 添加單寧酸對青貯葛藤莖葉品質的影響[J]. 草地學報,2012,20(4):784-787
[63]謝小來,馬逢春,焦培鑫,等. 添加單寧酸對紫花苜蓿青貯品質及瘤胃體外產氣量的影響[J]. 東北農業大學學報,2021,52(8):48-56
[64]徐曉鋒. 縮合單寧與水解單寧對奶牛日糧氮利用影響及作用機制的研究[D]. 北京:中國農業大學,2013:33-34
[65]趙淵. 不同劑量單寧酸對育肥牛生產性能、瘤胃發酵、血液生化及免疫功能的影響[D]. 長沙:湖南農業大學,2021:17
[66]MENCI R,COPPA M,TORRENT A. Effects of two tannin extracts at different doses in interaction with a green or dry forage substrate on in vitro rumen fermentation and biohydrogenation[J]. Animal Feed Science and Technology,2021(278):114997
[67]MEZZOMO R,PAULINO P V R,DETMANN E. Tannin on non-degradable digestible protein from proteic sources in cattle rumen[J]. Acta Scientiarum:Animal Sciences,2015,37(4):389-395
[68]ASQUITH T N,BUTLER L G. Interactions of condensed tannins with selected proteins[J]. Phytochemistry,1986,25(7):1591-1593
[69]BEAUCHEMIN K A,UNGERFELD E M,ECKARD R J. Review:Fifty years of research on rumen methanogenesis:lessons learned and future challenges for mitigation[J]. Animal,2020(14):S2-S16
[70]MARCO B,STEFANIA C,PIETRO P. In vitro effects of different levels of quebracho and chestnut tannins on rumen methane production,fermentation parameters,and microbiota[J]. Frontiers in Veterinary Science,2023(10):470-482
[71]LAVRENCIC A,PIRMAN T. In vitro gas and short-chain fatty acid production from soybean meal treated with chestnut and quebracho wood extracts by using sheep rumen fluid[J]. Journal of Animal and Feed Sciences,2021,30(4):312-319
[72]BHATTA R,UYENO Y,TAJIMA K. Difference in the nature of tannins on in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations[J]. Journal of Dairy Science,2009,92(11):5512-5522
[73]黃小乘,劉思揚,田怡豪,等. 低水平縮合單寧對以豆粕和苜蓿為底物的體外發酵特性的影響[J]. 飼料工業,2022,43(13):38-43
[74]COSTA M, ALVES C P, CAPPUCCT A, et al. Effects of condensed and hydrolyzable tannins on rumen metabolism with emphasis on the biohydrogenation of unsaturated fatty acids[J]. Journal of Agricultural and Food Chemistry,2018,66(13):3367-3377
[75]WAGHORN G. Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production-Progress and challenges[J]. Animal Feed Science and Technology,2008,147(1-3):116-139
[76]JONES G A,MCALLISTER T A,MUIR A D. Effects of Sainfoin (Onobrychis viciifolia Scop.) condensed tannins on growth and proteolysis by four strains of ruminal bacteria[J]. Applied and Environmental Microbiology,1994,60(4):1374-1378
[77]MALIK P K,TRIVEDI S,KOLTE A P. Effect of an anti-methanogenic supplement on enteric methane emission,fermentation,and whole rumen metagenome in sheep[J]. Frontiers Microbiology,2022(13):4678-4686
[78]CHEN L,BAO X Y,GUO G. Treatment of alfalfa silage with tannin acid at different levels modulates ensiling characteristics,methane mitigation,ruminal fermentation patterns and microbiota[J]. Animal Feed Science and Technology,2021(278):114997
[79]CARRASCO J M D,CABRAL C,REDONDO L M. Impact of chestnut and quebracho tannins on rumen microbiota of bovines[J]. Biomed Research International,2017(2017):1-11
[80]BAINBRIDGE M L,CERSOSIMO L M,WRIGHT A-D G. Rumen bacterial communities shift across a lactation in Holstein,Jersey and Holstein×Jersey dairy cows and correlate to rumen function,bacterial fatty acid composition and production parameters[J]. FEMS Microbiology Ecology,2016,92(5):fiw059
[81]GUDLA P,ABUGHAZALEH A A,ISHLAK A. The effect of level of forage and oil supplement on biohydrogenation intermediates and bacteria in continuous cultures[J]. Animal Feed Science and Technology,2012,171(2):108-116
[82]CARREO D,HERVS G,TORAL P G. Ability of different types and doses of tannin extracts to modulate in vitro ruminal biohydrogenation in sheep[J]. Animal Feed Science and Technology,2015(202):42-51
(責任編輯 閔芝智)
