功能性狀對鄉土植物混播草地產量及穩定性的影響
2024-12-31 00:00:00禹安然楊彥澤喬冠棟張俊董一丁侯向陽路文杰
草地學報 2024年8期
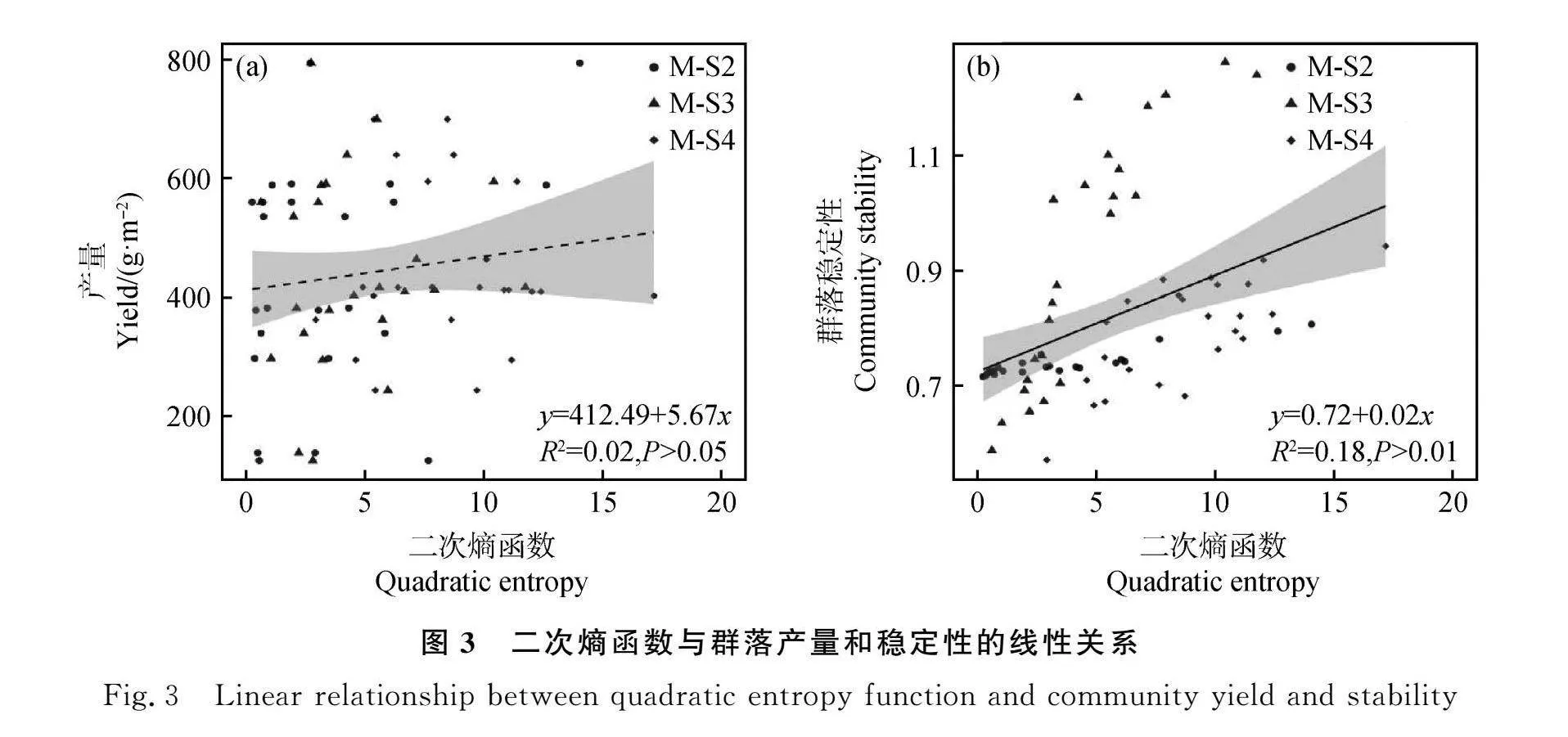

摘要:功能性狀能夠量化植物生理生態特征并預測生態系統功能。為探究功能性狀對鄉土植物混播草地群落產量和穩定性的影響,本研究以鄉土植物羊草(Leymus chinensis)、賴草(Leymus secalinus)、扁蓿豆(Medicago ruthenica)和非鄉土植物紫花苜蓿(Medicago sativa)為試驗材料,分別設置各物種單播以及羊草-扁蓿豆、羊草-扁蓿豆-賴草、羊草-扁蓿豆-賴草-苜蓿混播,比較不同處理下產量和穩定性的差異,并基于性狀加權平均值和功能多樣性指數,探究功能性狀對群落產量及穩定性的影響。結果表明:兩物種和三物種混播產量較低,三物種和四物種群落穩定性在大部分時期都高于兩物種;功能多樣性指數的增大有利于維持群落穩定。光合速率和呼吸速率的提高會對穩定性產生負面影響。綜上所述,混播處理沒有顯著提高產量,兩物種組合穩定性較差,光合速率和呼吸速率以及功能多樣性是影響穩定性變化的關鍵因素。
關鍵詞:鄉土植物;混播;功能性狀;功能多樣性;生態位互補效應
中圖分類號:S543+41""" 文獻標識碼:A"""" 文章編號:1007-0435(2024)08-2357-09
Effects of Functional Traits on Yield and Stability of Native Plant Mixed Sowing Grassland
YU An-ran1,2,3, YANG Yan-ze1,2,3, QIAO Guan-dong1,2,3, ZHANG Jun1,2,3,
DONG Yi-ding1,2,3, HOU Xiang-yang1,2,3, LU Wen-jie1,2,3*
(1. College of Grassland Science, Shanxi Agricultural University, Taigu, Shanxi Province 030801, China; 2. Key Laboratory of Model
Innovation in Forage Production Efficiency, Ministry of Agriculture and Rural Affairs, P. R. China, Taigu, Shanxi Province 030801, China;
3. Shanxi Key Laboratory of Grassland Ecological Protection and Native Grass Germplasm Innovation, Taigu, Shanxi Province 030801, China)
Abstract:Functional traits can be used to quantify the physiological and ecological characteristics of plants and to predict ecosystem functions. To explore the impact of plant functional traits on the yield and stability of native plant mixed sowing grassland,in this study we used native plants Leymus chinensis, Leymus secalinus, Medicago ruthenica,and the non-native plant Medicago sativa as experimental materials. The experiments were set up with monoculture of each species,mixture sowing of Leymus chinensis-Medicago ruthenica,Leymus chinensis-Medicago ruthenica-Leymus secalinus,and Leymus chinensis-Medicago ruthenica-Leymus secalinus-Medicago sativa. The differencesof yield and stability under different treatments were compared. Based on the weighted mean and functional diversity of plant traits the impact of functional traits on community yield and stability were explored. The results showed that two-species and three-species mixture sowing had lower yields. The stability of three-species and four-species communities was higher than two-species communities during most of the periods. The promotion of functional diversity was beneficial for maintaining community stability. The increased of photosynthetic rate and respiration rate had a negative impact on stability. The yield of mixture sowing treatments didn’t significantly increase,and the stability of two-species combination was poor. The key factors influencing stability changes were photosynthetic rate,respiration rate,and functional diversity.
Key words:Native plants;Mixed sowing;Functional traits;Functional diversity;Niche complementary effect
在自然因素和人類活動的影響下,草地植被群落結構發生改變[1],物種多樣性、豐富度和均勻度降低,生態系統結構和功能逐漸退化[2]。恢復退化草地,其根本目標是最終實現草地生態系統的全面恢復,包括生物多樣性、生態過程、生物之間相互作用在內的整個生態系統與生命過程[3]。由多物種組成的近自然群落能夠提升草地的產量、穩定性以及對環境變化的抵抗能力[4]。遵循因地制宜的原則,運用鄉土植物建設人工混播草地,往往能大幅度提升草地的產量和穩定性,促進草地生態功能的提高[5],是實施草地生態保護的有效措施[6]。
“功能性狀”一詞起源于植物群落生態學[7],其值量化了生物體的生態學特征,反映了植物對環境變化響應的生態策略[8]。作為可以預測生態系統功能的框架,這個概念在近年來被廣泛運用[9-10]。目前主要通過質量比假說和生態位補償假說這兩方面來解釋功能性狀對產量和穩定性的影響[11-12]。一方面,強調優勢物種性狀影響植物群落并調節生態系統功能特征[13-15]。Peter等[16]探究產量與優勢種功能性狀的關系,表明群落產量與植株高度呈正相關。另一方面,功能多樣性指數通過量化種間性狀值的分布來評估性狀多樣性[17-18],反映物種資源利用的差異,與“生態位補償假說”相關[19]。Wang等[20]探究喀斯特地區植物功能多樣性與群落穩定性,表明功能多樣性的提高有利于維持群落穩定。Tilman等[19]研究表明隨著物種之間性狀差異的增加,資源利用策略的多樣性也會增加,物種重疊程度減少。功能性狀的差異能夠促進資源的充分利用,同時避免物種間的惡性競爭[21]。在自然生態系統中,植物能夠通過改變其功能性狀如高度、葉面積、干物質含量、種子質量等方式,來適應外界環境條件的改變,這種改變同時也體現了生態系統功能的變化[22]。因此基于功能性狀有助于理解不同物種組合下優勢種對群落功能的影響,以及混播草地資源利用和生態位互補情況。
本研究通過開展鄉土植物田間種植試驗,設置單播及兩物種、三物種和四物種混播組合,對不同試驗處理下鄉土植物群落的功能性狀、產量和穩定性進行定量分析,探索功能性狀與植物群落生產力和穩定性之間的關系,旨在了解功能性狀對鄉土植物混播草地產量和穩定性的調控途徑,以期為退化草地植被恢復提供理論依據。
1 材料與方法
1.1 試驗區概況
試驗地位于山西省右玉縣(39°59.808′N,112°19.660′E),海拔1348 m,年均溫4.7℃,總積溫2600℃·d~3600℃·d。初霜期為9月上、中旬,終霜期在5月初,無霜期100~120 d。年均降雨435 mm,年日照2600~2700 h,全年太陽總輻射量598 KJ·cm-2。該區域是典型的北方農牧交錯帶,屬溫帶大陸性季風氣候,草地類型為溫性山地草原,以賴草(Leymus secalinus)、堿茅(Puccinellia tenuiflora)、羊草((Leymus chinensis)、草地風毛菊(Saussurea amara)、鵝絨委陵菜(Potentilla ansrina)、堿蒿(Artemisia anethifolia)、西伯利亞蓼(Polygonum sibiricum)、蘆葦(Phragmites communis)等植物為主。土壤為淡栗鈣土,土壤pH值約為9.2。
1.2 試驗設計
試驗材料包括鄉土植物扁蓿豆(Medicago ruthenica)、賴草和羊草以及非鄉土植物紫花苜蓿(Medicago sativa)。試驗采用完全隨機區組設計,設置4個單播(S)和3個混播(M)共7個處理,分別為:單播賴草(S-Lc)、單播羊草(S-Ls)、單播扁蓿豆(S-Mr)、單播紫花苜蓿(S-Ms);扁蓿豆-羊草混播(M-S2);扁蓿豆-羊草-賴草混播(M-S3);扁蓿豆-羊草-賴草-紫花苜蓿混播(M-S4);每個種植小區為3 m×3 m,種植行株距30 cm。2022年6月進行穴盤育苗,而后挑選長勢一致的幼苗移栽至樣地。混播處理中,各物種的比例相同且相互毗鄰。每個處理重復8次,共56個試驗小區。
1.3 樣品采集與性狀測定
1.3.1 產量測定 分別于2023年7月15日和8月26日進行刈割并測產。具體做法是:在各試驗小區中隨機放置1 m×1 m的樣方,并將樣方中的所有物種進行刈割,留茬約40 cm。樣方外同樣做刈割處理,兩次刈割位置以及留茬高度基本一致。記錄鮮重后分物種裝入信封,帶回實驗室,在105℃的烘箱中殺青30 min后設置65℃烘干48 h至恒重,單播產量即為干重,混播產量為各物種干重之和。
1.3.2 植物豐度和功能性狀測定 植株數量測定與植物性狀測定同步進行。將1 m×1 m的樣方隨機放在試驗田中,記錄樣方尺內的各物種的數量。
自6月初開始每隔15 d進行一次植物性狀測定,從禾草分蘗期開始至枯黃期時結束,共計6次(T1-T6)。在樣地中選擇每種植物各5株,以測定株高。用便攜式光合儀(CIRAS-4)測定植物光合速率、呼吸速率和水分利用效率。用便攜式植物效率分析儀(Pocket Pea)測定葉綠素含量。選取每種植物的6株個體,取得每株植物的3枚葉片,稱取葉片鮮重,使用千分尺測定葉片厚度,使用葉面積掃描儀(Epson Scan)測定葉片面積,以蒸餾水浸泡法測定葉片飽和含水量,通過烘箱65℃烘干至恒重獲取葉片干重。計算葉片相對含水率、比葉面積和干物質含量:葉片相對含水率=[(葉片鮮重-干重)×(飽和重-干重)]×100%[23],比葉面積=葉面積/葉片干重[24],干物質含量=葉片干重/鮮重[24]。重鉻酸鉀-外加熱法測定葉片碳含量,凱氏定氮法測定氮含量,火焰光度計法測定鉀含量,鉬銻抗比色法測定磷含量。
1.4 數據指標計算與分析
1.4.1 群落性狀加權平均值 群落性狀加權平均值(Community weighted mean trait values)是對群落中每種植物性狀值的加權平均,其中權重由每種植物的豐度來確定,群落優勢種對該值的影響最大[25]。群落中每個性狀的加權平均值(VCWM)計算公式如下[13]:
VCWM=∑Si=1Piti
其中,S是群落中的物種總數,Pi是第i種物種的相對豐度,ti是第i種物種的性狀值。
1.4.2 Rao二次熵函數 Rao二次熵函數(Rao′s quadratic entropy)能夠體現物種間生態位互補程度,其值較高時表明物種間生態位重疊程度較低,對資源的競爭較小。Rao二次熵(FDq)計算公式如下[26]:
FDq=∑S-1i=1∑Sj=i+1dijPiPj
dij=1n∑nk=1(Xik-Xjk)2
Pi和Pj分別為群落中物種i和物種j的個體數目占總個體數的百分比,即物種的相對豐度;dij為物種i和物種j的功能性狀差值絕對值;S為群落的總物種數,n為性狀數,xik為物種i的k性狀值,xjk為物種j的k性狀值。
1.4.3 群落穩定性指數 群落穩定性指數(Community stability index)為物種密度平均值與標準差之比。其比值越高表明群落穩定性越高,表明各物種密度的變異性小于物種間的平均密度變異性[27]。群落穩定性(ISTB)計算公式如下[28]:
ISTB=μσ
其中,μ為樣品中各物種密度的平均值,σ為各物種密度的標準差。
1.5 統計與分析
使用Microsoft Excel 2016對數據進行整理,對數據進行正態分布和方差齊性檢驗,并保證數據基本符合正態分布。調用R軟件中的“FD”包和“fundiversity”包分別計算功能性狀加權均值和功能多樣性指數;采用單因素方差分析比較不同處理下群落產量的差異;采用雙因素方差分析探究物種組合和生長周期及其交互作用對性狀加權平均值、二次熵函數和群落穩定性的影響;使用Pearson相關系數分析不同物種組合中功能性狀與群落產量和穩定性的關系;使用一元線性擬合二次熵函數與群落產量和穩定性的關系;繪圖在R 4.3.1中完成。
2 結果與分析
2.1 不同處理下群落產量和穩定性的差異
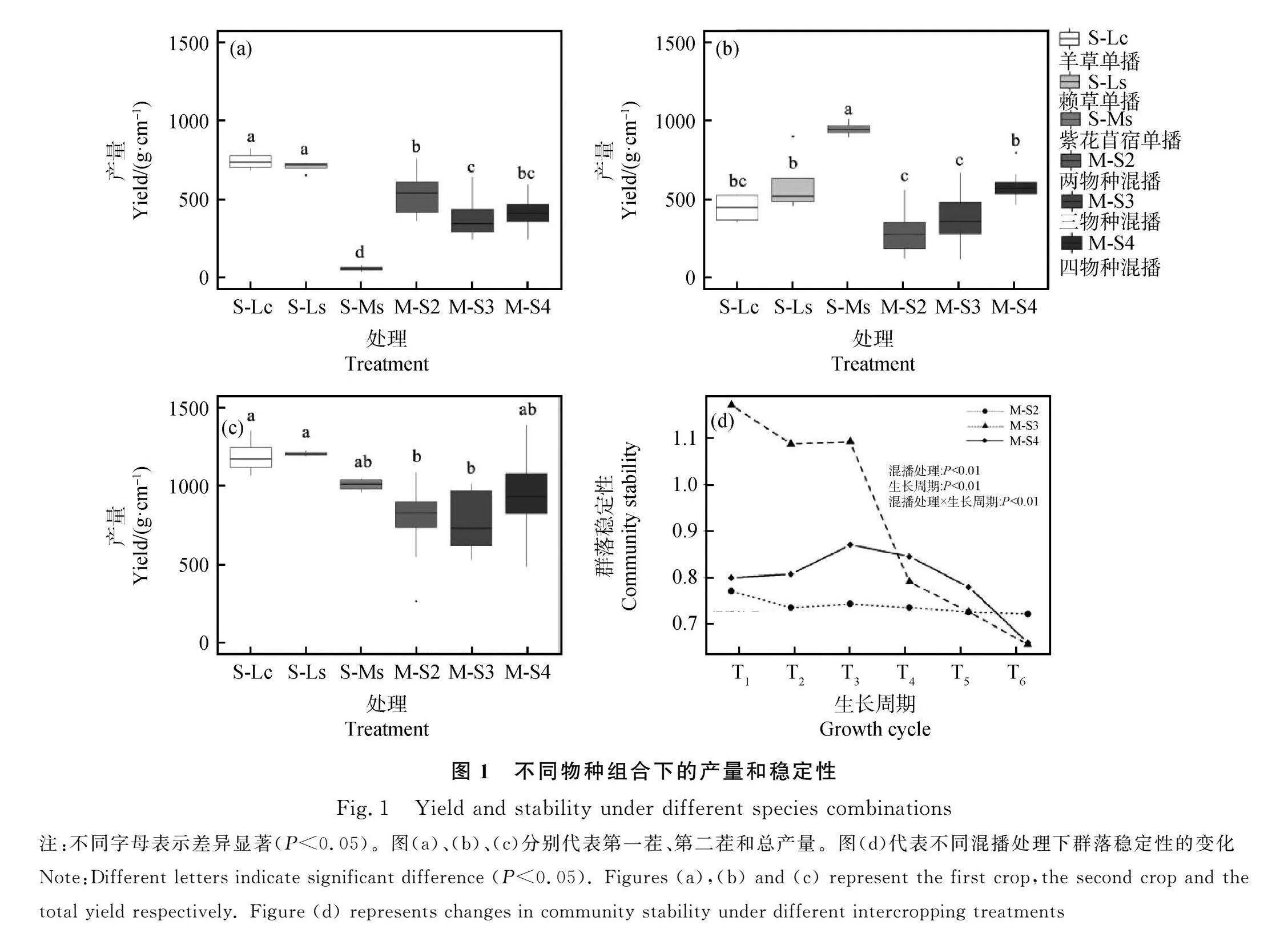
對于第一茬產量,S-Lc和S-Ls產量最高且顯著高于其他處理(P<0.05)。M-S2添加扁蓿豆后,產量相比S-Lc減少了27.6%(P<0.05)。M-S3添加賴草后,相比M-S2產量下降了28.3%(P<0.05),且與S-Ls相比下降了45.2%(P<0.05)。在第一茬的產量中,禾草單播產量顯著高于混播(P<0.05,圖1a)。
在第二茬產量中,S-Ms產量最高且顯著高于其他處理(P<0.05)。M-S2相較于S-Lc處理,產量沒有發生顯著變化。而M-S3在添加賴草后,與M-S2相比產量沒有顯著改變,但與S-Ls相比下降了56.4%(P<0.05)。M-S4相較于M-S3,添加苜蓿后產量提高了53.9%(P<0.05,圖1b)。
通過比較不同物種組合下的總產量,M-S2總產量顯著低于S-Lc(P<0.05),表明在群落中添加扁蓿豆反而使群落產量下降;M-S3添加賴草后與M-S2產量無顯著差異;在加入苜蓿后,M-S4產量相較于M-S3無顯著變化。綜上所述,混播處理并沒有顯著提高群落產量(圖1c)。
混播處理、生長周期及其交互作用對群落穩定性有極顯著的影響(P<0.01)。M-S2在分蘗期(T1)的群落穩定性最高,而后隨著植株生長群落穩定性整體呈下降趨勢。M-S3群落穩定性在分蘗期(T1)時最高,隨后逐漸降低,由開花期到成熟期的過渡階段下降幅度最大,秋末成熟期時到達最低點。M-S4群落穩定性呈先增加后減少的趨勢,在開花時(T3)達到最高,而后逐漸降低。通過對比同一時期下不同混播處理下群落穩定性的差異,除枯黃期外,M-S2群落穩定性在其他時期均低于M-S3和M-S4(P<0.05);M-S3群落穩定性在分蘗期(T1,T2)和開花期(T3)顯著高于其他兩個混播處理(P<0.05),但隨著成熟期的到來,群落穩定性驟然下降,并在成熟時(T4,T5)時低于M-S4處理(P<0.05)。M-S2群落穩定性一直處于較低的水平,在整個生長周期中其變化趨勢相對較小;M-S3和M-S4群落穩定性對植株生長變化的敏感性較強。綜上所述,三物種和四物種混播群落穩定性在大多數時期都高于兩物種(圖1d)。
2.2 性狀加權均值與群落產量和穩定性的相關性
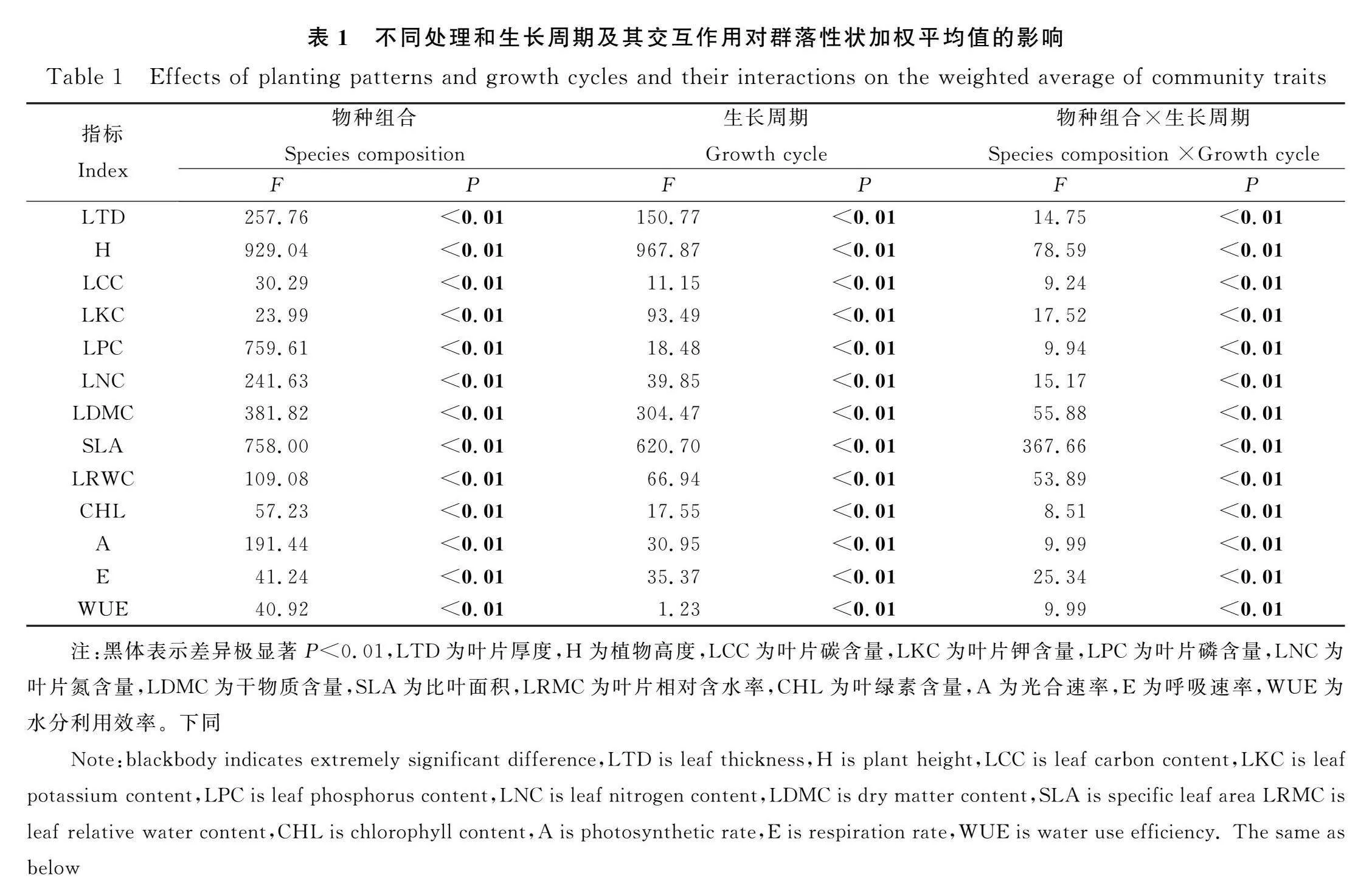
物種組合對性狀加權均值有極顯著的影響(P<0.01)。在不同生長周期下,性狀加權均值存在極顯著的差異(P<0.01)。物種組合和生長周期的交互作用對性狀加權均值有極顯著的影響(P<0.01,表1)。
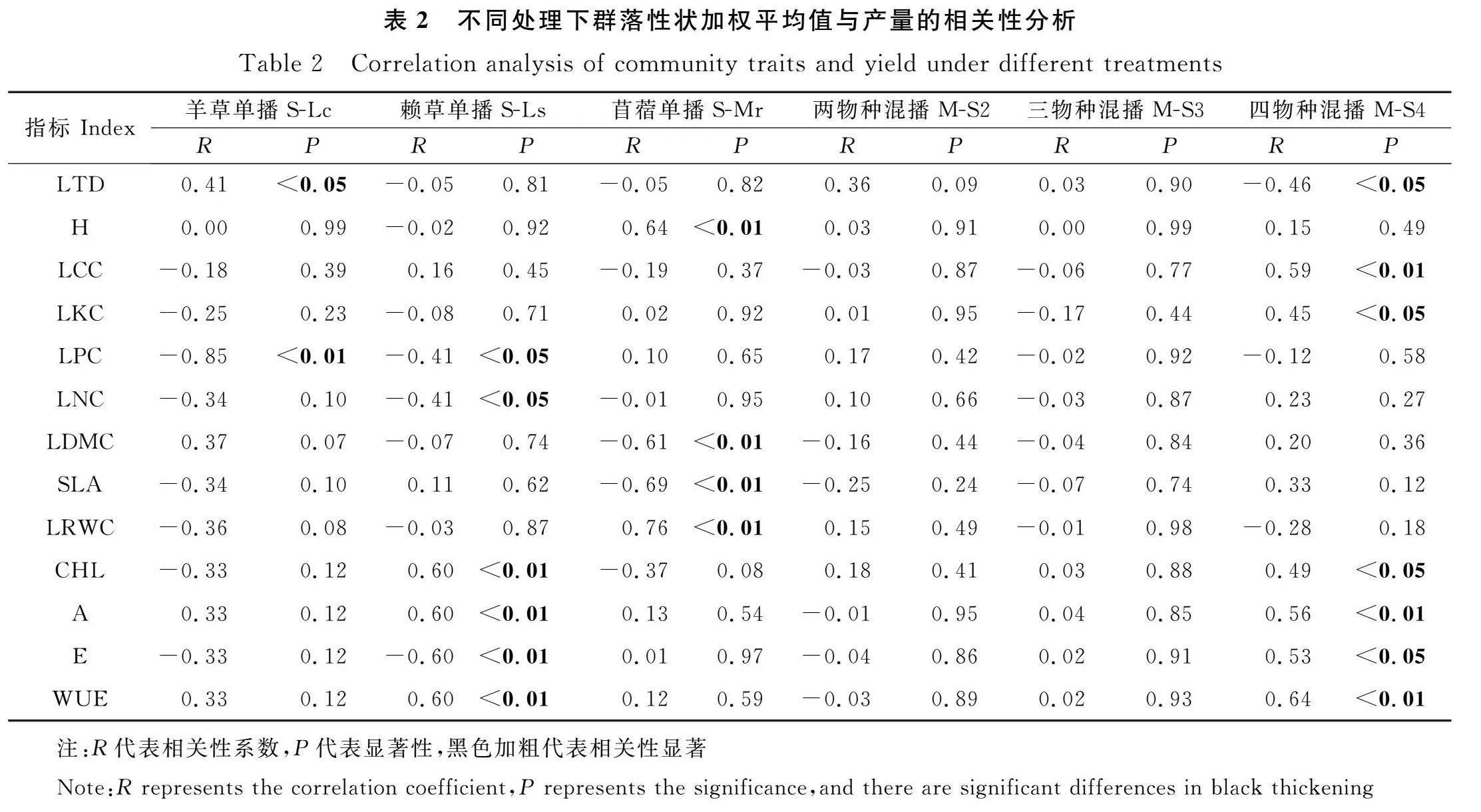
在S-Lc處理中,群落產量與葉片厚度呈顯著正相關(P<0.05),與葉片磷含量呈極顯著負相關(P<0.01)。在S-Ls處理中,群落產量與葉片磷含量和葉片氮含量呈顯著負相關(P<0.05),與呼吸速率呈極顯著負相關,與葉綠素含量、光合速率和水分利用效率呈極顯著正相關(P<0.01)。在S-Mr處理中,群落產量與植株高度和葉片相對含水率呈極顯著正相關(P<0.01),與干物質含量和比葉面積呈極顯著負相關(P<0.01)。在M-S2和M-S3處理中,群落產量與功能性狀無顯著相關性。在M-S4處理中,群落產量與葉片厚度呈顯著負相關(P<0.05),與葉片鉀含量、葉綠素含量和呼吸速率呈顯著正相關(P<0.05),與葉片碳含量、光合速率和水分利用效率呈極顯著正相關(P<0.01,表2)。
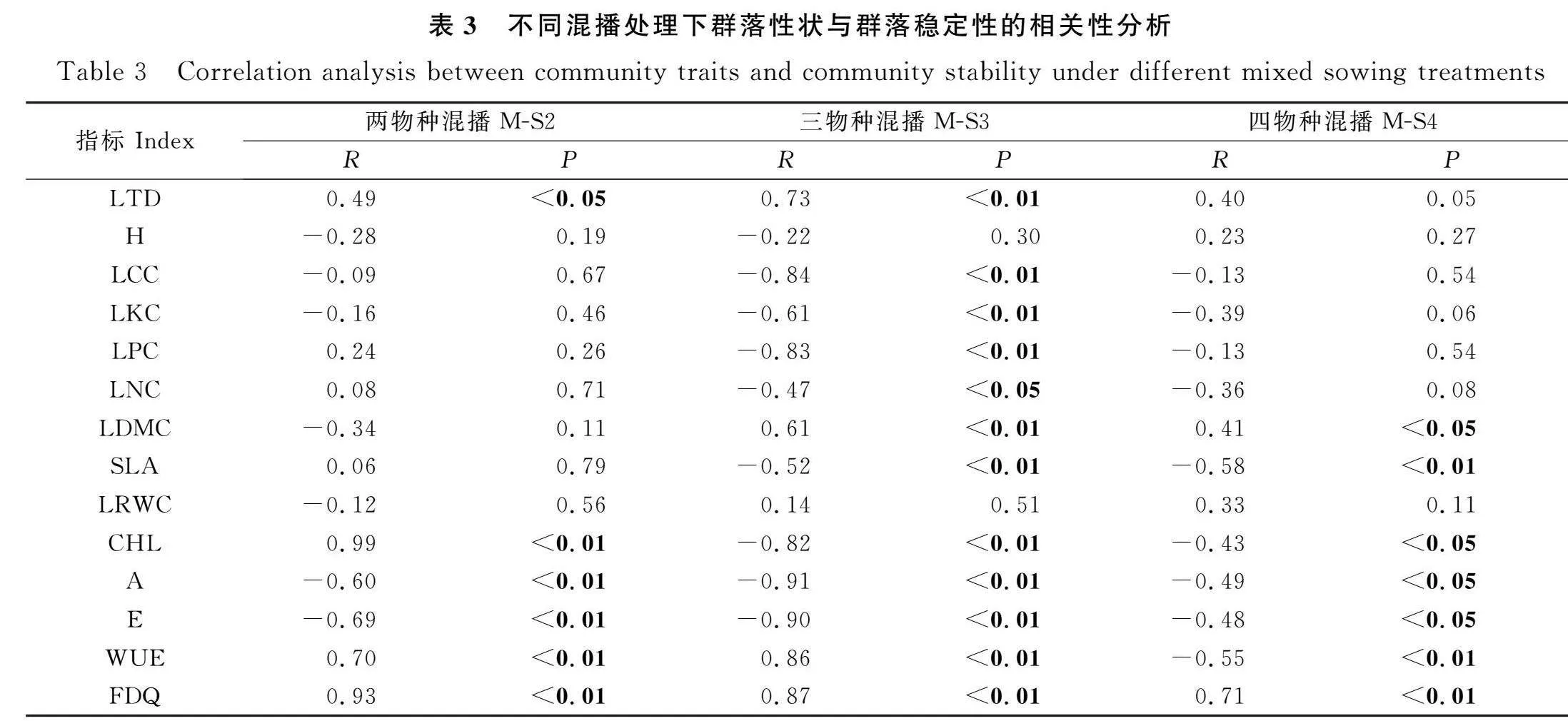
群落穩定性與葉片厚度和Rao二次熵函數呈正相關,光合速率和呼吸速率與穩定性顯著負相關。在M-S2中,群落穩定性與葉綠素含量呈極顯著正相關(P<0.01),與水分利用效率呈極顯著負相關。在M-S3中,群落穩定性與葉片碳、鉀、磷、葉綠素含量以及比葉面積呈極顯著負相關(P<0.01),與氮含量呈顯著負相關(P<0.05),與水分利用效率呈極顯著正相關。在M-S4中,群落穩定性與干物質含量呈顯著正相關(P<0.05),與葉綠素含量呈顯著負相關,與比葉面積和水分利用效率呈極顯著負相關(P<0.01,表3)。
2.3 功能多樣性與群落產量和穩定性的關系
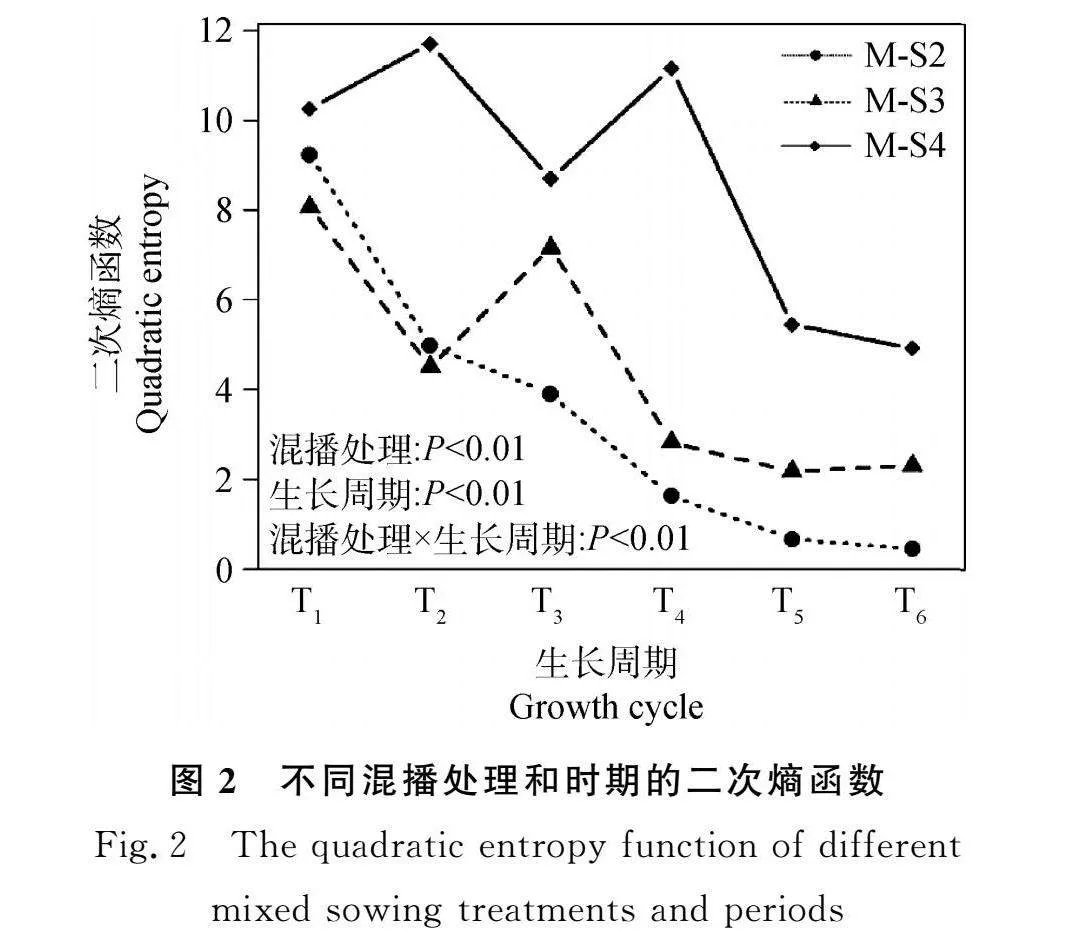
混播模式、生長周期及其交互作用均對功能多樣性指數有極顯著影響(P<0.01)。分蘗期時(T1,T2),M-S2功能多樣性高于M-S3處理,同時又低于M-S4。而后植株顯蕾,混播處理的性狀互補差異發生了變化,M-S3處理的功能多樣性高于M-S2。大部分時期性狀互補程度為M-S4>M-S3>M-S2。
在整個生長季內,M-S2功能多樣性呈逐漸降低的趨勢,且在T1-T2的生長期內下降的趨勢最大。M-S3功能多樣性呈先下降后上升而后再下降的趨勢,從分蘗期(T2)到開花期(T3)的過渡階段逐漸上升,隨著開花期結束再次呈下降趨勢。M-S4功能多樣性出現兩個峰值,分別是在春末分蘗期(T2)以及秋初成熟期(T4),除了這兩個時期外,其余都呈下降趨勢(圖2)。
通過線性擬合,Rao二次熵函數與混播群落總產量無顯著線性關系(R2=0.02,P>0.05),群落總產量會隨著性狀互補程度的增加呈上升趨勢,但線性關系并不顯著(圖3a)。Rao二次熵函數與群落穩定性有顯著線性關系(R2=0.18,P<0.01),性狀互補程度的增大有利于群落穩定性的維持(圖3b)。
3 討論
3.1 不同物種組合的群落產量和穩定性
群落產量對物種多樣性的響應機制主要來自于優勢種效應和生態位互補效應[29]。本研究中,M-S2與S-Lc相比,扁蓿豆的添加未能顯著提高群落總產量。扁蓿豆呈半匍匐式生長[30],會遮蔽部分光線,限制其周圍低層禾草生長,導致群落產量下降。與M-S2相比,賴草的添加并沒有顯著改變群落總產量。與M-S3相比,苜蓿添加后產量有上升的趨勢。來幸樑等[31]探究苜蓿與禾草混播的土壤養分特征,表明紫花苜蓿對養分積累有促進作用。產量與葉片碳含量和鉀含量呈正相關。間接表明在加入苜蓿后,葉片碳含量和鉀含量有所提升,M-S4產量略高于M-S3。由單一物種到四物種變化的過程中,產量并未顯著提高。由此可見,物種變化會對群落產量產生影響[4],但混播物種數量增加并不一定會提升群落產量[32]。混播中各物種性狀存在差異且呈現出互補效應,但功能多樣性與產量無密切關系,在本研究中生態位互補效應不能解釋產量變化。
物種多樣性變化、生態位分化以及特定環境波動下的生物量變化都會引起群落穩定性改變[33]。本研究中,混播的群落穩定性隨著植株生長整體呈下降趨勢,是因為前期群落密度較低資源競爭相對較少,但隨著群落的發展競爭會逐漸加劇,導致群落穩定性下降[34]。M-S3穩定性在大部分時期都高于M-S2,表明賴草的添加有利于提高群落穩定性。M-S3穩定性在禾草開花后迅速下降,并低于M-S4,其功能多樣性同樣自開花后下降,表明在此期間群落生態位重疊程度較大,資源競爭加劇,導致群落穩定性驟降。Dietrich等[35]探究不同物種組合對多年生混播草地的影響,也表明物種多樣性變化影響著群落穩定性。羊草和賴草都具備無性繁殖的能力[36],它們的繁殖數量往往隨著生長期的變化而產生波動。在秋末種子成熟后兩物種群落穩定性逐漸高于其他混播處理,這一時期為正是羊草擴大種群規模和生長空間的最重要時期[37],羊草擴繁迅速,在這個過程中賴草和羊草爭奪空間資源,同時賴草生長受到限制,導致三物種和四物種群落穩定性下降。
3.2 性狀加權均值與群落產量和穩定性
性狀加權平均值通常反映了優勢物種對群落的作用,側重解釋優勢物種功能性狀對生態系統過程的影響[14]。在本試驗中,生長周期和物種組成的變化均對性狀加權平均值產生了顯著影響,表明在不同的組合中的物種資源獲取以及向不同生理組織的投入存在著差異[38],在整個生命周期中植物的生長策略并不固定,而是依賴其生長軌跡的變化[39]。Peter等[16]探究物種多樣性與產量關系,表明性狀加權平均值會隨著物種組成的變化而改變,這與本研究結果一致。在M-S2和M-S3處理中,性狀加權平均值和功能多樣性指數都與產量無相關性,在試驗過程中性狀測量傾向于植株葉片,但根系性狀也屬生長經濟型性狀[40],未被測量的根系性狀同樣影響產量變化。Louise等[41]探究干旱條件下根系性狀與產量的關系,表明比根長和根長密度是驅動產量變化的重要因素。因此還需要進一步的研究。在添加了苜蓿后,M-S4群落產量與葉片碳含量、葉片鉀含量、葉綠素含量、光合速率、呼吸速率和水分利用效率呈顯著正相關,與葉片厚度呈顯著負相關。葉片厚度反映了水分供給對植物的限制[42];葉片鉀含量和磷含量反映了養分利用情況[43];葉綠素含量、光合速率、呼吸速率和水分利用效率則與光合作用密切相關[44]。光合作用的增強有利于提升產量[45]。因此,四物種組合中葉片厚度和光合性狀是影響產量的關鍵因素。
性狀加權平均值還對群落穩定性的變化有所貢獻,葉片厚度與穩定性呈正相關。葉片厚度反映了植物適應干旱環境的能力[46],葉片厚度的增加有利于半干旱環境中群落穩定性的維持。在本試驗中光合速率和呼吸速率與群落穩定性呈負相關。植株光合作用的增強能夠提高生長速率[47],而以保守策略相關性狀為主的群落(“慢速生長”)比快速生長群落更能抵抗干擾[48],因此光合速率和呼吸速率的增長不利于促進群落穩定。性狀與穩定性的關系在不同混播模式中各有不同。在M-S3和M-S4中,葉綠素含量與穩定性呈負相關,但在M-S2中,葉綠素含量與穩定性的關系截然相反。功能性狀與群落功能的關系較為復雜,不同群落中的關系往往存在著差異[49]。
3.3 功能多樣性與群落產量和穩定性
除了優勢物種的性狀外,功能多樣性也是群落功能的重要驅動因素[50],反映了混播組合的種間互補效應和資源利用效率[51]。本研究中,與M-S2相比,添加賴草促進了功能多樣性的提高;與M-S3相比,添加苜蓿進一步提高了群落性狀互補程度。表明功能多樣性會隨著物種增加而呈現上升趨勢[14]。隨著生長周期的推移,性狀間互補程度也發生變化。羊草和賴草的分蘗能力影響著空間生態位分化,進而使功能多樣性指數發生變化。在三個混播處理中,功能多樣性指數始終與穩定性呈正相關,并且群落穩定性隨著性狀間互補程度的增加而提高。Wood等[52]探究功能多樣性與穩定性的關系,也表明功能多樣性有利于維持群落穩定,這與本研究結果一致。但功能多樣性與群落產量并無顯著相關性。雖然產量會隨著功能多樣性的增加有所提高,但它們之間的線性關系并不顯著,表明互補效應并不能顯著預測產量變化。Weisser等[53]探究物種多樣性對生態系統功能的影響,表明群落中的優勢種性狀在解釋群落產量時比功能多樣性更重要。
植物功性狀間的差異能夠促進資源互補利用[54],物種對資源的利用不同,避免了過度競爭,保證群落穩定性的維持[21]。本試驗中葉綠素含量、光合速率、呼吸速率以及水分利用效率與群落穩定性相關,同時穩定性會隨著功能多樣性的增大而增加,表明群落優勢物種的功能性狀能夠調節植物群落的穩定性[55],同時互補性也可以在環境因素的干擾下發揮穩定群落的作用[21]。優勢種效應和互補效應共同解釋了群落穩定性的維持機制[56]。
4 結論
兩物種(羊草-扁蓿豆)和三物種(羊草-扁蓿豆-賴草)混播產量低于單播,兩物種群落穩定性低于三物種和四物種混播。就產量和穩定性而言,兩物種混播效果較不理想。光合速率、呼吸速率和水分利用效率是影響穩定性變化的重要因素。功能多樣性的增大有利于促進群落穩定性的維持。在建植混播草地的過程中,建議關注群落光合性狀以及性狀差異性,以發揮鄉土植物混播優勢。
參考文獻
[1] 童永尚,張春平,俞旸,等. 多年生人工混播草地暫穩態概念及維持技術解析[J]. 草地學報,2022,30(11):2845-2855
[2] 魏晶晶,秦瑞敏,張中華,等. 不同退化程度高寒草地植物群落與土壤性質變化及相關性分析[J]. 草地學報,2022,30(11):3035-3045
[3] 賀金生. 恢復退化草地,不能只是增加綠度[N]. 光明日報.2024-03-23(1)
[4] JORGENSEN M,BAKKEN A K,OSTREM L,et al. The effects of functional trait diversity on productivity of grass-legume swards across multiple sites and two levels of nitrogen fertiliser[J]. European Journal of Agronomy,2023,151:126993
[5] 王麗霞,史園莉,張宏偉,等. 2000-2020年北方農牧交錯區植被生態功能變化及驅動因子分析[J]. 生態環境學報,2021,30(10):1990-1998
[6] 丁子健,劉文,李天澤,等. 鄉土草草地建植對遼西北連作農田土壤的改良效果研究[J]. 草地學報,2024,32(5):1572-1582
[7] LAVOREL S,GARNIER E. Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail[J]. Functional Ecology,2002,16(5):545-556
[8] MOUILLOT D,GRAHAM N A,VILLEGER S,et al. A functional approach reveals community responses to disturbances[J]. Trends in Ecology amp; Evolution,2013,28(3):167-177
[9] GLADSTONE G R V,PILDITCH C A,STEPHENSON F,et al. Linking Traits across Ecological Scales Determines Functional Resilience[J]. Trends in Ecology amp; Evolution,2019,34(12):1080-1091
[10]LIU C,LI Y,YAN P,et al. How to Improve the Predictions of Plant Functional Traits on Ecosystem Functioning?[J]. Frontiers in Plant Science,2021,12:1-5
[11]SASAKI T,LAUENROTH W K. Dominant species,rather than diversity,regulates temporal stability of plant communities[J]. Oecologia,2011,166:761-768
[12]CRAINE J M,TILMAN D,WEDIN D,et al. Functional traits,productivity and effects on nitrogen cycling of 33 grassland species[J]. Functional Ecology,2002,16(5):563-574
[13]DIAZ S,LAVOREL S,BELLO F D,et al. Incorporating plant functional diversity effects in ecosystem service assessments[J]. Proceedings of the National Academy of Sciences,2007,104(52):20684-20689
[14]CADOTTE M W. Functional traits explain ecosystem function through opposing mechanisms[J]. Ecology Letters,2017,20(8):989-996
[15]HILLEBRAND H,BENNETT D M,CADOTTE M W. Consequences of dominance:a review of evenness effects on local and regional ecosystem processes[J]. Ecology,2008,89(6):1510-1520
[16]DIETRICH P,EISENHAUER N,ROSCHER C. Linking plant diversity productivity relationships to plant functional traits of dominant species and changes in soil properties in 15 year old experimental grasslands[J]. Ecology and Evolution,2023,13(3):e9883
[17]MOUCHET M A,VILLEGER S,MASON N W H,et al. Functional diversity measures:an overview of their redundancy and their ability to discriminate community assembly rules[J].Functional Ecology,2010,24(4):867-876
[18]PETCHEY O L,GASTON K J. Functional diversity:back to basics and looking forward[J]. Ecology Letters,2006,9(6):741-758
[19]TILMAN D,REICH P B,KNOPS J,et al. Diversity and Productivity in a Long-Term Grassland Experiment[J]. Science,2001,294(5543):843-845
[20]WANG Y,CHEN J,ZHANG L,et al. Relationship between diversity and stability of a karst plant community[J]. Ecology and Evolution,2022,12(8):e9254
[21]BROOKER R W,HAWES C,IANNETTA P P M,et al. Plant diversity and ecological intensification in crop production systems[J]. Journal of Plant Ecology,2023,16(6):rtad015
[22]ALBERT C H,BELLO F D,BOULANGEAT I,et al. On the importance of intraspecific variability for the quantification of functional diversity[J]. Oikos,2011(1),121:116-126
[23]ARNDT S K,IRAWAN A,SANDERS G J. Apoplastic water fraction and rehydration techniques introduce significant errors in measurements of relative water content and osmotic potential in plant leaves[J]. Physiologia Plantarum,2015,155(4):355-368
[24]GARNIER E,SHIPLEY B,ROUMET C,et al. A standardized protocol for the determination of specific leaf area and leaf dry matter content[J]. Functional Ecology,2001,15:688-695
[25]ENGEL T,BRUELHEIDE H,HOSS D. Traits of dominant plant species drive normalized difference vegetation index in grasslands globally[J]. Global Ecology and Biogeography,2023,32(5):695-706
[26]RAO CR. Quadratic entropy and analysis of diversity[J]. Sankhya A,2010,72:70-80
[27]LEHMAN C,TILMAN D. Biodiversity,Stability,and Productivity in Competitive Communities[J]. The American Naturalist,2000,156(5):534-552
[28]WANG Y,CHEN J,ZHANG L,et al. Relationship between diversity and stability of a karst plant community[J]. Ecology and Evolution,2022,12(8):e9254
[29]ROSCHER C,SCHUMACHER J,LIPOWSKY A,et al. A functional trait-based approach to understand community assembly and diversity-productivity relationships over 7 years in experimental grasslands[J]. Perspectives in Plant Ecology,Evolution and Systematics,2013,15(3):139-149
[30]張雨桐. 扁蓿豆株型特征及其直立莖調控機制解析[D]. 呼和浩特:內蒙古農業大學,2022,13-14
[31]來幸樑,師尚禮,吳芳,等. 紫花苜蓿與3種多年生禾草混播草地的土壤養分特征[J]. 草業科學,2020,37(1):52-64
[32]SANDERSON M A. Stability of production and plant species diversity in managed grasslands:A retrospective study[J]. Basic and Applied Ecology,2010,11(3):216-224
[33]HECTOR A,HAUTIER Y,SANER P,et al. General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding[J]. Ecology,2010,91(8):2213-2220
[34]XIAO J,LI Y,MEINERS SJ,et al. Joint effects of resource supply and resource types on properties of population dynamic models[J]. Journal of Ecology,2023,112(2):360-373
[35]DIETRICH P,EBELING A,MEYER ST,et al. Plant diversity and community age stabilize ecosystem multifunctionality[J]. Global Change Biology,2024,30(3):e17225
[36]張兆軍. 松嫩平原羊草莖頂分生組織分化研究[D]. 長春:東北師范大學,2006,2-3
[37]白烏云. 羊草根莖克隆生長特性種內分化及與原生境地理和氣候因素的關聯研究[D]. 北京:中國農業科學院,2021,94-95
[38]FALSTER D S,DUURSMA R A,FITZJOHN R G. How functional traits influence plant growth and shade tolerance across the life cycle[J]. Proceedings of the National Academy of Sciences,2018,115(29):E6789-6798
[39]GIBERT A,GRAY E F,WESTOBY M,et al. On the link between functional traits and growth rate:meta-analysis shows effects change with plant size,as predicted[J]. Journal of Ecology,2016,104(5):1488-1503
[40]FRESCHET G T,ROUMET C,COMAS L H. Root traits as drivers of plant and ecosystem functioning:current understanding,pitfalls and future research needs[J]. New Phytologist,2021,232(3):1123-1158
[41]COMAS L H,BECKER S R,CRUZVON M V,et al.. Root traits contributing to plant productivity under drought[J]. Frontiers in Plant Science,2013,4:442
[42]張曉飛. 秦嶺兩種櫟屬植物葉性狀及其關系沿海拔梯度的變化[D].西安:西北大學,2012,3-4
[43]HOMOLOVA L,MALENOVSKY Z,CLEVERS J G,et al. Review of optical-based remote sensing for plant trait mapping[J]. Ecological Complexity,2013,15:1-16
[44]PAVLOVIC D,NIKOLIC B,DJUROVIC S,et al. Chlorophyll as a measure of plant health:Agroecological aspects[J]. Pesticidi I Fitomedicina,2014,29(1):21-34
[45]ZHU X,LONG S P,ORT D R. Improving photosynthetic efficiency for greater yield[J]. Annual Review of Plant Biology,2010,61:235-261
[46]AFZAL A,DUIKER S W,WATSON J E. Leaf thickness to predict plant water status[J]. Biosystems Engineering,2017,156:148-156
[47]KIRSCHBAUM M U. Does Enhanced Photosynthesis Enhance Growth? Lessons Learned from CO2 Enrichment Studies[J]. Plant Physiology,2010,155(1):117-124
[48]BELLO F D,LAVOREL S,HALLETT L M,et al. Functional trait effects on ecosystem stability:assembling the jigsaw puzzle[J]. Trends in Ecology amp; Evolution,2021,36(9):822-836
[49]何念鵬,劉聰聰,張佳慧,等. 植物性狀研究的機遇與挑戰:從器官到群落[J]. 生態學報,2018,38(19):6787-6796
[50]ENQUIST B J,NORBERG J,BONSER S P,et al. Scaling from traits to ecosystems:developing a general trait driver theory via integrating trait-based and metabolic scaling theories[J]. Advances in Ecological Research,2015,52:249-318
[51]PALACIOS P G,GROSS N,GAITAN J,et al. Climate mediates the biodiversity-ecosystem stability relationship globally[J]. Proceedings of the National Academy of Sciences,2018,115(33):8400-8405
[52]WOOD C M,MCKINNEY S T,LOFTIN C S. Intraspecific functional diversity of common species enhances community stability[J]. Ecology and Evolution,2017,7(5):1553-1560
[53]WEISSER W W,ROSCHER C,MEYER S T. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment:Patterns,mechanisms,and open questions[J]. Basic and Applied Ecology,2017,23:1-73
[54]JOCHUM M,FISCHER M,ISBELL F,et al. The results of biodiversity-ecosystem functioning experiments are realistic[J]. Nature Ecology amp; Evolution,2020,49(11):1485-1494
[55]LUO W,SHI Y,WILKINS K,et al. Plant traits modulate grassland stability during drought and post-drought periods[J]. Functional Ecology,2023,37(10):2611-2620
[56]WU J,BAO X,ZHANG J,et al. Temporal stability of productivity is associated with complementarity and competitive intensities in intercropping[J]. Ecological Applications,2022,33(1):e2731
(責任編輯 付 宸)
