菊苣CiGL2基因克隆及功能分析
2024-12-31 00:00:00毛佳靜陸爽費星宇杜洋許立新
草地學報 2024年8期

摘要:菊苣(Cichorium intybus L.),菊科菊苣屬多年生草本生植物,是一種新型的牧草。菊苣葉片具有表皮毛,表皮毛可能影響其作為牧草的飼用價值。GLABRA 2(GL2)是植物轉錄因子,有研究顯示其可能參與植物表皮毛的發育。本研究克隆了菊苣的GL2基因(CiGL2),并通過遺傳轉化手段初步分析和驗證該基因的功能。CiGL2開放閱讀框為2208 bp,編碼一個735個氨基酸的轉錄因子。進化分析表明,菊苣與生菜(Lactuca sativa L.)的親緣關系較近;亞細胞定位顯示,該蛋白定位于細胞核內;過表達植株葉片邊緣、葉腹表皮毛密度較高。利用CRISPR/Cas9基因編輯技術獲得菊苣CiGL2突變植株,與對照普通菊苣株系相比,突變植株葉腹面、葉片背面表皮毛長度較短,表皮毛密度下降。研究結果說明菊苣CiGL2基因功能可能與菊苣表皮毛發育相關。本研究為菊苣GL2基因功能分析提供理論基礎,也為菊苣牧草品質的改良提供了創新種質資源。
關鍵詞:菊苣;表皮毛;CiGL2;CRISPR/Cas9
中圖分類號:Q344+.13""" 文獻標識碼:A"""" 文章編號:1007-0435(2024)08-2366-08
Clone and Functional Analysis of CiGL2 Gene in Cichorium intybus
MAO Jia-jing, LU Shuang, FEI Xing-yu, DU Yang, XU Li-xin*
(School of Grassland Science, Beijing Forestry University, Beijing 100083, China)
Abstract:Cichorium intybus is a perennial herbaceous plant of the genus Cichorium,family Asteraceae and can be used as forage. The development of epidermal hair on chicory leaf may affect feeding value as well as forage quality. It has been documented that the GLABRA 2 (GL2) gene family may regulate the development of epidermal hair and trichome through different mechanisms. In this study,the CiGL2 gene was cloned,and the function of the gene was further analyzed. The open reading frame of CiGL2 was 2208 bp. Evolutionary analysis showed that chicory was more closely related to lettuce (Lactuca sativa var. ramosa). Subcellular localization test showed that the protein was localized in the nucleus. Leaves of over expressed plants showed an increase in the density of epidermal hair at the leaf margin and on the ventral surface of the leaf. Chicory GL2 mutant plants were obtained through CRISPR/Cas9 gene editing,and the length of epidermal hair on the ventral surface of the leaf was shorter and the density of epidermal hair decreased compared with that of the wild-type plants. This indicated that CiGL2 was associated with epidermal hair development in chicory. This study provided a theoretical basis for the functional study of the GL2 gene,and the mutant lines obtained could be important germplasm for chicory improvement.
Key words:Cichorium intybus;Epidermal hairs;CiGL2;CRISPR/Cas9
菊苣(Cichorium intybus L.) 是菊科菊苣屬多年生草本植物,廣泛分布在全球的溫帶和熱帶地區[1],多見于陽光充足的環境。菊苣因具有栽培簡易、葉質鮮嫩、適口性較好、產量較高等特點可作為飼用牧草,被認為是最有發展潛力的經濟作物之一[2],一些品種也可作為食用菜[3]。
植物表皮毛是植物表皮細胞發育形成的一種具有特殊結構的毛狀附屬物,不同類型的表皮毛在植物中行使的功能也不同[4]。表皮毛主要分為兩種類型:腺毛狀體和無腺毛狀體[5],有腺毛狀體的表皮毛可以產生有害物質,如酚類化合物,揮發性油脂等,導致有害生物因中毒而死亡[6];無腺毛狀體在預防生物侵害和機械損傷等方面發揮著同等重要的作用[7]。菊苣葉片表皮毛對動物的進食和消化可能產生一定的影響[8]。
GLABRA 2(GL2)屬于同源結構域亮氨酸拉鏈(HD-ZIP)IV類TF家族,其具有植物特異性結構域排列[9],是表皮分化的關鍵調節因子[10]。研究顯示擬南芥(Arabidopsis thaliana (L.) Heynh.) GLEBRA2(GL2)基因可能和表皮毛發育相關[11]。GL2基因的表達貫穿在整個表皮毛發育過程,并在發育的表皮毛中高水平表達[12]。在根表皮中,GL2促進無毛細胞分化;在葉表皮中,它很可能參與毛狀體的形成和分化[13];此外GL2可能參與種皮黏液的產生[11]。黃瓜(Cucumis sativus L)果刺為多細胞表皮毛,CsGL2參與調控黃瓜果刺(表皮毛)的分化形成[14]。此外,GL2基因也被證明參與芽表皮的發育和植物花青素的合成[15]。草類植物的GL2基因家族研究較少。
本研究從菊苣優質牧草品種‘將軍’中克隆得到CiGL2基因,利用遺傳轉化手段、基因編輯技術對其進行功能驗證和分析。該基因的克隆及功能研究,將為闡明菊苣表皮毛形成機制提供依據和參考,獲得的轉基因株系為改良菊苣牧草品質提供可利用的創新種質。
1 材料與方法
1.1 植物材料及試劑
菊苣牧草品種‘將軍’(‘K042’),購自黃家百綠公司。‘普那菊苣’由貴州草業所惠贈。‘結球菊苣’、擬南芥gl2-3突變體為本課題組前期保存。溫室培養條件25℃,光照16 h/20℃黑暗8 h,濕度60%。土壤為珍珠巖∶草炭灰∶蛭石=1∶4∶1。
基因克隆載體為pTOPO001 Blunt Simple Cloning Kit,菌株為Efficom 5α,購自于北京金沙;農桿菌株為EHA105(北京GenStar)。培養基為LB培養基,配方為胰蛋白(Tryptone)10 g·L-1,酵母提取物為Yeast extract 5 g·L-1,NaCl 10 g·L-1。熒光定量PCR試劑盒、反轉錄試劑盒等均購自諾唯贊生物科技股份有限公司,測序由北京睿博興科生物技術有限公司完成。
1.2 試驗方法
1.2.1 菊苣葉片組織總RNA提取及cDNA合成 參照Total試劑盒說明提取‘K042’菊苣葉片總RNA,-80℃冰箱保存。使用反轉錄試劑盒將RNA進一步反轉錄合成為cDNA。
1.2.2 CiGL2基因克隆 根據擬南芥AtGL2CDS序列,經BLAST比對,從‘普那’菊苣基因數據庫[16]中獲得KAI3766631.1同源基因,以特異性引物CiGL2-F1/R1(表1)和菊苣cDNA為模板克隆CiGL2基因。PCR總反應體系(50 μL)為:cDNA模板1 μL,上下游引物各2.5 μL,DMSO1 μL,dd H2O 18 μL。PCR反應程序為:98℃預變性30 s;98℃變性30 s,56℃退火5 s,72℃延伸30 s,28~35個循環;72℃延伸1 min,4℃保存。電泳檢測PCR產物,將回收產物連接在pTOPO001 Blunt Simple Vector載體,連接產物轉到Efficom 5α Chemically Competent Cell中,涂板,挑取白色單菌落PCR。PCR總反應體系(20 μL)為:T5 10 μL,上下游引物各為1 μL,菌液為2 μL,ddH2O為6 μL。PCR反應程序為:98℃預變性5 min;98℃變性10 s,56℃退火10 s,72℃延伸30 s,33個循環;72℃延伸1 min。經測序驗證,獲得CiGL2基因的cDNA全長序列。
1.2.3 生物信息學分析 NCBI(http://www.ncbi.nlm.nih.gov/)數據庫BLAST(http://blast.ncbi.nlm.nih.gov/)比對CiGL2同源序列;DNAMAN進行氨基酸序列對比,同源性分析;利用Pfamscan (https://www.ebi.ac.uk/Tools/pfa/pfamscan/)以及InterProScan (http://www.ebi.ac.uk/interpro/)做蛋白質結構域預測和蛋白三級結構預測;使用MEME (http://meme-suite.org/tools/meme)構建系統發育進化樹。
1.2.4 亞細胞定位 參考張建福等人[17]方法,混勻原生質體后24℃培養12~16 h,制作玻片,用于激光共聚焦顯微鏡觀察Leica SP-5(Leisa,Germany)鑒定。
1.2.5 實時熒光定量PCR檢測 分別提取菊苣根、莖、葉、幼根以及幼葉樣品總RNA,反轉錄cDNA,進行實時熒光定量PCR。根據IDT(Integrated DNATechnologies)設計RT-qPCR特異性引物CiGL2-F2/R2和內參基因引物18S-rRNA-F/R,使用熒光定量PCR儀(Roche,Switzerland),體系參考夏曾潤[18],2-ΔΔCt法計算CiGL2基因相對表達量。
1.2.6 擬南芥gl2-3突變體恢復系的獲得 CiGL2基因特異性引物CiGL2-F4/R4,HindIII和BamH I雙酶切,PCR擴增;轉化至Efficom 5α Chemically,回收酶切產物進行體外連接,篩選陽性單克隆進行菌液PCR,將擴增產物送公司測序,構建植物表達載體。質粒轉化農桿菌GV3101,利用農桿菌介導的花序侵染法侵染擬南芥,黑暗條件下培養6~8 h后光照培養;按需二次侵染。
1.2.7 CRISPR/Cas9基因編輯技術獲得突變菊苣植株 在線軟件(http://crispor.tefor.net/)設計sgRNA(small guide RNA,sgRNA)序列,命名為sgRNA1、sgRNA2、sgRNA3,T4連接酶連接入Bbs I酶切sgRNA表達框載體后測序驗證。PTG::CiGL2載體轉化農桿菌EHA105。利用農桿菌介導法侵染普那菊苣,以引物CiGL2-F3/R3篩選抗性植株并測序驗證。
1.2.8 統計分析 2-ΔΔCt法計算分析CiGL2基因相對表達量,采用Excel 2010整理分析數據,Origin 2022作圖。
2 結果與分析
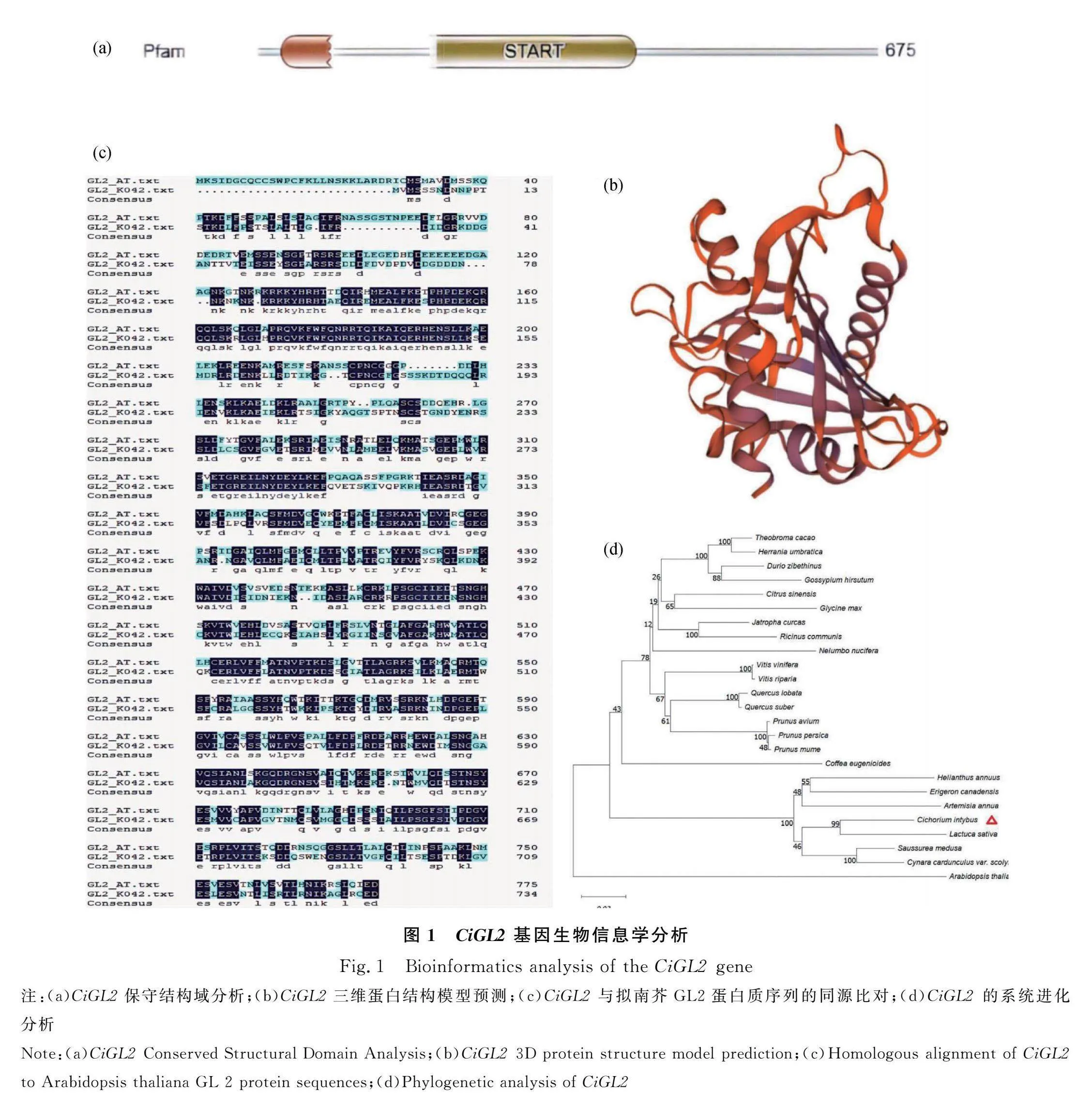
2.1 CiGL2基因生物信息學分析
CiGL2的開放閱讀框為2208 bp,編碼735個氨基酸。CiGL2蛋白保守結構及特性分析預測(圖1a)在26~82氨基酸之間含有同源結構域HOX(homeobox),在191~415氨基酸之間含有START(StAR和磷酰膽堿轉移蛋白)。CiGL2蛋白質三級結構預測(圖1b)顯示在不同的折疊處,包括轉角和不規則的卷曲均可發生與DNA/RNA結合。CiGL2與擬南芥AtGL2氨基酸序列同源對比(圖1c)顯示二者共有“PRQVKFWFQNRRTQTKAIQERHENSLLK”序列,初步預測該氨基酸序列是‘將軍’菊苣CiGL2和擬南芥AtGL2的保守區。CiGL2與其他24種植物的GL2同源蛋白序列構建進化樹(圖1d)結果顯示CiGL2蛋白與生菜(Lactuca sativa varL.t.)GL2蛋白的氨基酸序列同源性比較高,為87.27%;與向日葵(Helianthus annuus L.)氨基酸序列同源性較低,為80.82%。
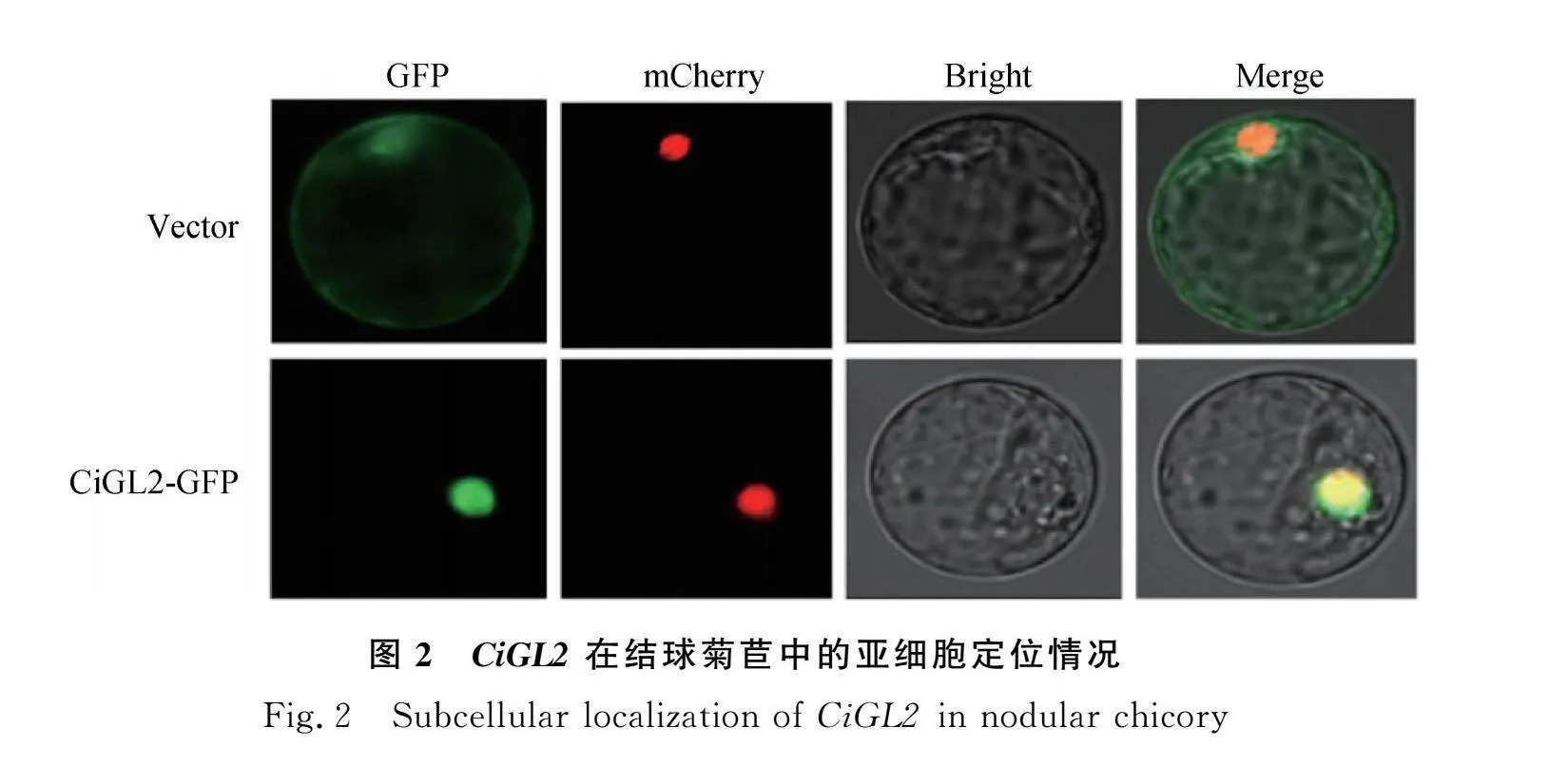
2.2 CiGL2亞細胞定位
共聚焦激光顯微鏡照片結果顯示(圖2):1105:GFP在激發光下的熒光信號分布于整個結球菊苣細胞,1104-CiGL2:GFP在細胞核捕獲到強烈紅色信號蛋白,表明CiGL2蛋白的亞細胞定位在細胞核中。
2.3 CiGL2基因表達分析:
熒光定量分析(圖3)顯示CiGl2基因在根、莖、葉、幼根及幼葉均有表達,根、莖、葉表達量最低;幼芽的表達量最高,說明CiGL2可能在幼芽發育階段有重要作用。
2.4 gl2-3突變體和過表達恢復株系基因表達分析
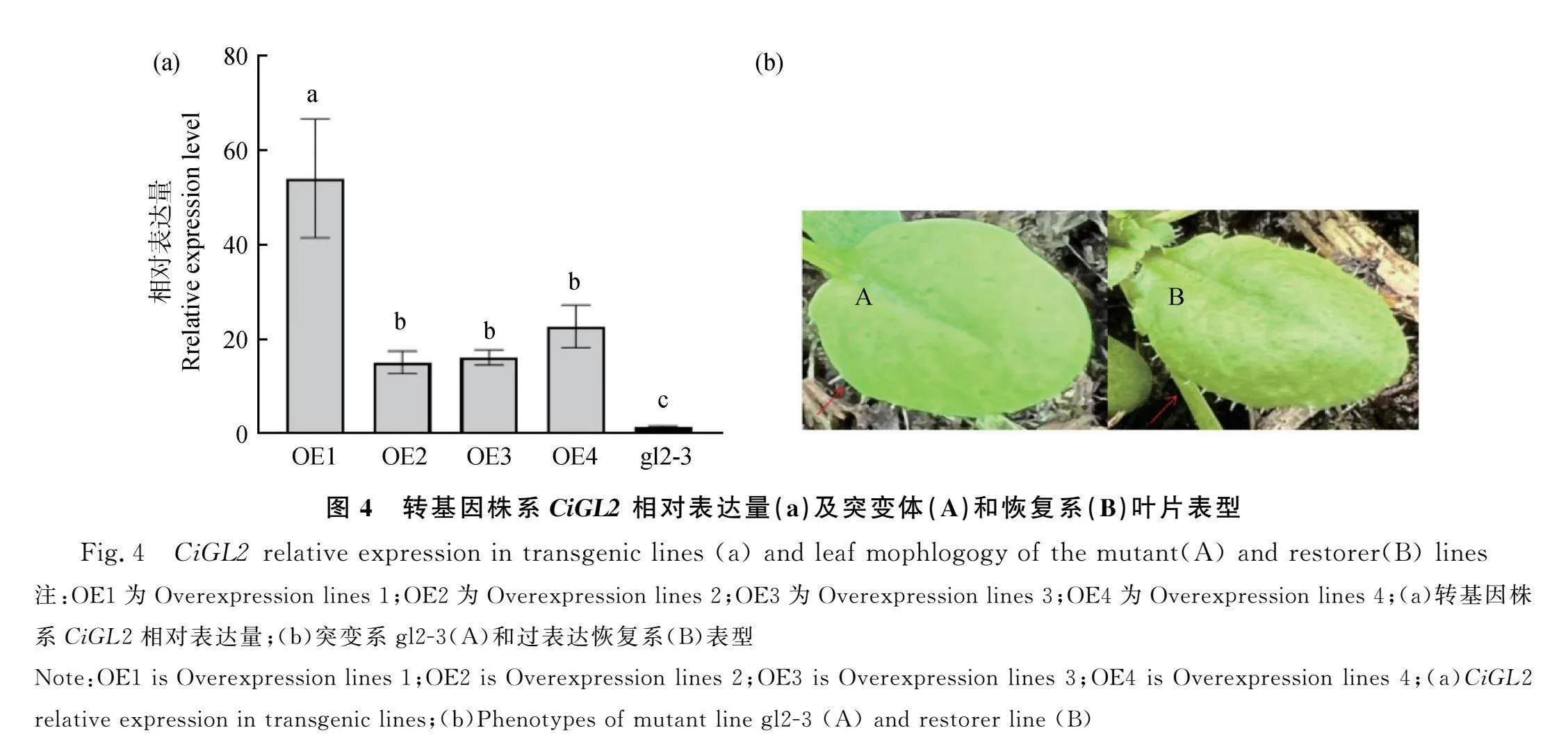
突變體和過表達株系的基因表達結果如圖(圖4a),過表達株系目標基因相對表達量顯著升高,尤其是OE1(Overexpression lines 1)與對照突變植株(gl2-3)相比表達量急劇增加。葉片表型(圖4b)圖片顯示恢復系植株葉片邊緣表皮毛的密度上升;葉片腹面出現表皮毛,表皮的長度較短。
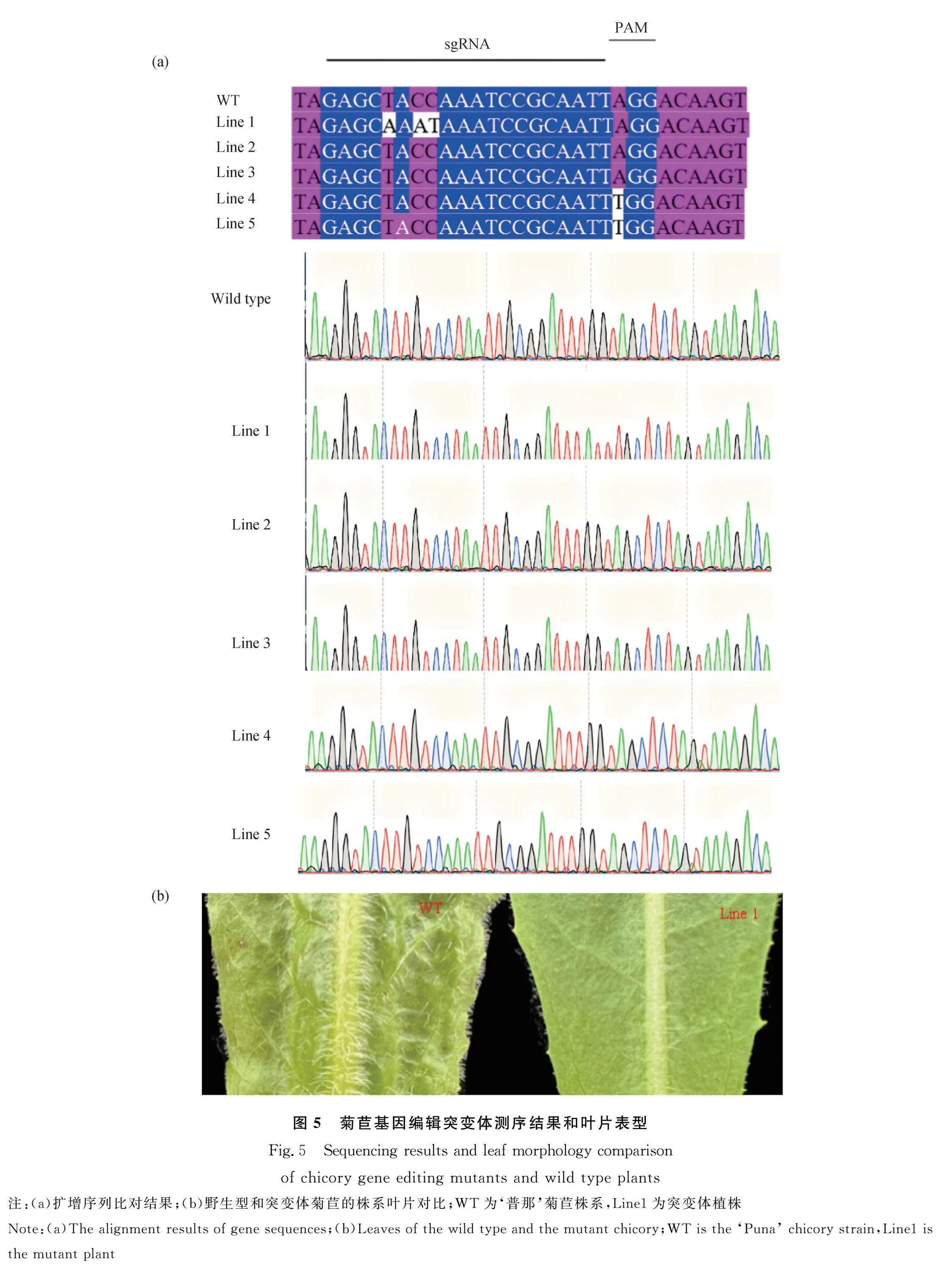
2.5 菊苣基因編輯突變體鑒定和表型分析
CTAB法提取‘普那’菊苣株系葉片基因組DNA,PCR后測序驗證,獲得3個突變株系:Line1、Line4、Line5 (圖5a)。Line 4和Line 5產生堿基A的替換,由于密碼子的簡并性,該單堿基突變未影響氨基酸編碼,葉片表型無差異;Line 1靶位點的第5,7和8位發生堿基替換。Line 2和Line 3的靶位點沒有發生突變。Line 1是最終驗證獲得的突變株系,與‘普那’菊苣株系對比(圖5b),Line 1葉腹面和背面的表皮毛的長度均變短,表皮毛密度下降。菊苣突變體Line1葉片表皮毛表型發生變化,說明菊苣的表皮毛發育在一定程度上受CiGL2基因影響。
3 討論
GL2基因在擬南芥中研究比較深入,但菊苣GL2基因研究很少。本研究進一步證明CiGL2基因在菊苣表皮毛形成中具有重要生物學功能。GL2編碼的蛋白質屬于同源結構域亮氨酸拉鏈(HD-ZIP)IV類轉錄因子家族,被認為參與表皮發育的關鍵調節因子。擬南芥AtGL2的START結構域與脂質配體結合,通過START結構域的脂質結合特性,在表皮細胞分化中起關鍵作用[19]。菊苣CiGL2中也含有START結構。CiGL2蛋白三級結構預測在折疊處能夠與DNA/RNA結合,可能該同源結構域HOX(homeobox)起到重要作用[20]。
菊苣CiGl2基因在幼芽中表達量最高,在幼葉中也有所表達。擬南芥GL2基因研究顯示其主要也在幼葉原基表達[21],這些器官可能需要更高的GL2表達閾值才能產生毛狀體[22]。擬南芥gl2突變體多表現為葉片的表皮毛退化或數量減少[11],而包含完整GL2 轉錄單元的gl2轉基因系能夠觀察到正常的根毛產生[23]。在本研究中,過表達CiGL2基因的恢復株系葉片邊緣表皮毛密度上升,葉片腹面出現表皮毛,說明其一定程度可恢復擬南芥gl2-3突變體葉片的無毛表型,證明該基因是AtGL2的同源基因。
CRISPR/Cas9基因編輯菊苣CiGL2突變植株葉片背面和葉片腹部表皮數量減少,表皮毛密度下降,進一步驗證了CiGL2在調控菊苣葉片表皮毛方面有一定作用。擬南芥AtGL2基因的突變體葉片主要表現為完全無毛或有發育不完全的幾根表皮毛[24]。據此可知,除了CiGL2基因,菊苣葉片表皮毛發育調控可能還有其他基因的參與。
4 結論
菊苣CiGL2亞細胞定位于細胞核,在根、莖、葉、幼根和幼葉均有表達,幼芽的表達量最高。利用擬南芥突變體異源過表達該基因,使葉片恢復了突變體的無毛或少毛表型,恢復系邊緣表皮毛密度上升,葉片腹面也出現表皮毛,說明該基因參與表皮毛發育。菊苣CiGL2突變體葉片背面和葉片腹部表皮數量減少,表皮毛密度下降,進一步驗證了該基因調控菊苣葉片表皮毛發育。本研究為深入研究菊苣表皮毛發育機制提供參考。
參考文獻
[1] 費星宇,趙泓,杜洋,等. 菊苣的基因組大小估算及基因組調查測序[J]. 草地學報,2022,30(12):3207-3214
[2] 金忠民,李春月,劉本松,等. 菌株JB12影響鉛鎘脅迫下菊苣黃酮合成的轉錄組分析[J]. 草地學報,2023,31(6):1648-1655
[3] HUR S N,PARK H S. Developing chicory for forage crop by new technology[J]. Journal of the Korean Society of Grassland Science,1995,15(4):265-273
[4] 李潔,張銘芳,于見麗,等. 植物表皮毛發育調控的研究進展[J]. 北方園藝,2020(23):133-139
[5] 陳文婷,李慧靈,肖強,等. 木芙蓉不同時期花的結構變化研究[J]. 北方園藝,2016(7):50-54
[6] RIDDICK E W,SIMMONS A M,et al. Do plant trichomes cause more harm than good to predatory insects[J]. Psat Management Science,2014,2014,70(11):1655-1665
[7] RIDDICK E W,SIMMONS A M,et al. Plant trichomes have mixed impacts on predatory insects[J]. Psat Management Science,2014,70(11):1668-1668
[8] LI J,TANG B,LI Y,et al. Rice SPL10 positively regulates trichome development through expression of HL6 and auxin-related genes[J]. Plant Biology,2021,63(8):1521-1537
[9] GALINDO-TRIGO S. STARTing to dissect the molecular determinants of GLABRA2 activity[J]. Plant Physiol,2022,190(4):2064-2065
[10]MUKHERJEE T,SUBEDI B,KHOSLA A,et al. The START domain mediates Arabidopsis GLABRA2 dimerization and turnover independently of homeodomain DNA binding[J]. Plant Physiology,2022,190(4):2315-2334
[11]QING L,AOYAMA T. Pathways for epidermal cell differentiation via the homeobox gene GLABRA2:update on the roles of the classic regulator[J]. Journal of Integrative Plant Biology,2013,55(5):485
[12]SZYMANSKI D B,LLOYD A M,MARKS M D. Progress in the molecular genetic analysis of trichome initiation and morphogenesis in Arabidopsis[J]. Trends in Plant Science,2000,5(5):214-219
[13]RERIE W G,FWLDMANN K A,MARKS M D. The GLABRA2 gene encodes a homeo domain protein required for normal trichome development in Arabidopsis[J]. Genes amp; Development,1994,8(12):1388-1399
[14]陳春花.黃瓜表皮毛發育及其相關基因CsTTG1和CsGL2的功能分析[D].北京:中國農業大學,2016:20-21
[15]LIN Q,OHASHI Y,KATO M,TSUGE T,et al. GLABRA2 directly suppresses basic helix-loop-helix transcription factor genes with diverse functions in root hair development[J]. Plant Cell,2015,27(10):2894-2906
[16]FAN W,WANG S,WANG H,et al. The genomes of chicory,endive,great burdock and yacon provide insights into Asteraceae palaeo-polyploidization history and plant inulin production[J]. Molelcular Ecology Resources,2022,22(8):3124-3140
[17]張建福.一種水稻原生質體分離與轉化方法:中國,201911018183.9[P]. 2023-11-02
[18]夏曾潤,王文穎,劉亞琪,等. 羅布麻K~+通道編碼基因AvAKT1的克隆與表達分析[J]. 草業學報,2019,28(8):180-189
[19]NAGATA K,ABE M. A conserved mechanism determines the activity of two pivotal transcription factors that control epidermal cell differentiation in Arabidopsis thaliana[J]. Journal of Plant Research,2023,136(3):349-358
[20]MUKHERJEE T,SUBEDI B,KHOSLA A,et al. The START domain mediates Arabidopsis GLABRA2 dimerization and turnover independently of homeodomain DNA binding[J]. Plant Physiology,2022,190(4):2315-2334
[21]SZYMANSKI D B,JILK R A,POLLOCK S M,et al. Control of GL2 expression in Arabidopsis leaves and trichomes[J]. Development,1998,125(7):1161-1171
[22]WANG S,BAEEON C,SCHIEFELBEIN J,et al. Distinct relationships between GLABRA2 and single-repeat R3MYB transcription factors in the regulation of trichome and root hair patterning in Arabidopsis[J]. New Phytologist,2010,185(2):387-400
[23]MASUCCI J D,RERIE W G,FOREMAN D R,et al. The homeobox gene GLABRA 2 is required for position-dependent cell differentiation in the root epidermis of Arabidopsis thaliana[J]. Development,1996,122(4):1253-1260
[24]OHASHI Y,OKA A,RODRIGUES-POUSADA R,et al. Modulation of phospholipid signaling by GLABRA2 in root-hair pattern formation[J]. Science,2003,300(5624):1427-1430
(責任編輯 劉婷婷)
