西氏熊蜂線粒體基因組結構特征及系統發育分析
2024-12-31 00:00:00馬曉璇梁程博劉道鑫孫國閆京艷
草地學報 2024年8期

摘要:本研究利用二代測序技術對西氏熊蜂(Bombus sichelii)的線粒體基因組進行了測序、組裝和注釋,并對其線粒體基因組的組成、結構特征、基因重排及系統發育等方面進行了生物信息學分析。結果表明:西氏熊蜂線粒體基因組長度為15 711 bp,包含13個蛋白編碼基因、22個tRNA基因和2個rRNA基因;線粒體基因組全序列、蛋白編碼基因的串聯序列、tRNA串聯序列、rRNA串聯序列的A+T含量均大于80%,表現出明顯的堿基偏倚;13 個蛋白編碼基因的起始密碼子均為ATN,終止密碼子為TAA,TAG或T --。tRNA二級結構預測發現,trnS1與其他昆蟲具有相同的特征,即缺少DHU臂和DHU環,其余21個tRNA均能形成典型的三葉草結構;系統發育進化分析結果高度支持西氏熊蜂與火紅熊蜂(B. pyrosoma)隸屬于黑熊蜂亞屬(Melanobombus)。本研究首次獲得西氏熊蜂線粒體基因組序列,初步鑒定了其線粒體基因組的特征,為進一步對熊蜂線粒體基因組學和系統發育關系的研究奠定了基礎。
關鍵詞:西氏熊蜂;線粒體基因組;生物信息學;系統發育分析
中圖分類號:S891;Q963""" 文獻標識碼:A"""" 文章編號:1007-0435(2024)08-2408-11
The Mitochondrial Genome Characteristics and Phylogenetic Analysis
of Bombus sichelii
MA Xiao-xuan, LIANG Cheng-bo, LIU Dao-xin, SUN Guo, YAN Jing-yan*
(School of Agriculture and Animal Husbandry, Qinghai University, Xining, Qinghai Province 810016, China)
Abstract:In this study,the mitochondrial genome of Bombus sichelii was sequenced by next-generation sequencing technology,then was assembled and annotated. And the composition,structural characteristics,gene rearrangement and phylogeny of the mitochondrial genome were analyzed by bioinformatics. The results showed that the mitochondrial genome of B. sichelii was 15 711 bp in length,containing 13 protein-coding genes,22 tRNA genes and 2 rRNA genes. The A+T content of the whole mitochondrial genome sequence,the tandem sequence of protein-coding genes,the tandem sequence of tRNA,and the tandem sequence of rRNA were all greater than 80%,showing obvious base bias. The initiation codon of 13 protein-coding genes was ATN,and the stop codons was TAA,TAG,or T--. The secondary structure prediction of tRNAs showed that trnS1 had the same characteristics as other insects. That is,it lacked DHU arm and DHU ring,and the other 21 tRNAs could form a typical clover structure. Phylogenetic findings support the affiliation of B. sichelii and B. pyrosoma to Melanobombus. In this study,the mitochondrial genome sequence of B. sichelii was obtained for the first time,and the characteristics of the mitochondrial genome of Bumblebee were preliminarily identified,which laid a foundation for further research on the relationship between mitochondrial genomics and phylogeny of bumblebees.
Key words:Bombus sichelii;Mitochondrial genome;Bioinformatics;Phylogenetic analysis
熊蜂(Bombus spp.)是膜翅目(Hymenoptera)、細腰亞目(Apocrita)、蜜蜂科(Apidae)、熊蜂屬(Bombus)昆蟲的總稱[1]。熊蜂是眾多野生植物的重要傳粉者,在高海拔生態系統中,鳥類和蝴蝶的傳粉作用逐漸減弱,熊蜂則由于其對高海拔環境的高度適應性、較強的飛行能力和高的花粉傳遞效率成為主導的傳粉者,對維持高海拔生態系統的平衡和穩定至關重要[2-4]。但由于棲息地喪失、環境污染和氣候變化等多種因素的影響,許多生境內的熊蜂種類及數量顯著降低,目前已經有36種熊蜂被列入國際自然保護聯盟瀕危物種紅色名錄[5-8]。我國在近些年的調查中發現,黑足熊蜂(B. atripes)已經從云南省內消失[9],所以熊蜂物種多樣性的保護已迫在眉睫。
線粒體基因組(Mitochondrial genome,mtDNA)是在線粒體內發現的小型環狀染色體,具有基因進化速率快、母系遺傳、高拷貝數、結構簡單及易獲得等特點,已廣泛應用于物種鑒定、種群遺傳結構、系統發育和系統地理學等研究領域[10-11]。熊蜂的傳統形態學鑒定主要以體色圖案為依據,然而,一些不同的熊蜂物種常常出現相似的毛色配置,而同種內不同級型或不同區域的個體間毛色也會存在較大變異,這些都給熊蜂的形態分類和鑒定帶來了較大的困難[12]。隨著分子生物學技術的快速發展,基于線粒體基因序列等分子標記的熊蜂分類學和遺傳學研究日益受到研究人員的關注。其中,COI,Cytb,16S rRNA和微衛星標記等是熊蜂中普遍使用的分子標記,Bertsch[13]和劉蘋等[14]利用COI基因明確了明亮熊蜂復合種的分類地位及其分布特性;Pirounakis等[15]利用Cytb基因分析發現B. pascuorum在阿爾卑斯山的13個不同地理分區中存在遺傳變異。但由于單一基因所攜帶的遺傳信息較少,所以多基因聯合分析在熊蜂研究中日益增加。例如Williams等[16]利用16S rRNA和COI 2種分子標記,對山熊蜂亞屬(Alpinobombus)中基于形態學鑒定存在問題的物種進行了綜合鑒定;Cwjas等利用線粒體基因標記和微衛星標記,研究了伊比利亞半島上地熊蜂(B. terrestris)的時空格局[17]。隨著高通量測序技術的發展,線粒體基因組序列的獲得變得越來越容易,2023年通過對山東和遼寧2個地區的日光蜂(Aphelinus mali)[18]進行線粒體基因組測序分析,發現2個支系之間僅存在歷史地理隔離,尚未出現種群間的生殖隔離,證明了2個支系仍然屬于同一物種。所以在分子技術不斷發展的情況下,利用分子手段比利用形態學鑒定熊蜂更加高效準確。近年來,通過眾多學者不斷豐富線粒體基因的研究,線粒體基因在物種鑒定、遺傳進化等方面的應用價值正逐漸增強。
西氏熊蜂隸屬于黑熊蜂亞屬,是目前眾多學者普遍認同的分類結果,且在我國大部分省份都有分布[2]。2023年胥勛露等[7]在對青藏高原東北部邊緣的調查研究中,發現西氏熊蜂是此地區的常見種,廣泛分布在 3100~3600 m的高海拔地區,并且西氏熊蜂的訪花植物種類涉及范圍達到5科11種,其中對菊科(Asteraceae)植物、豆科(Fabaceae)植物等具有明顯的訪問偏好。近年來,對熊蜂的研究工作主要是集中在形態學鑒定、蜜源植物的調查、種質資源的保護及人工繁育等方面,但對線粒體基因組方面的分子研究尚不完備,其中黑熊蜂亞屬僅有火紅熊蜂(B. pyrosoma)和B. lapidarius的線粒體基因組數據被記錄。鑒于此,本研究首先補充西氏熊蜂線粒體基因組數據,即采用二代測序技術對西氏熊蜂的線粒體基因組測序、組裝、注釋,然后對其線粒體基因組的組成和結構特征等方面進行分析,豐富熊蜂線粒體基因組的基礎研究;最后采用最大似然法(Maximum likelihood,ML)和鄰接法(Neighbor-Joining,NJ)構建基于13個亞屬熊蜂線粒體基因組全序列的進化樹,探討西氏熊蜂在熊蜂屬內的分類地位及各熊蜂亞屬間的進化關系,從而為后續的群體遺傳學研究提供重要的基礎信息。
1 材料與方法
1.1 標本采集
西氏熊蜂樣品2021年7月在青海省玉樹市(33°11′ N,97°24′ E)采集,浸泡于無水乙醇中并置于-80℃冰箱內,保存在青海大學農牧學院昆蟲學實驗室。
1.2 線粒體基因組的測序、組裝及注釋
西氏熊蜂樣品送至北京諾禾致源生物科技有限公司構建350 bp的小片段測序文庫并在Illumina NovaSeq平臺進行高通量測序,測序讀長為PE150。利用Getorganelle軟件對公司測得的clean reads進行組裝,獲得西氏熊蜂的線粒體基因組序列[19]。采用MITOS在線服務器(http://mitos.Bioinf.uni-leipzig.de)對西氏熊蜂線粒體基因組序列進行功能注釋。將注釋結果導入Geneous 9.0.2對比參考序列,手動調整蛋白編碼基因起始密碼子和終止密碼子位置,遵循盡可能少的基因間隔和基因重疊且起始終止密碼完整的原則。
1.3 西氏熊蜂的生物信息學分析
采用在線網站CGView Server(http://cgview.ca/)繪制西氏熊蜂線粒體基因組圈圖。利用Phylosuite v1.2.2提取并分別計算西氏熊蜂線粒體基因組中各編碼基因的堿基組成、AT偏倚(AT-skew)和GC偏倚(GC-skew),其中,堿基偏倚的計算公式為:AT偏倚= (A-T) / (A+T)、GC偏倚= (G-C) / (G+C)。利用Phylosuite v1.2.2和Rstudio 4.3.0分析并繪制蛋白編碼基因的相對同義密碼子使用度柱狀圖(Relative synonymous codon usage,RSCU)[20]。采用在線軟件MITOS和ARWEN(http://130.235.244.92/ARWEN/)對tRNA基因進行查找及二級結構的預測[21]。對西氏熊蜂和其余11種熊蜂及果蠅(Drosophila yakuba)的線粒體基因組序列,利用在線網站ITOL(https://itol.embl.de)繪制13條序列中37種基因的排列,分析不同基因的重排。
1.4 西氏熊蜂系統發育分析
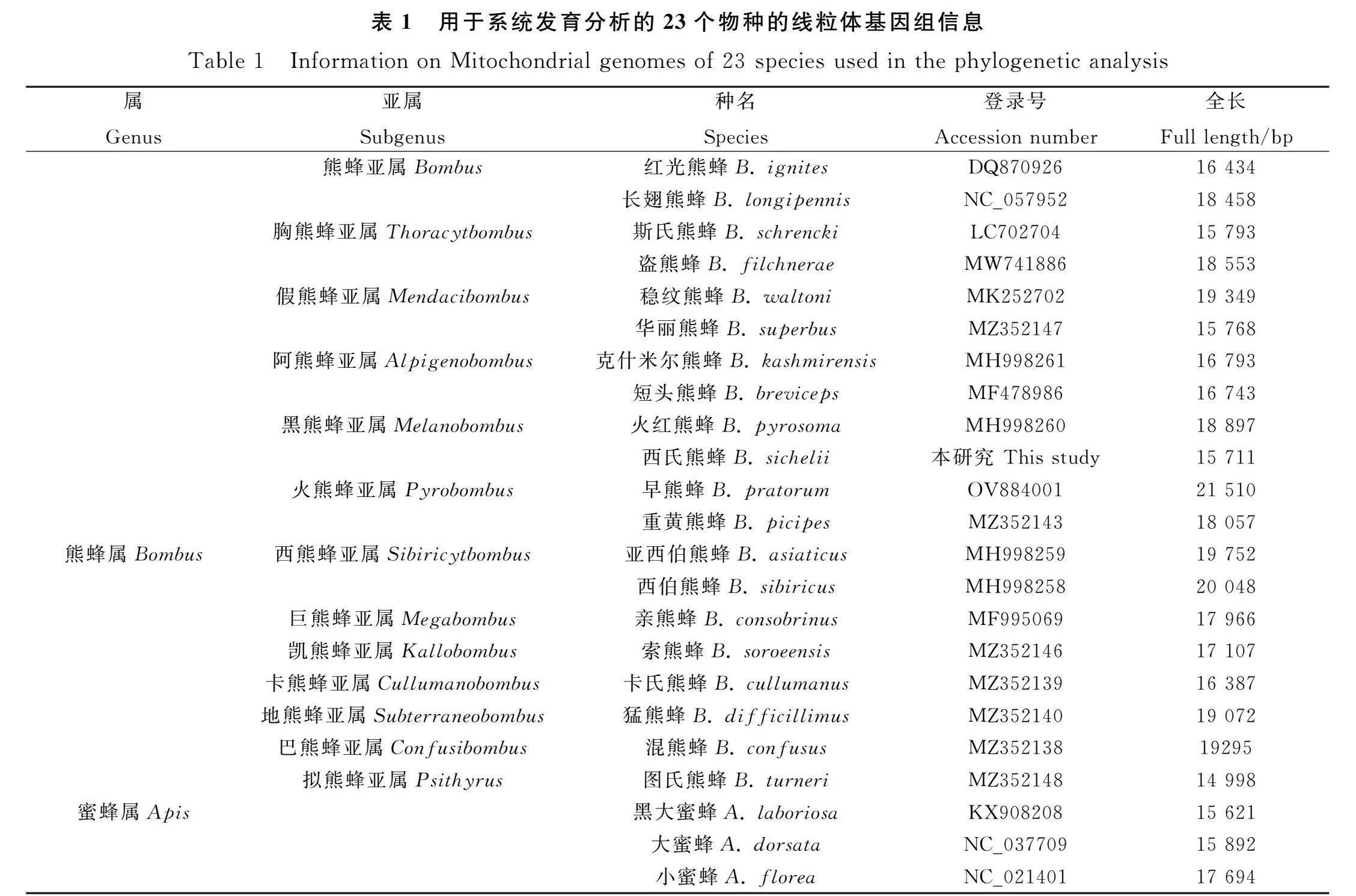
為研究西氏熊蜂與其他熊蜂的系統發育關系,選取已報道的熊蜂屬13個亞屬的20條序列作為內群,蜜蜂屬的3種昆蟲為外群(表1),使用MEGA 10軟件中的鄰接法和最大似然法構建系統進化樹[22]。
2 結果與分析
2.1 西氏熊蜂線粒體基因組的結構特征分析
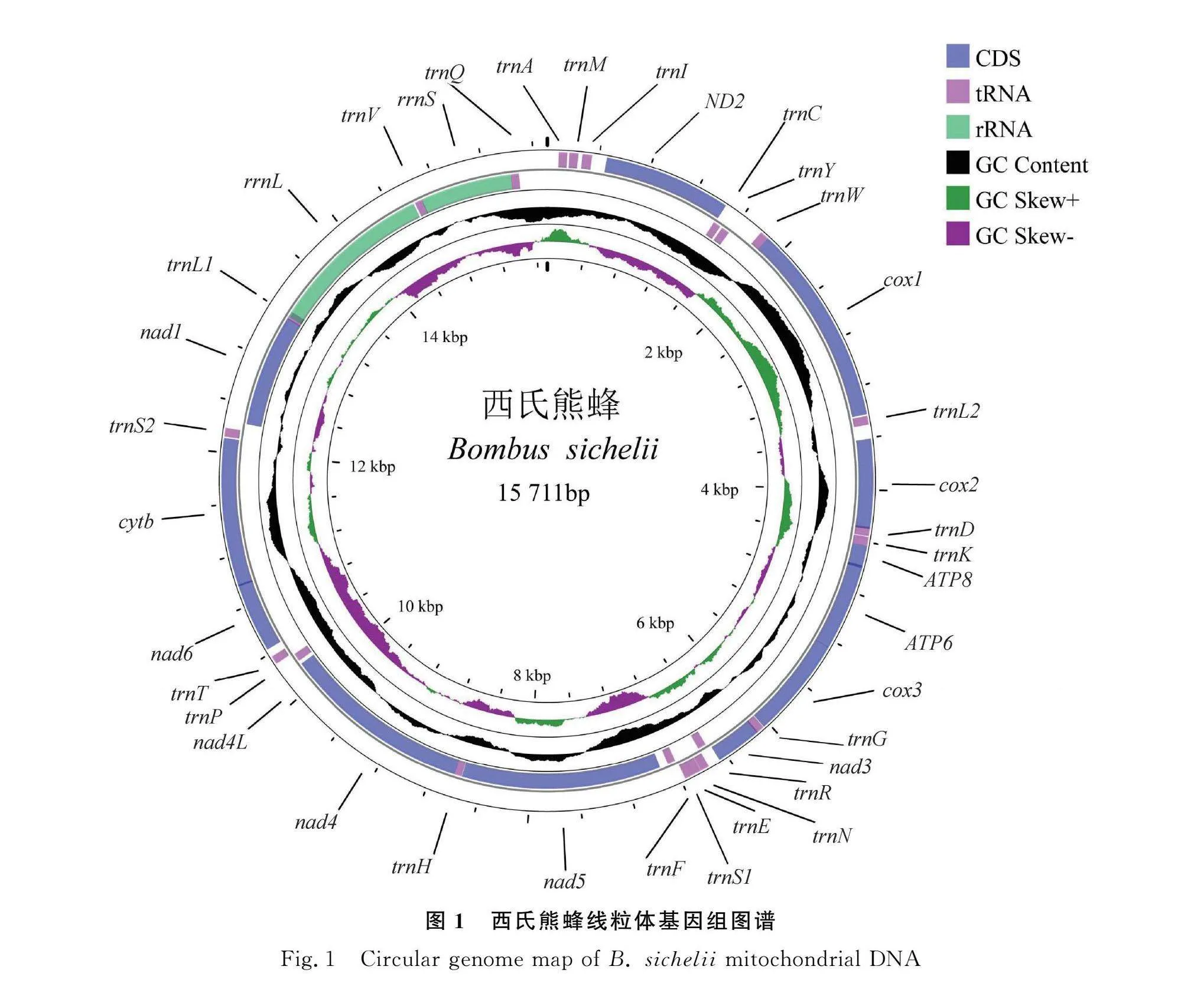
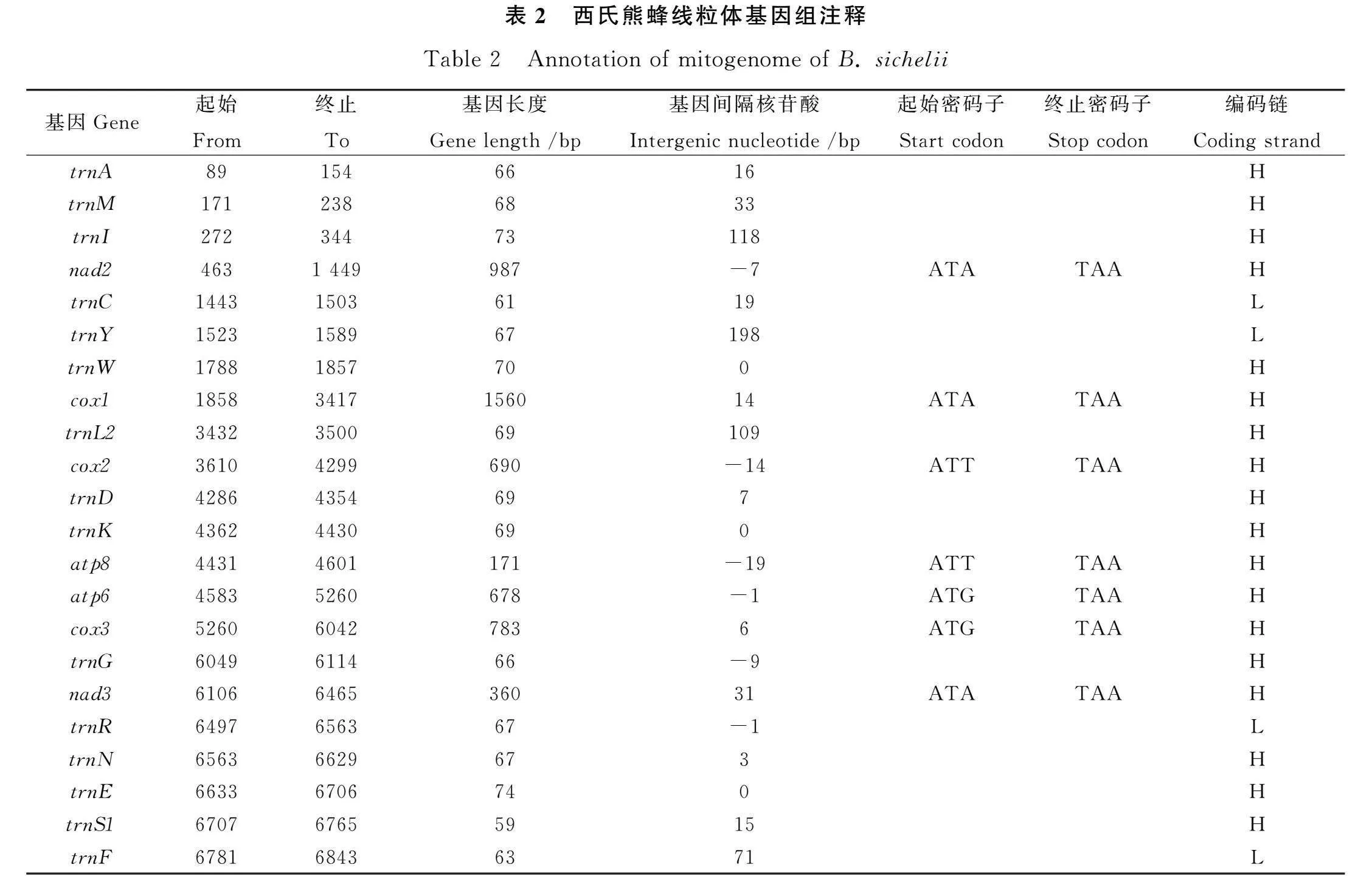
西氏熊蜂線粒體基因組長度為15 711 bp,包含37個基因,分別為13個蛋白編碼基因(protein-coding genes,PCGs)、22個tRNA基因和2個rRNA基因(圖1)。其中有9個PCGs和13個tRNA基因在主要編碼鏈(H鏈)上;4個PCGs、9個tRNA基因和2個rRNA基因在次要編碼鏈(L鏈)上。
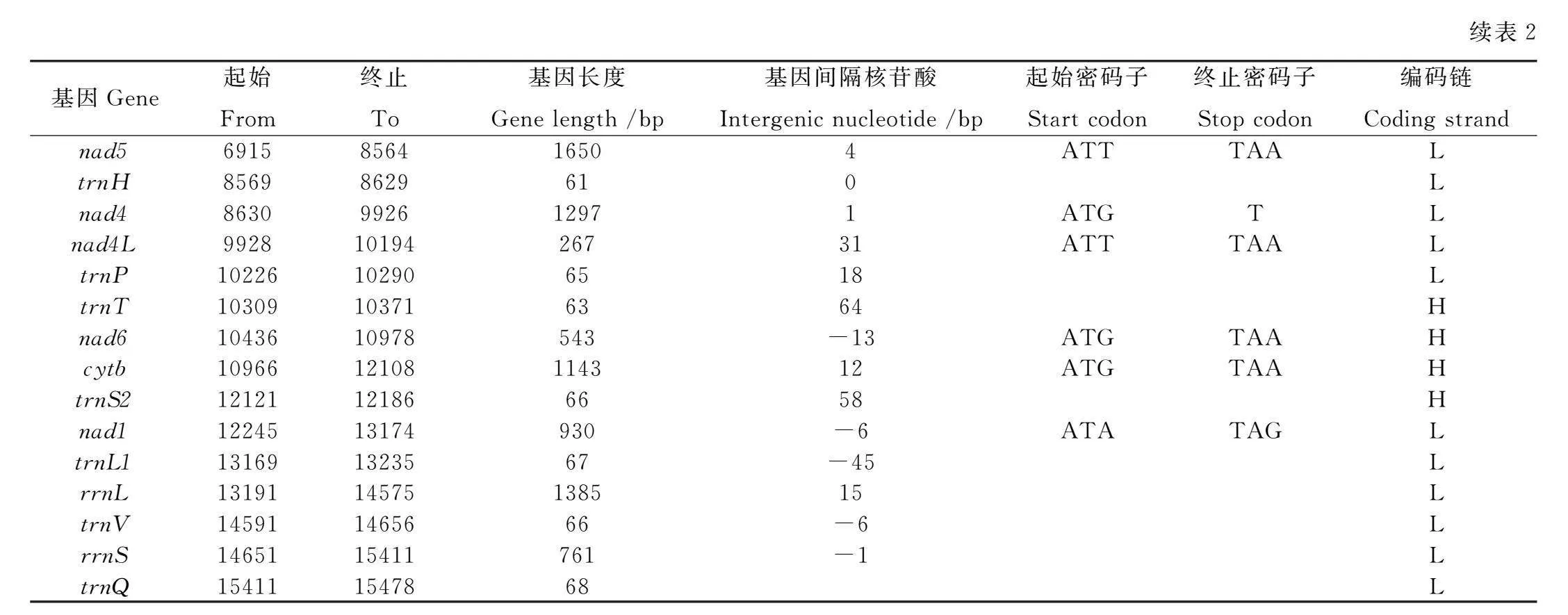
西氏熊蜂的線粒體相鄰基因間存在長度不等的基因間隔和基因重疊,本研究共發現21個基因間隔和11個基因重疊,trnY和trnW間基因間隔最長(198 bp),nad4和nad4L間間隔最短(1 bp);在相鄰基因的基因重疊中,trnL1和rrnL間基因重疊最長,為45 bp(表2)。
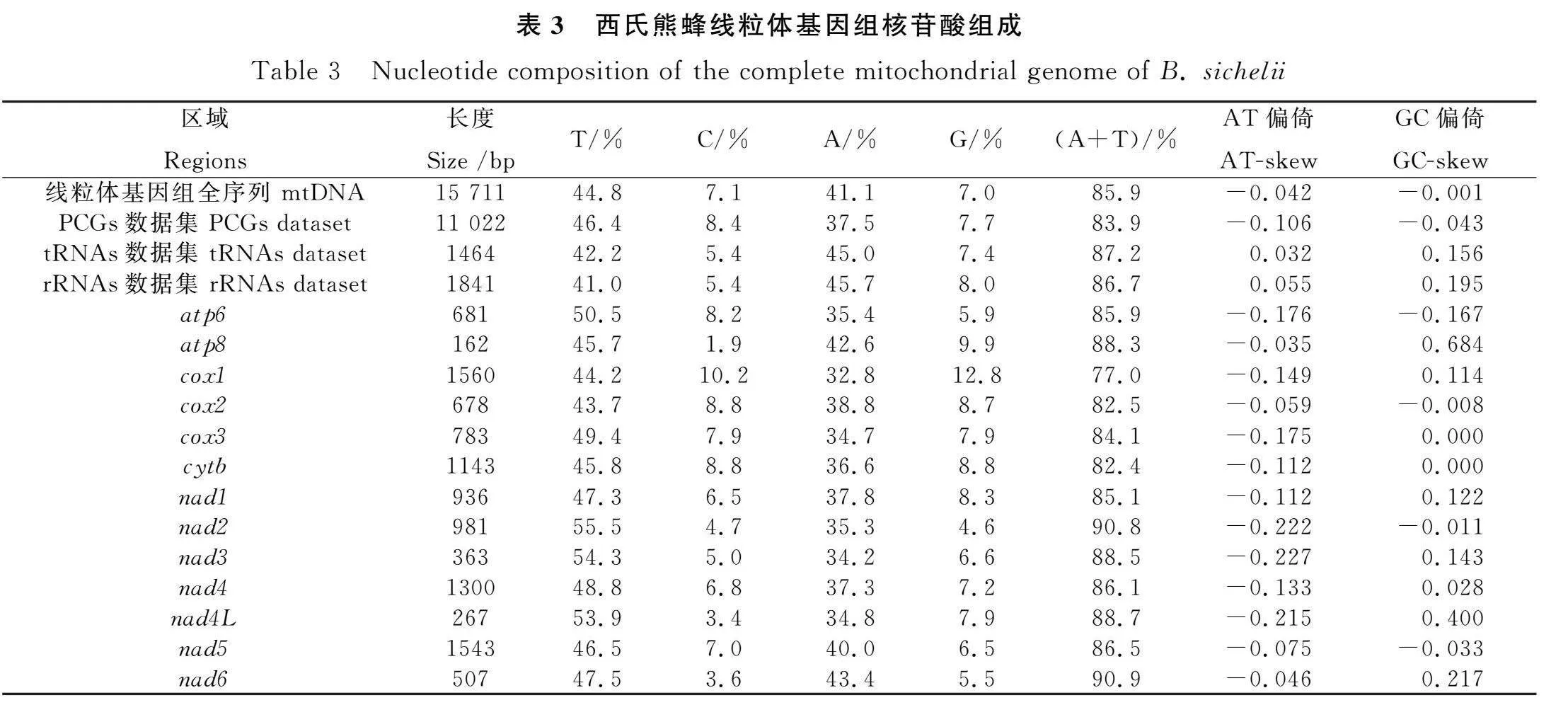
2.2 西氏熊蜂線粒體基因組的堿基組成分析
西氏熊蜂線粒體基因組全序列、蛋白編碼基因、tRNA基因和rRNA基因4個數據集的A+T含量分別為85.9%,83.9%,87.2%和86.7%,其中13個蛋白編碼基因的A+T含量分別在77%~90.9%之間,其中cox1最低,為77%,nad6最高,達到90.9%(表3)。對每個蛋白編碼基因的AT偏倚分析中發現,所有蛋白編碼基因都呈現T偏倚,但在GC偏倚分析中4個蛋白編碼基因(atp6,cox2,nad2,nad5)呈現C偏倚,cox3和cytb無堿基偏倚,其余7個蛋白編碼基因呈現G偏倚。對4個數據集的堿基偏倚分析發現,13個PCGs串聯數據集與線粒體基因組全序列的堿基偏倚結果一致,均對T堿基和C堿基具有明顯的堿基偏倚;22個tRNAs和2個rRNAs數據集的分析結果卻表明對A堿基、G堿基具有明顯的堿基偏倚。所以與其他數據集相比,13個PCGs的串聯數據集與線粒體基因組的組成特征更加一致。
2.3 西氏熊蜂蛋白編碼基因的密碼子使用分析
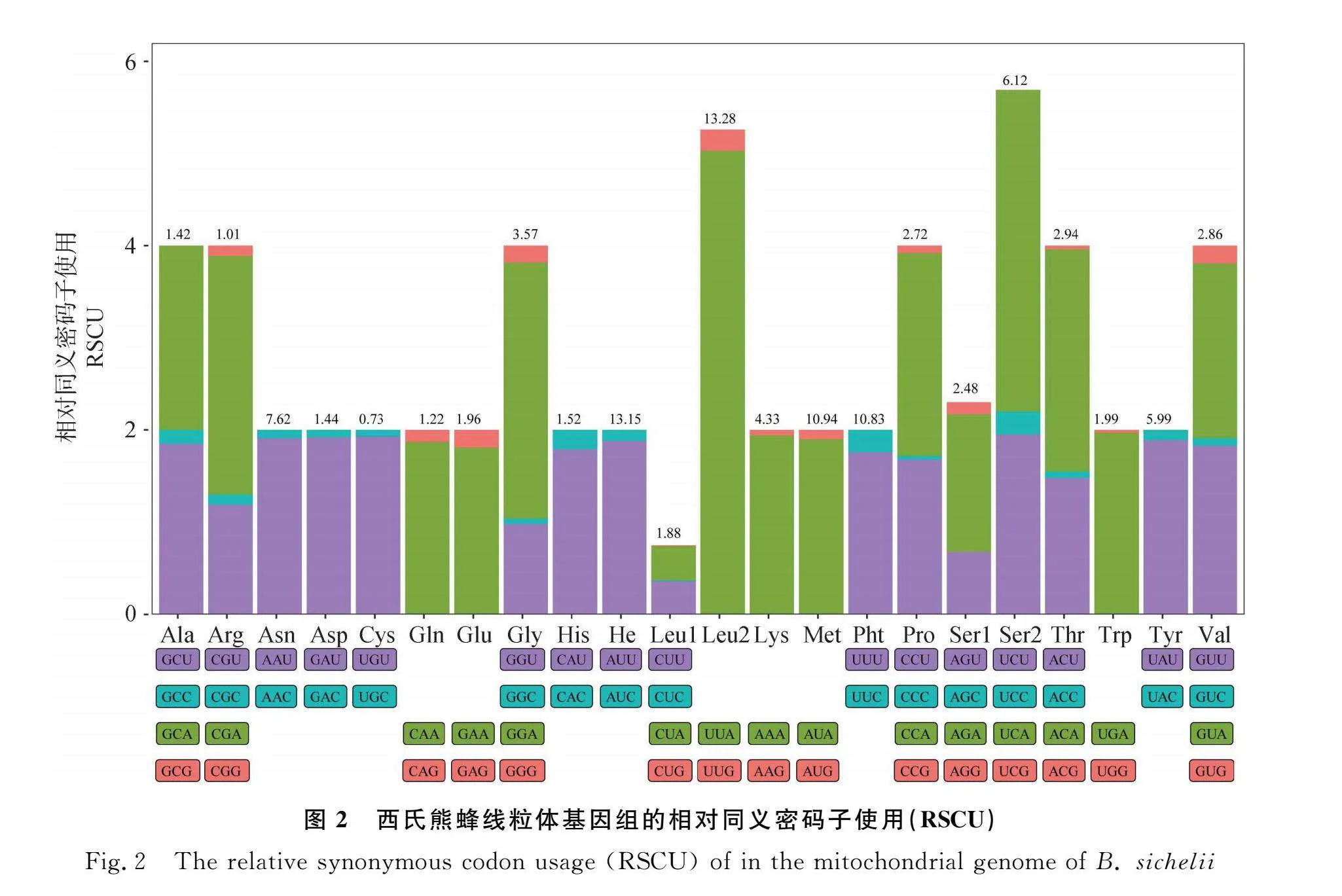
西氏熊蜂13個蛋白編碼基因的串聯序列總長度為11 022 bp,占全基因組的71.15%,共編碼了3 674個氨基酸。氨基酸的百分含量由高到低排列如下:Leu>Ile>Met>Phe>Ser>Asn>Tyr>Lys>Gly>Thr>Val>Pro>Trp>Glu>His>Asp>Ala>Gln>Arg>Cys。其中氨基酸占比最高的是亮氨酸(Leu),為15.16%,半胱氨酸(Cys)占比最少,為0.73%。在相對同義密碼子使用度分析中發現,所有氨基酸對第3位堿基是A/U的密碼子具有明顯使用偏好,如精氨酸(Arg)偏好使用CGA、脯氨酸(Pro)偏好使用CCA、天冬氨酰(Asn)偏好使用AAU、纈氨酸(Val)偏好使用GUU等(圖2)。其中13個蛋白編碼基因的起始密碼子均為ATN,除nad4以不完整的終止密碼子T結尾,其余12個蛋白編碼基因均以完整的終止密碼子(TAA/TAG)結尾。
2.4 西氏熊蜂tRNA二級結構預測分析
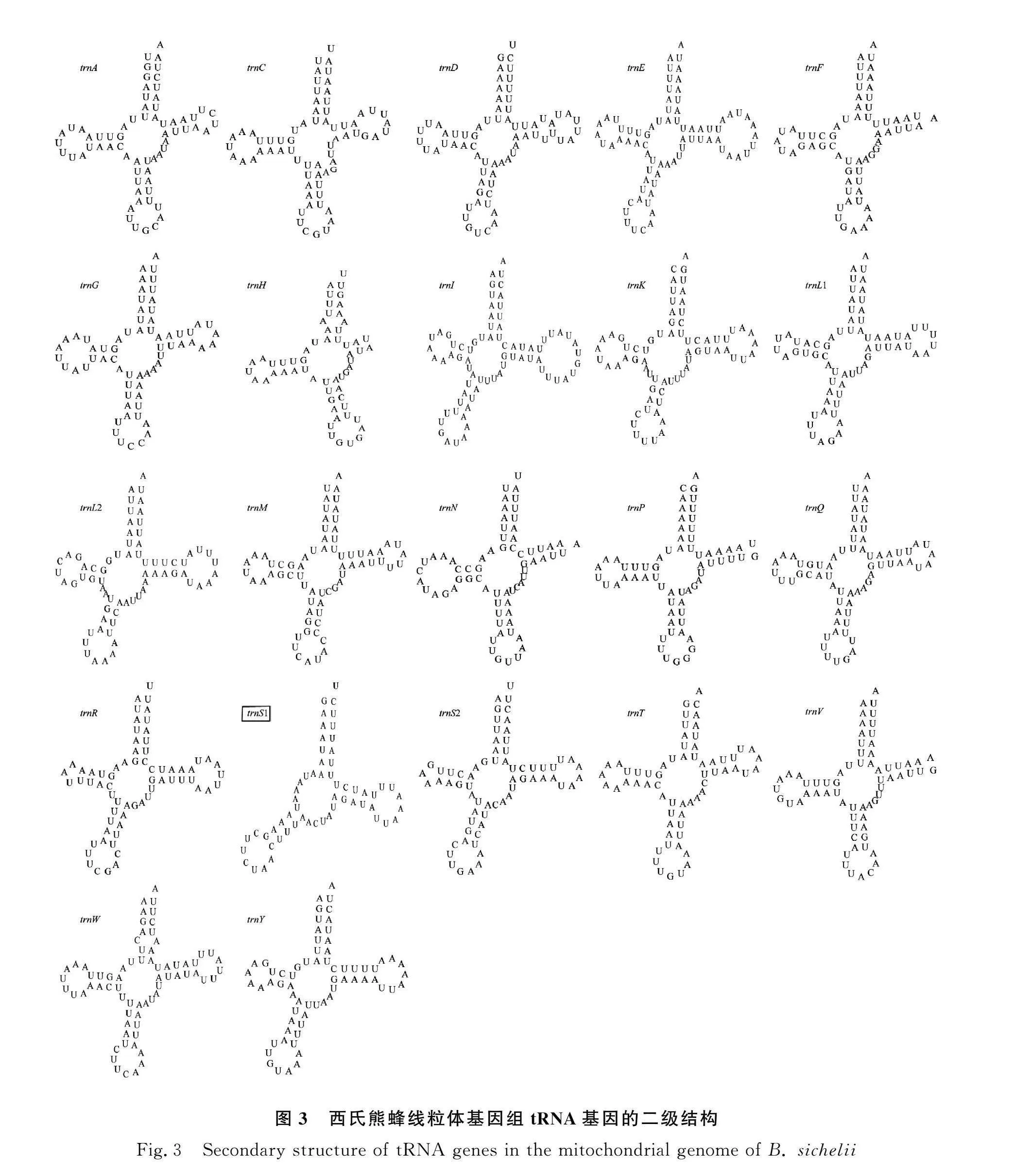
西氏熊蜂的線粒體基因組注釋得到22個tRNA基因,其中13個位于H鏈,9個位于L鏈,單個tRNA基因的序列長度在59~74 bp之間。tRNA基因的二級結構預測結果如圖3所示,其中trnS1因缺少1個二氫尿嘧啶臂(DHU臂)和二氫尿嘧啶環(DHU環)不能形成典型的三葉草結構,其余tRNA均能構成典型的三葉草結構,但trnF,trnH,trnN和trnV基因的二級結構中缺少TφC環。22個tRNA中共發現12次G-U堿基錯配,分別在trnA,trnC,trnD,trnH,trnI,trnK,trnP,trnL1,trnQ和trnV中各出現1次,trnF中出現2次。
2.5 線粒體基因組的基因重排分析
熊蜂與果蠅的線粒體基因排列相比具有明顯不同(圖4),其基因重排主要發生在以下6個區域:trnK移位的trnD-trnK基因簇;trnA移位的trnQ-trnA-trnM-trnI基因簇;trnS1移位的trnE-trnS1-trnF基因簇;trnT移位的trnP-trnT基因簇;trnI移位的trnA-trnM-trnI基因簇;trnW移位的trnC-trnY-trnW基因簇。
不同熊蜂亞屬與果蠅線粒體基因重排的分析結果如下:
(1)在trnR的移位分析發現,胸熊蜂亞屬的trnR移位到trnN的下游,形成trnN-trnR基因簇;巨熊蜂亞屬的trnR移位到trnQ的下游,形成trnQ-trnR基因簇,其他5個亞屬則為trnR-trnN基因簇。(2)trnT移位到nad6上游,形成trnP-trnT基因簇,這種基因重排在熊蜂亞屬、黑熊蜂亞屬、阿熊蜂亞屬、西熊蜂亞屬共4個亞屬中均有出現,其他亞屬未見此種基因重排。(3)在trnA的移位分析中共發現2種不同的移位現象,一種是在胸熊蜂亞屬、假熊蜂亞屬、黑熊蜂亞屬、西熊蜂亞屬的西伯熊蜂和阿熊蜂亞屬的克什米爾熊蜂中發現的trnA移位到trnQ下游,形成trnQ-trnA-trnM-trnI基因簇。另一種在熊蜂亞屬、阿熊蜂亞屬的短頭熊蜂和西熊蜂亞屬的亞西伯熊蜂中發現的trnA移位到trnM下游,形成trnQ-trnM-trnA-trnI基因簇。(4)在trnI的移位分析中發現,除假熊蜂亞屬外其他6個亞屬的trnI均移位到nad2上游,形成trnA-trnM-trnI基因簇,而假熊蜂亞屬的trnI卻移位到trnM上游,形成trnA-trnI-trnM基因簇。
本研究除阿熊蜂亞屬和西熊蜂亞屬內具有trnA的基因重排,阿熊蜂亞屬內的短頭熊蜂具有nad2的缺失,其他各亞屬內均無線粒體基因重排。西氏熊蜂與同樣隸屬于黑熊蜂亞屬的火紅熊蜂的線粒體基因排列順序一致。通過上述的比較分析,發現熊蜂各亞屬間具有較為頻繁的基因重排,各亞屬內的基因重排不頻繁。
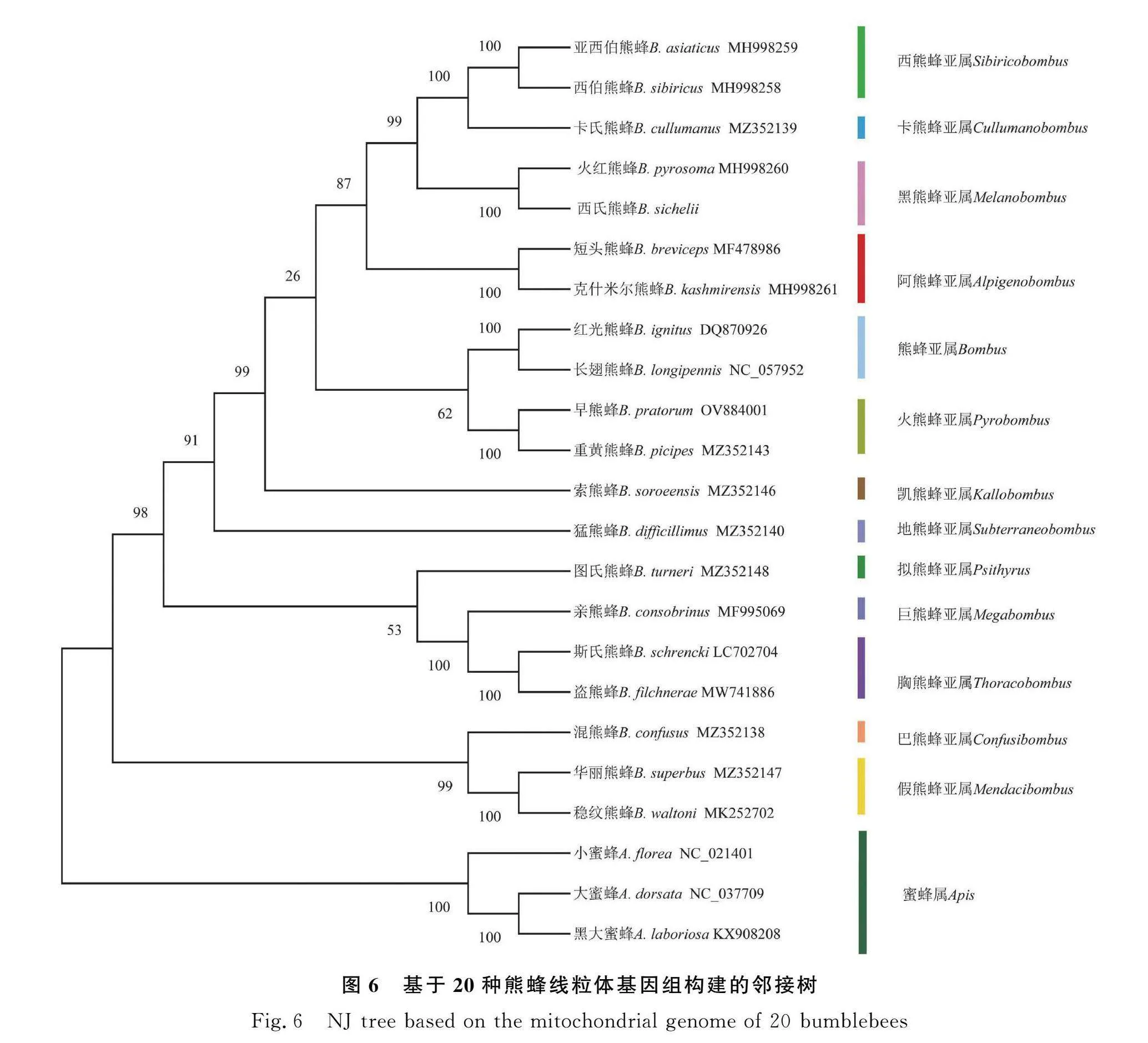
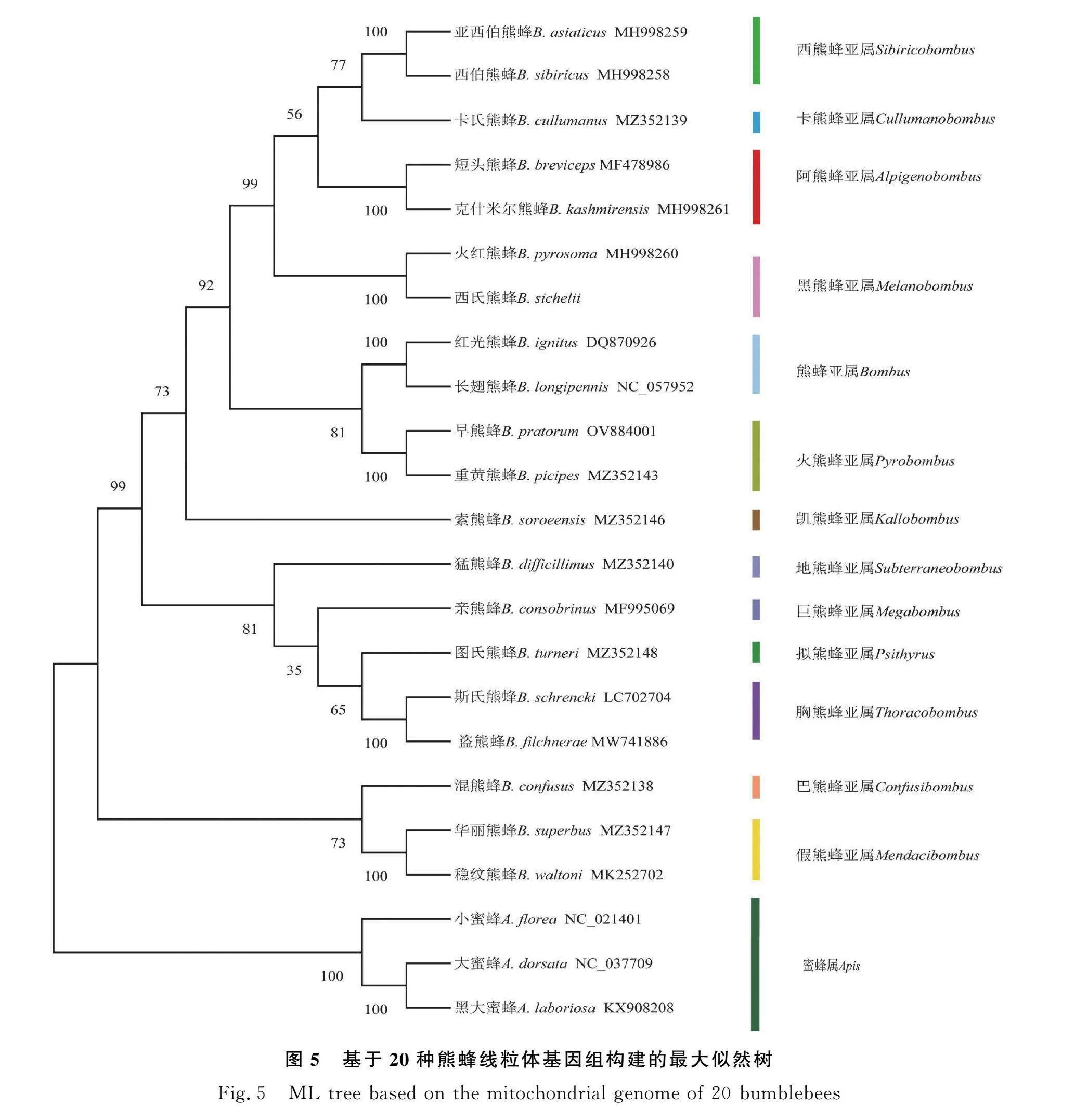
2.6 線粒體基因組系統發育分析
本研究利用最大似然法和鄰接法構建基于20種熊蜂線粒體基因組全序列的系統發育樹(圖5、圖6)。2種方法構建的系統發育樹均高度支持西氏熊蜂與火紅熊蜂共同隸屬于黑熊蜂亞屬,且支持率為100%。2個系統發育樹的拓撲結構對各亞屬之間親緣關系的劃分結果不完全相同,但兩者均證明假熊蜂亞屬(Mendacibombus)和巴熊蜂亞屬(Confusibombus)是最先分化的一支。在2個系統發育樹上可以看出,遠離根部的分支支持率高,對亞屬的分類具有100%的支持率;距離根部近的分支節點支持率低,普遍在80%以下。其中地熊蜂亞屬在ML發育樹中與巨熊蜂亞屬(Megabombus)的親緣關系更近,在NJ發育樹中與凱熊蜂亞屬(Kallobombus)的親緣關系更近,由此可見利用線粒體基因組序列對亞屬間的親緣關系劃分尚且不夠準確。但2個系統發育樹的結果均可以看出短臉型的熊蜂亞屬(Bombus)、黑熊蜂亞屬(Melanobombus)、阿熊蜂亞屬(Alpigenobombus)、火熊蜂亞屬(Pyrobombus)、西熊蜂亞屬(Sibiricytbombus)和卡熊蜂亞屬(Cullumanobombus)之間的親緣關系更近,與長臉型的巨熊蜂亞屬(Megabombus)、胸熊蜂亞屬(Thoracytbombus)和擬熊蜂亞屬(Psithyrus)等亞屬親緣關系相對較遠,這與前人根據頭部的形態學特征[23]和單基因分子鑒定的結果一致。
3 討論
西氏熊蜂首次完成線粒體基因組測序及功能注釋,其長度為15 711 bp。序列組成分析結果表明,西氏熊蜂線粒體基因組存在明顯的堿基偏倚,一方面,與前人所發現的昆蟲線粒體基因組具有高A+T含量的特點相一致,西氏熊蜂的線粒體基因組全序列、蛋白編碼基因、tRNAs基因、rRNAs基因的A+T含量均大于80%;另一方面,在相對同義密碼子的分析中也發現第3位堿基是A/U的密碼子具有明顯的高使用現象[10]。西氏熊蜂線粒體蛋白編碼基因的起始密碼子為典型的ATN,終止密碼子主要是TAA和TAG,僅nad4是以不完整的終止密碼子T結尾,這種使用不完整的終止密碼子在其他膜翅目昆蟲中也時有發生[24]。西氏熊蜂線粒體基因組的蛋白編碼序列全長11 022 bp,共編碼氨基酸3647個,其中頻率最高的依次為亮氨酸(Leu,15.16%)、異亮氨酸(Ile,13.15%)、蛋氨酸(Met,10.94%)和苯丙氨酸(Phe,10.83%),而頻率最低的依次為丙氨酸(Ala,1.42%)、谷氨酰胺(Gln,1.22%)、精氨酸(Arg,1.01%)和半胱氨酸(Cys,0.73%),這種豐度排序與其他膜翅目昆蟲中基本一致[25]。
對西氏熊蜂線粒體基因組的tRNA二級結構預測發現,除了trnS1缺少DHU臂和DHU環,其他都呈現典型的三葉草結構,其中trnS1缺臂的現象在后生動物線粒體基因組中普遍存在[26]。本研究在22個tRNA中共發現12次堿基錯配,這種錯配在tRNA的折疊過程中是被允許的,且在其他后生動物的線粒體中也普遍存在[27]。本研究所發現的堿基錯配為G-U錯配,有學者認為這種錯配可在RNA編輯中校正,并不影響tRNA基因的轉運功能[28]。
果蠅線粒體基因組的基因排列順序一直被認為是昆蟲線粒體基因組的原始排列方式[29]。早在上個世紀,研究人員就已經發現,膜翅目昆蟲的線粒體基因組比其他目的昆蟲存在更為頻繁的基因重排,且細腰亞目的基因重排比廣腰亞目更多[30]。通過將西氏熊蜂線粒體基因組的基因排列與果蠅比較,結果發現明顯的基因重排,且均為tRNA的移位,蛋白編碼基因無基因重排。這與前人發現膜翅目線粒體基因組的基因重排主要是tRNA的重排結果一致[31]。而trnR在巨熊蜂亞屬內的遠距離移位尚不能確定這是否在亞屬內普遍發生,在其他亞屬的研究中也并沒有此類發現。
本研究中,NJ和ML系統發育樹的結果均顯示,西氏熊蜂與火紅熊蜂聚為一支,且與形態學鑒定結果一致均隸屬于黑熊蜂亞屬。由于在2個系統發育樹中,各亞屬的親緣關系不完全一致,但兩棵樹對遠離根部的分支聚類均具有高的支持率(100%),即利用線粒體基因組序列對當前物種亞屬的劃分是具有高支持率的,符合Willims等[32]基于形態學和線粒體基因片段對各亞屬的分類及修訂結果。在2個系統發育樹中,距離根部近的分支節點普遍在80%以下,其結果對探討亞屬間親緣關系的準確度不高,這或許是由于熊蜂物種的線粒體基因組序列不充足導致,所以對熊蜂屬內更多物種的線粒體基因組進行測序,補充其線粒體基因組數據,對后續探索各亞屬間的親緣關系具有重要意義。
4 結論
本研究獲得了西氏熊蜂的線粒體基因組序列,明確了其線粒體基因組組成和結構特征,即線粒體基因組包含37個基因的基本組成,對A/T明顯的堿基偏倚等結果。西氏熊蜂線粒體基因組與已報道的膜翅目中大部分昆蟲的線粒體基因組相似,且與蜜蜂科昆蟲線粒體基因組的一般特征一致。西氏熊蜂的研究結果進一步豐富了熊蜂屬的線粒體基因組數據,對西氏熊蜂的分子鑒定和種群遺傳學研究以及熊蜂屬系統發育關系的深入探討等具有重要意義。
參考文獻
[1] 安建東,黃家興,WILLIAMS P H,等. 河北地區熊蜂物種多樣性與蜂群繁育特性[J]. 應用生態學報,2010,21(6):1542-1550
[2] 黃家興,安建東. 中國熊蜂多樣性、人工利用與保護策略[J]. 生物多樣性,2018,26(5):486-497
[3] 蒙艷華,徐環李,陳軒,等. 塔落巖黃芪主要傳粉蜂的傳粉效率研究[J]. 生物多樣性,2007,15(6):633-638
[4] 何亞平,劉建全. 青藏高原高山植物麻花艽的傳粉生態學研究[J]. 生態學報,2004,24(2):215-220
[5] 梁程博,王久利,孫國,等. 三江源地區熊蜂物種多樣性研究[J]. 草地學報,2022,30(8):2126-2134
[6] 張廣碩,丁桂玲,安建東. 熊蜂主要病蟲害的發生和危害現狀[J]. 應用昆蟲學報,2021,58(2):277-288
[7] 胥勛露,常雪梅,曾桂,等. 甘南草原和若爾蓋濕地的熊蜂物種資源調查[J]. 西華師范大學學報(自然科學版),2024,45(3):239-245
[8] 劉青青,赫苗花,李飛,等. 不同坡向下高寒礦區混播草地的恢復效果評估[J]. 草地學報,2023,31(9):2834-2842
[9] 梁鋮,張學文,黃家興,等. 云南熊蜂地理區劃及物種多樣性分析[J]. 應用昆蟲學報,2018,55(6):1045-1053
[10]朱國淵,張永科,孔祥東,等. 二斑素瓢蟲線粒體基因組全序列測定和分析[J]. 南方農業學報,2023,54(5):1293-1302
[11]ZHANG J Y,ZHANG L P,YU D N,et al. Complete mitochondrial genomes of Nanorana taihangnica and N.yunnanensis (Anura:Dicroglossidae) with novel gene arrangements and phylogenetic relationship of Dicroglossidae[J]. BMC Evolutionary Biology,2018,18(1):26-39
[12]黃家興. 華北地區熊蜂屬(Hymenoptera:Apidae)系統發育的初步研究[D]. 北京:中國農業科學院,2006:2
[13]BERTSCH A. Barcoding cryptic bumblebee taxa:B. lucorum,B. crytarum and B. magnus,a case study[J]. Beitrage Zur Entomol,2009,59(2):287-310
[14]劉蘋,黃家興,吳杰,等. 中國明亮熊蜂復合種的分子鑒定及分布特性[J]. 昆蟲學報,2014,57(2):235-243
[15]PIROUNAKIS K,KOULIANOS S,SCHMID-HEMPEL P. Genetic variation among European populations of Bombus pascuorum (Hymenoptera:Apidae) from mitochondrial DNA sequence data[J]. European Journal of Entomology,2013,95(1):27-33
[16]WILLIAMS P H,BEREZIN M V,CANNINGS S G,et al. The arctic and alpine bumblebees of the subgenus Alpinobombus revised from integrative assessment of species’ gene coalescents and morphology (Hymenoptera,Apidae,Bombus)[J].Zootaxa,2019,4625(1):1-68
[17]CEJAS D,DE LA RU′A P,ORNOSA C,et al. Spatial and temporal patterns of genetic diversity in Bombus terrestris populations of the Iberian Peninsula and their conservation implications[J]. Scientific Reports,2021,11(1):22471
[18]SU M,DU L J,ALI M Y,et al. Differences in morphology,mitochondrial genomes,and reproductive compatibility between two clades of parasitic wasps Aphelinus mali (Hymenoptera:Aphelindae) in China[J]. Plos One,2023,18(2):e0279663
[19]郭冉昊,趙淑文,米福貴,等. 直立型半野生大豆葉綠體基因組分析[J]. 草地學報,2023,31(11):3334-3342
[20]郝新艷,趙淑文,劉嘉偉,等. 雜花苜蓿葉綠體基因組特征及系統發育分析[J]. 草地學報,2023,31(6):1665-1672
[21]LIN Y J,CAI L N,ZHAO Y Y,et al. Novel mitochondrial gene rearrangement and intergenic regions exist in the mitochondrial genomes from four newly established families of praying mantises (Insecta:Mantodea)[J]. Insects,2022,13(7):564-564
[22]李瑤瑤,劉云國,劉凌霄,等. 高原鰍屬魚類線粒體全基因組序列結構特征及其系統發育信息[J]. 煙臺大學學報(自然科學與工程版),2021,34(1):93-99
[23]CAMERON A S,HINES M H,WILLIAMS H P. A comprehensive phylogeny of the bumble bees (Bombus)[J]. Biological Journal of the Linnean Society,2007,91(1):161-188
[24]趙婉清,李巧,李瑩,等. 獵蝽科昆蟲線粒體基因組的比較研究[J]. 江蘇農業科學,2020,50(12):34-41
[25]彭艷,陳斌,李廷景. 黃側異腹胡蜂線粒體基因組全序列測定和分析[J]. 昆蟲學報,2017,60(4):464-474
[26]WOLSTENHOLME D R. Animal mitochondrial DNA:structure and evolution[J]. International Review of Cytology,1992(141):173-216
[27]陳建琴,張振華,邢雨輝,等. 光滑異裝蟹(Heteropanope glabra)線粒體基因組測定及其在系統發生研究中的意義[J]. 南京師大學報(自然科學版),2018,41(4):114-120
[28]韋春夢,陳亞珍,劉靜,等. 云南瘤蚱線粒體基因組特征與蚱總科系統發育分析[J]. 南方農業學報,2023,54(5):1303-1316
[29]CLARY D O,WOLSTENHOOLME D R. The mitochondrial DNA molecular of Drosophila yakuba:nucleotide sequence,gene organization,and genetic code[J]. Journal of Molecular Evolution,1985,22(3):252-271
[30]JERMIIN L S,CROZIER R H. The cytochrome b region in the mitochondrial DNA of the ant Tetraponera rufoniger:sequence divergence in Hymenoptera may be associated with nucleotide content[J]. Journal of Molecular Evolution,1994,38(3):282-294
[31]KORKMAZ E M,AYDEMIR H B,TEMEL B,et al. Mitogenome evolution in Cephini (Hymenoptera:Cephidae):Evidence for parallel adaptive evolution[J]. Biochemical Systematics and Ecology,2017(71):137-146
[32]WILLIAMS P H,CAMERON S A,HINES H M,et al. A simplified subgeneric classification of the bumblebee (genus Bombus)[J]. Apidologie,2008,39(1):46-74
(責任編輯 閔芝智)
